Open Journal of Forestry
Vol.04 No.02(2014), Article ID:43110,5 pages
10.4236/ojf.2014.42016
Restoration of Coast Redwood (Sequoia sempervirens) Forests through Natural Recovery
Will Russell, Jeff Sinclair, Kristin Hageseth Michels
Department of Environmental Studies, San José State University, San Jose, USA
Email: will.russell@sjsu.edu
Copyright © 2014 Will Russell et al. This is an open access article distributed under the Creative Commons At- tribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. In accordance of the Creative Commons Attribution License all Copyrights © 2014 are reserved for SCIRP and the owner of the intellectual property Will Russell et al. All Copyright © 2014 are guarded by law and by SCIRP as a guardian.
The management of second-growth Sequoia sempervirens (coast redwood) forests for the purpose of res- toration and ecological conservation is a growing trend. However, little is known about the long-term re- generative potential of this forest type in the absence of post-harvest management techniques such as thinning and planting. Data on forest composition and structure were collected on a chronosequence (80 - 160 years) of mature recovering stands in the southern coast redwood range using a replicated, random- ized, plot design. Results indicated that many stand characteristics including tree density, canopy cover, redwood dominance, species richness, herbaceous cover, and shrub cover reached levels statistically equivalent with old-growth reference sites in recovering stands within the time frame of this chronosequence. The recovery of individual herbaceous understory species was inconsistent however. While the cover of redwood-associated species (Oxalis oregana, Trientalis latifolia, and Disporum hookeri) reached levels statistically equivalent to old-growth reference sites, others (Trillium ovatum and Viola sempervirens) did not. Total basal area and species evenness also trended toward, but did not reach, old-growth conditions. The arboreal aspects of coast redwood forests appear to be remarkably resilient following a single logging event, and recover rapidly in the absence of active restoration techniques. The protracted recovery of certain redwood associated herbaceous understory species will require further study.
Keywords: Coast Redwood; Sequoia sempervirens; Restoration; Timber Harvest; Natural Recovery
Introduction
The majority of the original old-growth Sequoia sempervi- rens (coast redwood) forest has been converted into managed timber stands and other land uses (Noss, 2000). Preservation efforts have been successful in protecting most of the remaining old-growth in parks and preserves. However, so little of the original forest remains (<5%) that conservation groups are ac- quiring previously harvested forestlands to serve as additional preserves and buffers. While management of old-growth forests is mainly limited to preservation, management of previously harvest- ed forests often includes some level of active restoration (O’Hara et al., 2010). Whether the restoration objectives, and the tools re- quired meet those objectives, however, is a matter of debate.
In general terms, restoration is defined as “bringing back to a former position or condition” (Merriam-Webster, 2010). How- ever, defining the “former condition” of a particular forest is a difficult task. Forests, as living systems, are in a constant state of flux. Any specific point in time, or set of forest conditions, is essentially arbitrary. In North America the term “restoration” has sometimes been used to indicate an idealized pre-Euro- American condition, based on the unfounded assumption that the pre-European population did not actively manage forests (Litvalis, 2003; Anderson, 2006). Restoration efforts towards idealized conditions taken from historic photographs are equal- ly subjective, as the photographers’ choice of scenic vistas must be considered. Conditions found in currently extant reference sites with minimal anthropogenic disturbance is perhaps the best option for a restoration template (Laughlin et al., 2004; Josefsson et al., 2009), though these stands can vary signifi- cantly.
Regardless of the choice of reference, forest restoration (much like commercial timber management) has tended to fo- cus primarily on the rapid production of large trees (Meisel et al., 2009) rather than the development of viable ecological communities. As a result, silivicultural treatments in the form of pre-commercial and commercial thinning have become a common tool leading to an almost synonymous use of the terms “restoration”, “thinning”, and “fuel reduction” (Korb et al., 2007; O’Hara et al., 2010). Regrettably, the reintroduction of human disturbance to an already damaged system may be counterproductive (Brown et al., 2004). In many forest types, espe- cially coast redwood forests, natural regenerative properties allow the development of old-growth characteristics over time without the need for additional active management.
Coast redwood forests have remarkable regenerative proper- ties. They are resistant to fire, insects, and disease due to a high crown, thick insulating bark, and insect and fungi resistant sec- ondary chemicals that permeate the bark and wood (McBride, 1977; Espinosa-Garcia, & Langenheim, 1991; Veirs, 1996; Barbour et al., 2001). They are also highly resilient following disturbance due to prolific basal and epicormic sprouting (Sawyer et al., 2000; Douhovnikoff et al., 2004). Sprouting occurs when buds at the base of a tree, and under the bark throughout the bole of a tree, are released following disturbance (Roy, 1966; McBride, 1977; Sawyer et al., 2000; Barbour et al., 2001). This sprouting ability is rare among conifers, although it is present to a lesser extent in Torreya californica (California nutmeg), Pinus sylvestris (Scots pine), and Taxus brevifolia (pacific yew) (Dieguez-Aranda et al., 2005).
The ability to produce new shoots and roots from undifferen- tiated tissue is not only a disturbance survival mechanism, but also the primary reproductive strategy for S. sempervirens (Barbour et al., 2001; Douhovnikoff et al., 2004). The majority of reproduction takes place asexually resulting in a landscape populated by clonal groups. When a mature redwood is felled, due to natural or human causes, shoots are produced from the remaining stump. The shoots compete, but not in the traditional sense. Because S. sempervirens is a coppicing species, with shoots connected to a shared root system, individual shoots compete for apical dominance rather than for the survival of the organism (Kauppi et al., 1987; Burrows, 1990; Laureysens et al., 2003). As one shoot achieves dominance, other shoots begin to senesce (Sach et al., 1993) thinning the stand naturally. While S. sempervirens self-thins with facility, other coniferous forests face issues of over-crowding due to competition, which can lead to etiolation for individual trees and significant stand- level mortality due to reduced crown to height ratio and pre- disposition to insect and fungal attack (Floyd et al., 2009; Lutz & Halpern, 2006). Coast redwood forests are less susceptible to these dangers because regenerating tissue is connected to well established root systems, S. sempervirens is highly shade tole- rant, and suppressed trees can increase their crown to height ratio when released through epicormic sprouting (Sawyer et al., 2000).
The regenerative properties of S. sempervirens have allowed foresters to manage second growth stands with minimal effort for decades (Fritz, 1945). These same properties suggest that a natural regenerative approach to restoration could be successful. However, though much work has been done on the effects of forest thinning in coast redwood forests (Cole, 1983; Chittick & Keyes, 2007; O’Hara et al., 2010), very little is known about the natural processes of recovery in the absence of continued human disturbance. This study was designed to address the question; will regenerating coast redwood forests develop structure and composition comparable to old-growth forest reference sites over time in the absence of post harvest man- agement?
Methods
This analysis was conducted in the central and southern range of the coast redwood forest. Study sites were located in five State Parks in the Santa Cruz Mountains in California: Henry Cowell Redwood State Park, Butano State Park, Castle Rock State Park, Big Basin Redwood State Park, and Portola Redwood State Park. These parks support land use associated with eco-tourism such as camping, hiking, and fishing. Sur- rounding land uses, outside of the parks, include agriculture, small town residential, commercial development, and logging. The majority of the redwood forests in Santa Cruz county, in- cluding the five State Parks listed above, were heavily logged in the second half of the 19th and early part of the 20th centuries. Within the State Park system these second-growth forests have been protected for the most part, and allowed to regenerate naturally.
Vegetation in the Santa Cruz Mountains consists of forests, chaparral, coastal scrub, and grasslands (Sawyer et al., 2000). In the forests, Douglas-fir (Pseudotsuga menziesii) and S. sem- pervirens typically make up the canopy trees and the understory is typically comprised of tan oak (Notholithocarpus densiflo- rus), coast live oak (Quercus agrifolia), interior live oak (Quer- cus wislizeni), and California bay (Umbellularia californica). The soils of the area are derived from the Franciscan assem- blage, consisting mainly of sandstone and marine sediments, ideal conditions for redwood communities, which thrive on young, fertile soils with abundant nutrients and moderate pH levels (Zinke, 1988). Climate in the Santa Cruz Mountains is characterized as Mediterranean because of the predominantly mild, rainy winters and cool, dry summers (Lorimer et al., 2009). The climate of the southern range differs from the cen- tral and northern redwood region in precipitation levels which tend to be lower in the south, resulting in a longer dry season (Sawyer et al., 2000). Average annual precipitation in the Santa Cruz Mountains varies from the coast to inland. The coast av- erages 70 cm a year while the higher elevations receive up to 147 cm of precipitation per year.
Data Collection
Ten sites were selected including five old-growth reference sites and five recovering second-growth redwood sites between 80 and 160 years old, following the initial timber harvest. For inclusion in this study, each site was required to be dominated by Sequoia sempervirens, previously clear-cut, large enough for adequate sampling without edge effects (Russell & Jones, 2001), and must not have received post-harvest management such as seeding, thinning, or planting. Sites were selected using detailed timber harvest and land management history maps.
Twenty, 20 m diameter (0.031 ha), circular sample plots were randomly selected within each of the 10 study sites. Each sample plot was placed a minimum of 20 m from adjacent plots, 10 m from special habitats such as riparian areas and rock out- croppings, and 200 m from adjacent age class boundaries and main access roads. Plot size and sampling intensity were deter- mined through a pilot study using the species-area curve me- thod (Cain, 1938) and are consistent with previous research conducted in this forest type (Russell & Jones, 2001; Loya & Jules, 2007).
Data were collected on each plot to describe the composition and structure of the existing stand. Canopy cover (measured at waist height with a spherical crown densiometer), the occur- rence and abundance of each tree species, the dbh (diameter at breast height) of all individuals greater than one meter in height, and the percent cover for all understory species, were recorded for each 20 m plot. Understory perennial and herbaceous spe- cies were sampled in three 2 m diameter sub-plots within each of the 160 tree plots (Figure 1).
Data Analysis
Linear regression analyses were conducted using SPSS for analysis of correlation with stand age. Second-growth characte- ristics that were significantly correlated to age the linear regres-
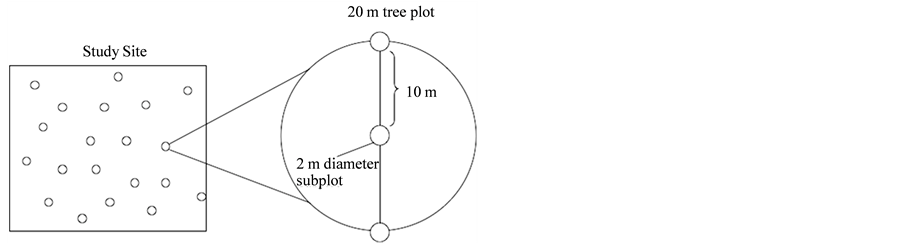
Sample plot configuration with 20 m diameter tree plot, and 2 m di- ameter nested understory sub-plots.
sion were graphed and compared to the old-growth average in order to determine the age at which the second-growth charac- teristic began trending towards or reached statistical equiva- lence with the old-growth reference average. T-tests were used to compare differences between age groups. The Levene’s test was used to check the homogeneity of variances and the K-S test was conducted to check for normality of the data.
Results
Tree Density and Dominance
Regression analysis indicated that the combined density of trees was negatively correlated with stand-age (R2 = 0.113, p = 0.038) suggesting a decline in the total stems/ha over time. In addition, the individual densities of several species were statis- tically equivalent between second-growth and old-growth ref- erence sites (Table 1), including Sequoia sempervirens (coast redwood), Aesculus californica (California buckeye), Corylus cornuta var. californica (California hazelnut), and Acer ma- crophyllum (big leaf maple). In contrast, Pseudotsuga menziesii (Douglas fir), Notholithocarpus densiflorus (tanoak), and Um- bellularia californica (California bay laurel) all had signifi- cantly higher densities in old-growth stands compared to second-growth, and Quercus wislizenii (interior oak), Arbutus menziesii (madrone) both exhibited lower densities in old- growth stands.
Sequoia sempervirens exhibited the highest relative domin- ance of all species in both second (87%) and old-growth (82%) stands, with marginally higher measures in the second-growth (p = 0.002) (Figure 2). However, average combined basal area in old-growth stands was significantly greater than in second- growth stands (p < 0.001). This pattern was repeated for indi- vidual species including Sequoia sempervirens, Pseudotsuga menziesii, and Notholithocarpus densiflorus (p = 0.000; p = 0.000; p = 0.000) suggesting that while the ratio of species within stands reached statistical equivalence with old-growth within the time frame of the chronosequence, total basal area did not.
Canopy and Understory Cover
Tree canopy cover exhibited a positive linear correlation (p = 0.002) with stand age and reached statistical equivalence with old-growth reference sites at 140 years (Figure 3). A high de- gree of variation between samples (52% - 91% in second- growth and 50% - 89% in old-growth) resulted in fairly low
predictive power for the regression (R2 = 0.113). However, this variation was consistent between old-growth and second- growth sites and appeared to be an indicator of natural stand complexity rather than an artifact of human management. The average cover of perennial shrubs was statistically equivalent between second-growth (14% ± 1.346% std. error) and old- growth (16% ± 1.98% std. error) stands (p = 0.893). The aver- age cover of herbaceous species was also found to be statisti- cally equivalent between second-growth (5% ± 1.06% std. error) and old-growth (7.88 ± 1.45 std. error) (p = 0.963) (Figure 4). Individually, the recovery of specific understory species was mixed. The recovery of Trillium ovatum (western wake robin), a perennial associated with old-growth forests, was incomplete with significantly lower cover found in second-growth com- pared to old-growth stands (Table 2). This was also true for Viola sempervirens (redwood violet), a shade tolerant herba- ceous species closely associated with coast redwood forests. In contrast, three other species closely associated with coast red- wood forests―Oxalis oregana (redwood sorrel), Trientalis latifolia (pacific starflower), and Disporum hookeri (Hooker’s fairy bells)―reached levels of cover statistically equivalent to old-growth reference sites within recovering stands.
Total species richness was positively correlated with stand age in second-growth (p = 0.000, R2 = 0.43), reaching old- growth equivalence at 100 years. However, there was greater variation in richness in second-growth stands (15.36 species/ plot, 0.64 std. error) than in old-growth stands (10.33 species/ plot, 0.46 std. error). Total understory species richness was also significantly correlated with stand age (p = 0.000) (R2 = 0.44). Combined second-growth understory richness (11.78 species/ plot, 0.57 std. error) was significantly higher than old-growth understory richness (7.36 species/plot, 0.45 std. error) (p = 0.000).
Shannon-Weaver diversity (evenness) was positively corre- lated with stand age (p = 0.000, R2 = 0.50), indicating a trend toward old-growth conditions. The index did not reach old- growth equivalence however, and combined evenness was sig- nificantly higher in second-growth stands (1.63, 0.61 std. error) than old-growth stands (0.97, 0.57 = std. error).

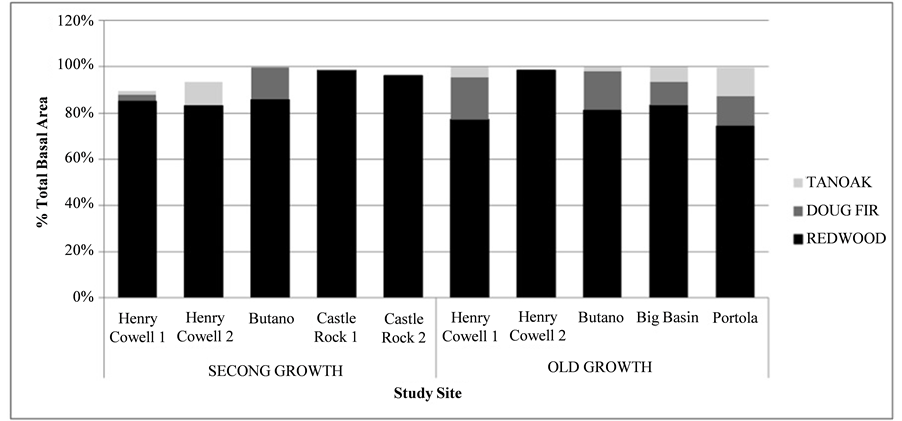
Relative dominance of the three most common tree species―Sequoia sempervirens (Coast Redwood), Pseudotsuga menziesii (Douglas-fir), and Notholithocarpus densiflorus (tanoak)―on ten sites in the Santa Cruz mountains.
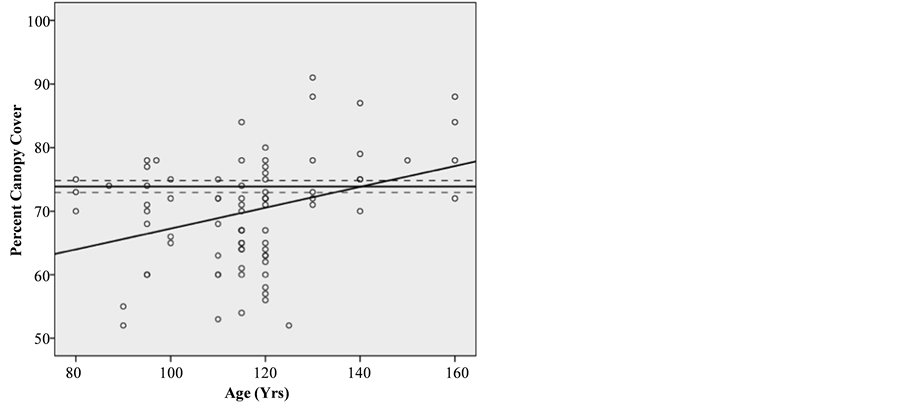
Linear Regression of percent canopy cover on a chronosequence of second-growth stands with old-growth percent canopy cover mean re- ference line and standard error confidence band.
Conclusion
The results of this study indicate that natural recovery is an effective technique for the restoration of coast redwood forests. The overall density of trees declined over time in recovering stands reaching statistical equivalence with old-growth refer- ence sites for most tree species. The dominance of S. sempervi- rens also reached statistical equivalence with old-growth, as did canopy cover, understory cover, and species richness. Asso- ciated herbaceous species also trended toward recovery, though the cover of some species (Trillium ovatum and Viola semper- virens) did not reach statistical equivalence with old-growth reference sites in the time frame of this chronosequence. Active management tools such as stand thinning are counter indicated as they are generally employed as a restoration technique to
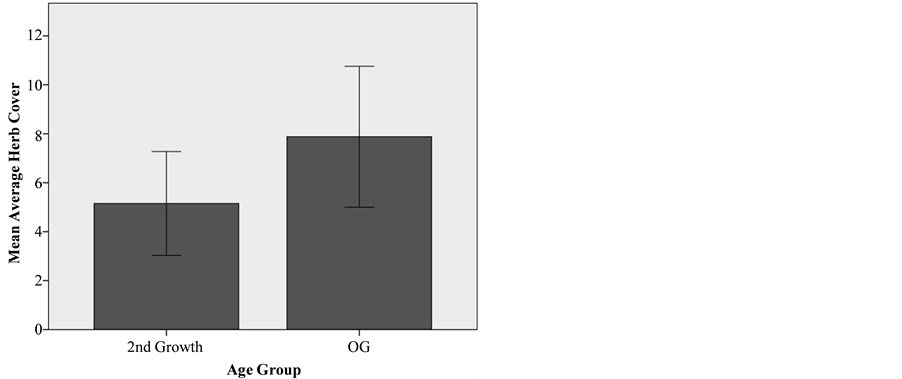
Mean averaged herb cover in coast redwood second-growth and old- growth stands with 95% confidence intervals.
reduce tree density in overstocked stands and to promote the dominance of desired species. Additionally, the added distur- bance of mechanical thinning is likely to impact sensitive un- derstory herbaceous species that are dependent on the moist shady environment provided by a mature forest canopy.
Discussion
Forest structure and composition on the chronsequence ana- lyzed exhibited significant trends toward old-growth conditions for stand density, the relative dominance of Sequoia sempervi- rens, tree canopy cover, shrub cover, species richness, and total herbaceous cover. However, results were mixed for the shade- adapted understory species that are common in healthy coast redwood stands, with some recovering fully while others did not. This finding was concerning, though not unexpected. Pre- vious research on a chronosequence in the central part of the
coast redwood range offered similar results (Russell & Michels, 2010). Overall, the arboreal features of the forest appeared to be quite resilient to human disturbance, while shade loving herba- ceous species were not. With this in mind, implementation of mechanical restoration techniques, such as stand thinning, are counter indicated.
Mechanical disturbance in the form of stand thinning has been presented as an effective restoration technique for many forest types as a means of quickly returning stands to old- growth density levels (Boe, 1965; Bosch, 1971; Oliver et al., 1994; Brown et al., 2004; Lindquist, 2004; Smith et al., 2005; North et al., 2007). Such treatments can also increase the growth rate of selected trees through the removal of competing individuals thereby promoting the growth of large individual trees (Cole, 1983; Oliver et al., 1994; Lindquist, 2004; O’Hara et al., 2007).
In coast redwood forests, however, this model is not well ap- plied, as regenerative clonal sprouts do not compete in the tra- ditional sense. In addition, mechanical thinning can have unin- tended consequences. Removal of trees exposes the forest floor to increased levels of solar radiation (Kjeldsen-Ederer & Rivas, 1998; Russell & Jones, 2001) allowing opportunistic non-native species to colonize disturbed areas, and can alter soil conditions through compaction and reduction of nitrogen levels (Corns, 1988; Ebrect & Schmidt, 2003; Jussy et al., 2004). Thinning has been shown to decrease epicormic sprouting response of coast redwood (Powers & Wiant, 1970; Cole, 1983) and allows for non-native shade intolerant species to persist in the commu- nity. In addition, stand density often increases in coast redwood stands a few years following thinning as a response to increased solar radiation (Russell & Jones, 2001), thus the effects of me- chanical thinning are temporary.
The purpose of stand thinning as a restoration tool is to speed forest development in order to produce certain old-growth cha- racteristics more quickly. However, the development of a forest with a dominant species, such as Sequoia sempervirens, that commonly persists for 1500 to 2000 years is a lengthy process. And though the temptation to manipulate a regenerating forest so that management goals can be reached in a human time scale is compelling, the ecological costs of continued disturbance, particularly when using the same tools that caused the original damage, must be considered. The reason why some associated understory species reached old-growth equivalence, and others did not, is not clear. However, the answer likely relates to soil conditions and the relationships between plant roots and com- munities of soil microorganisms. Therefore, the protection of the soil from further human disturbance is essential so that nat- ural soil development processes can occur. In addition, allow- ing natural processes to restore a system allows for stochastic factors to create stand complexity, while eliminating the sim- plifying influence of subjective human management.
Busing and Fujimori (2002; 2005) determined that small- scale natural disturbances and unmanaged processes of com- munity development were sufficient for the regeneration of coast redwood forests. The results of this study support Busing and Fujimori’s conclusions by providing quantitative data on the development of coast redwood stands over a significant period of time. For maximum effectiveness, the focus of resto- ration should shift from arboreal canopy species to the recovery of old-growth associated understory species, such as Trillium ovatum, that have been shown to be severely impacted through logging (Jules & Rathcke, 1999).
Acknowledgements
This research was supported by grants from the Save-the- Redwoods League and the Mendocino Institute. Logistical support was provided by the California Department of Parks and Recreation.
References
- Anderson, K. (2006). Tending the wild: Native American knowledge and the management of California’s natural resources. Berkeley, CA: University of California Press.
- Barbour, M., Lydon, S., Borchert, M., Popper, M., Whitworth, V., & Evarts, J. (2001). Coast redwood: A natural and cultural history. Los Olivos, CA: Cachuma Press.
- Boe, K. (1965). Natural regeneration in old-growth redwood cuttings. Berkeley, CA: USDA Forest Service, Pacific Southwest Forest and Range Experiment Station, Research Note PSW 94.
- Bosch, C. A. (1971). Redwoods: A population model. Science, 172, 345- 349. http://dx.doi.org/10.1126/science.172.3981.345
- Brown, R. T., Agee, J. K., & Franklin, J. F. (2004). Forest restoration and fire: Principles in the context of place. Conservation Biology, 18, 903-912. http://dx.doi.org/10.1111/j.1523-1739.2004.521_1.x
- Burrows, G. E. (1990). The role of axillary meristems in coppice and epicormic bud initiation in Araucaria cunninghamii. Botanical Ga- zette, 151, 293-301. http://dx.doi.org/10.1086/337829
- Busing, R. T., & Fujimori, T. (2002). Dynamics of composition and structure in an old Sequoia sempervirens forest. Vegetation Science, 13, 785-792.
- Busing, R. T., & Fujimori, T. (2005). Biomass, production and woody detritus in an old coast redwood (Sequoia sempervirens) forest. Plant Ecology, 177, 177-188. http://dx.doi.org/10.1007/s11258-005-2322-8
- Cain, S. (1938). The species-area curve. American Midland Naturalist, 19, 573-581. http://dx.doi.org/10.2307/2420468
- Chittick, A. J., & Keyes, C. R. (2007). Holter Ridge thinning study, Redwood National Park: Preliminary results of a 25-year retrospec- tive. Albany, California: Pacific Southwest Research Station, USDA Forest Service
- Cole, D. W. (1983). Redwood sprout growth three decades after thin- ning. Journal of Forestry, 81, 148-157.
- Corns, I. W. (1988). Compaction by forestry equipment and effects on coniferous seedling growth on four soils in the Alberta foothills. Ca- nadian Journal of Forest Research, 18, 75-84. http://dx.doi.org/10.1139/x88-012
- Dieguez-Aranda, U., Castedo-Dorado, F., Alvarez-Gonzalez, J. G., Ro- drıguez-Douhovnikoff, V., Cheng, A. M., & Dodd, R. S. (2004). In- cidence, size, and spatial structure of clones in second-growth stands of Coast Redwood, Sequoia sempervirens (Cupressaceae). American Journal of Botany, 9, 1140-1146.
- Ebrecht, L., & Schmidt, W. (2003). Nitrogen mineralization and vege- tation along skidding tracks. Annals of Forest Science, 60, 733-740. http://dx.doi.org/10.1051/forest:2003067
- Espinosa-Garcia, F. J., & Langenheim, J. H. (1991). Effect of some leaf essential oil phenotypes in coastal redwood on the growth of several fungi with endophytic stages. Biochemical Systematics and Ecology, 19, 629-642. http://dx.doi.org/10.1016/0305-1978(91)90079-F
- Floyd, M. L., Clifford, M., Cobb, N. S., Hanna, D., Delph, R., Ford, P., & Turner, D. (2009). Relationship of stand characteristics to drought- induced mortality in three Southwestern pinon-juniper woodlands. Ecological Applications, 19, 1223-1230. http://dx.doi.org/10.1890/08-1265.1
- Fritz, E. (1945). Twenty years’ growth on a redwood sample plot. Journal of Forestry, 43, 30-36.
- Josefsson, T., Hornberg, G., & Ostlund, L. (2009). Long-term human impact and vegetation changes in a boreal forest reserve: Implica- tions for the use of protected areas as ecological references. Ecosys- tems, 12, 1017-1036. http://dx.doi.org/10.1007/s10021-009-9276-y
- Jules, E. S., & Rathcke, B. J. (1999). Mechanisms of reduced trillium recruitment along edges of old-growth forest fragments. Conserva- tion Biology, 13, 784-793. http://dx.doi.org/10.1046/j.1523-1739.1999.97435.x
- Jussy, J. H., Ranger, J., Bienaime, S., & Dambrine, E. (2004). Effects of a clear-cut on the in situ nitrogen mineralisation and the nitrogen cycle in a 67-year-old Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) plantation. Annals of Forest Science, 61, 397-405. http://dx.doi.org/10.1051/forest:2004033
- Kauppi, A., Rinne, P., & Ferm, A. (1987). Initiation, structure and sprout- ing of dormant basal buds in Betula pubescens. Flora, 179, 55-83.
- Kjeldsen-Ederer, C. K., & Rivas-Ederer, D. (1998). Seral vascular plant communities on clearcut sites in Jackson Demonstration State Forest, Mendocino County, California. Rohnert Park, CA: Sonoma State University, California Department of Forestry and Fire Protec- tion.
- Korb, J. E., Fulé, P. Z., & Gideon, B. (2007). Different restoration thinning treatments affect level of soil disturbance in ponderosa pine forests of Northern Arizona, USA. Ecological Restoration, 25, 43- 49. http://dx.doi.org/10.3368/er.25.1.43
- Laughlin, D. C., Bakker, J. D., Stoddard, M. T., Daniels, M. L., Sprin- ger, J. D., Gildar, C. N., Green, A. M., & Covington, W. W. (2004). Toward reference conditions: Wildfire effects on flora in an old- growth ponderosa pine forest. Forest Ecology and Management, 199, 137-152. http://dx.doi.org/10.1016/j.foreco.2004.05.034
- Laureysens, I., Deraedt, W., Indeherberge, T., & Ceulemans, R. (2003). Population dynamics in a 6-year old coppice culture of poplar: Clon- al differences in stool mortality, shoot dynamics and shoot diameter distribution in relation to biomass production. Biomass and Bioener- gy, 24, 81-95. http://dx.doi.org/10.1016/S0961-9534(02)00105-8
- Lindquist, J. L. (2004b). Growth and yield report for the whiskey springs redwood commercial thinning study: A twenty-nine year sta- tus report (1970-1999) (25 p). California Forestry Report (3), Sacramento, California: California Department of Forestry and Fire Protection.
- Litvaltis, J. A. (2003). Are pre-Columbian conditions relevant baselines for managed forests in the northeastern United States? Forest Ecolo- gy and Management, 185, 113-126. http://dx.doi.org/10.1016/S0378-1127(03)00250-0
- Lorimer, C. G., Porter, D. J., Madej, M. A., Stuart, J. D., Veirs, S. D., Norman, S. P., O’Hara, K. L., & Libby, W. J. (2009). Presettlement and modern disturbance regimes in coast redwood forests: Implica- tions for the conservation of old-growth stands. Forest Ecology and Management, 258, 1038-1054. http://dx.doi.org/10.1016/j.foreco.2009.07.008
- Loya, D. T., & Jules, E. S. (2007). Use of species richness estimators improves evaluation of understory plant response to logging: A study of redwood forests. Plant Ecology, 194, 179-194. http://dx.doi.org/10.1007/s11258-007-9283-z
- Lutz, J. A., & Halpern, C. B. (2006). Tree mortality during early forest development: A long-term study of rates, causes, and consequences. Ecological Monographs, 76, 257-275. http://dx.doi.org/10.1890/0012-9615(2006)076[0257:TMDEFD]2.0.CO;2
- McBride, J. R. (1977). The ecology of redwood [Sequoia sempervirens (D. Don) Endl.] and the impact of man’s use of the redwood forest as a site for recreational activities. Arcata, Csalifornia: California De- partment of Forestry and Fire Protection. Report Prepared for the United States National Park Service.
- Merriam-Webster Online Dictionary (2010). Restoration. http://www.merriam-webster.com
- Miesel, J. R., Boerner, R. E. J., & Skinner, C. N. (2009). Mechanical restoration of California mixed-conifer forests: Does it matter which trees are cut? Restoration Ecology, 17, 784-795. http://dx.doi.org/10.1111/j.1526-100X.2008.00414.x
- North, M., Innes, J., & Zald, H. (2007). Comparison of thinning and prescribed fire restoration treatments to Sierran mixed-conifer his- toric conditions. Canadian Journal of Forest Research, 37, 331-342. http://dx.doi.org/10.1139/X06-236
- Noss, R. F. (2000). The redwood forest: History, ecology, and conser- vation of the coast redwoods. In: R. F. Noss (Ed.), More than big trees (pp. 1-6). Washington DC: Island Press.
- O’Hara, K. L., Nesmith, J. C. B., Leonard, L., & Porter, D. L. (2010). Restoration of old forest features in coast redwood forests using early- stage variable-density thinning. Restoration Ecology, 18, 125-135. http://dx.doi.org/10.1111/j.1526-100X.2010.00655.x
- O’Hara, K. L., Stancioiu, P. T., & Spencer, M. A. (2007). Understory stump sprout development under variable canopy density and leaf area in coast redwood. Forest Ecology and Management, 244, 76-85. http://dx.doi.org/10.1016/j.foreco.2007.03.062
- Oliver, W. W., Lindquist, J. L., & Strothmann, R. O. (1994). Young- growth redwood stands respond well to various thinning intensities. Western Journal of Applied Forestry, 9, 106-112.
- Powers, R. F., & Wiant, H. V. (1970). Sprouting of old-growth coastal redwood stumps on slopes. Forest Science, 16, 339-341.
- Roy, D. F. (1966). Silvicultural characteristics of redwood (Sequoia sempervirens [D.Don] Endl.). Albany, California: Pacific Southwest Research Station, USDA Forest Service.
- Russell, W. H., & Jones, C. (2001). The effects of timber harvesting on the structure and composition of adjacent old-growth coast redwood forest, California, USA. Landscape Ecology, 16, 731-741. http://dx.doi.org/10.1023/A:1014486030462
- Russell, W., & Michels, K. H. (2010). Stand development on a 127- year chronosequence of naturally regenerating Sequoia sempervirens (Taxodiaceae) forests. Madrono, 57, 229-241. http://dx.doi.org/10.3120/0024-9637-57.4.229
- Sachs, T., Novoplansky, A., & Cohen, D. (1993). Plants as competing populations of redundant organs. Plant, Cell and Environment, 16, 765-770. http://dx.doi.org/10.1111/j.1365-3040.1993.tb00498.x
- Sawyer, J. O., Sillett, S. C., Popenoe, J. H., LaBanca, A., Sholars, T., Largent, D. L., Euphrat, F., Noss, R. F., & Van Pelt, R. (2000). Cha- racteristics of redwood forests. In R. F. Noss (Ed.), The redwood forest: History, ecology, and conservation of the coast redwoods (pp. 39-79). Washington DC: Island Press.
- Smith, J. E., McKay, D., Brenner, G., McIver, J., & Spatafora, J. W. (2005). Early impacts of forest restoration treatments on the ecto- mycorrhizal fungal community and fine root biomass in a mixed conifer forest. Journal of Applied Ecology, 42, 526-535. http://dx.doi.org/10.1111/j.1365-2664.2005.01047.x
- Veirs, S. D. (1996). Ecology of the coast redwood. Proceedings of Conference on Coast Redwood Forest Ecology and Management. Arcata, CA: Humboldt State University.
- Zinke, P. J. (1988). The redwood forest and associated north coast forests. In M. G. Barbour, & J. Major (Eds.), Terrestrial vegetation of California (2nd ed., pp. 670-699). Sacramento, CA: California Plant Society Press.