Advances in Biological Chemistry
Vol.4 No.1(2014), Article ID:42708,10 pages DOI:10.4236/abc.2014.41003
Roles of NADPH oxidase 2 and 4 in endothelial cell survival and death under serum depletion
![]()
Department of Molecular Medicine, Cell and Matrix Research Institute, BK21 plus Program, Kyungpook National University School of Medicine, Daegu, Republic of Korea
Email: ycboo@knu.ac.kr
Copyright © 2014 Hyeona Jeon, Yong Chool Boo. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. In accordance of the Creative Commons Attribution License all Copyrights © 2014 are reserved for SCIRP and the owner of the intellectual property Hyeona Jeon, Yong Chool Boo. All Copyright © 2014 are guarded by law and by SCIRP as a guardian.
Received 10 December 2013; revised 20 January 2014; accepted 29 January 2014
KEYWORDS
Endothelial Cells; Apoptosis; Serum Depletion; Reactive Oxygen Species; NADPH Oxidases
ABSTRACT
Oxidative stress and redox-signal pathways are known to be involved in endothelial apoptosis induced by serum depletion. However, the associated mechanism is not well understood and thus, was investigated in the present study focusing on NADPH oxidases (NOX). Serum removal from the culture medium led to an increase in reactive oxygen species (ROS) production and apoptotic death of human umbilical vein endothelial cells. Serum depletion also increased the gene expression of the NOX2 and NOX4 subunits. The selective suppression of NOX4 expression by small interfering RNA (siRNA) attenuated ROS production and cell death due to serum-depletion whereas siRNA for NOX2 increased cell death. Expression of exogenous NOX2 or NOX4 subunit alone had no significant effects on ROS production or cell death. Coexpression of the subunits of the NOX4 complex (NOX4 and p22phox) or the NOX2 complex (NOX2, p22phox, p47phox and p67phox) increased ROS production and cell death under serum-depleted conditions. This study suggests that endothelial cell survival and death are differentially regulated by expression levels of the subunits of NOX2 and NOX4 complexes.
1. INTRODUCTION
The endothelium constituting the inner surface of blood vessels plays a key role in the regulation of vascular physiology and structure through the production of a variety of substances including nitric oxide [1,2]. An imbalance between endothelial cell apoptosis and renewal causes discontinuity of the endothelial layer, favoring the initiation and progression of pathological events that lead to atherosclerosis, plaque rupture, and thrombosis [3]. In patients with cardiovascular risk factors, increased endothelial apoptosis results in endothelial dysfunction which is a common mechanism affecting vascular function and structure [4,5]. This is supported by mounting evidence showing the association of endothelial dysfunction with markers of vascular damage and with cardiovascular events in patients with essential hypertension and/or atherosclerotic disease [6].
Endothelial cells are physiologically exposed to the blood stream which functions in the transport of nutrients/ wastes, growth factors/cytokines and oxygen/carbon dioxide. Various cardiovascular risk factors such as increased oxidized low-density lipoprotein can increase endothelial apoptosis [7,8]. Fluid shear stress due to blood flow can have positive or negative effects on endothelial cell survival [9]. In addition, under such clinical conditions as ischemia, angioplasty, and organ transplantation, endothelial cells may experience shortages of oxygen, nutrients and growth factors which can lead to increased apoptosis [10]. Reactive oxygen species (ROS) from various biological sources mediate oxidative stress and redox signaling involved in endothelial apoptosis [11, 12].
The NAD(P)H oxidase (NOX) family is a well established source of ROS that confers oxidative stress and mediates redox signaling pathways in cells [13,14]. NOX family members include NOX1, NOX3, NOX4, NOX5, DUOX1 and DUOX2. Although all these members share a common activity to transfer electrons from NADPH to oxygen, they are different in terms of subunit composition, enzyme activation mechanism, catalytic mechanism producing specific ROS, subcellular localization, and so on. The NOX2 complex consists of the membrane proteins NOX2 (CYBB, gp91phox) and p22phox (CYBA), and cytosolic proteins such as p47 phox, and p67phox, whereas the NOX4 complex has the membrane proteins NOX4 and p22phox (CYBA) [13]. NOX4 is known to generate hydrogen peroxide in preference of superoxide anion radical, unlike other NOX isoforms [15,16]. NOX-derived ROS has been demonstrated to stimulate cell death by stimulating apoptotic signaling pathways directly and indirectly [12,17,18]. In some circumstance, however, NOX-derived ROS can also exhibit prosurvival effects [19-21]. Thus, it seems likely that NOX-derived ROS can induce either cell death or cell survival depending on where and how much they are produced. In addition, different isotypes of NOX family can influence the cell fate differently. In a previous study, we have demonstrated that NOX2 mediated the prosurvival effect of laminar shear stress on endothelial cells whereas NOX4 had the opposite effects [22].
The absence of serum with plenty of growth factors is a strong inducer of endothelial cell death. Although oxidative stress is presumed to be involved in endothelial apoptosis induced by serum depletion, the associated molecular mechanism is not well understood. In the present study, we examined the role of NOX2 versus NOX4 in endothelial apoptotic cell death due to serum depletion. These isoforms were chosen because they are known to be constitutively expressed in endothelial cells [19,23].
2. MATERIALS AND METHODS
2.1. Cell Culture and Treatments
Human umbilical vein endothelial cells (HUVECs) obtained from Clonetics Cambrex (Rockland, ME, USA) were cultured on 0.2% gelatin coated culture dishes at 37˚C and 5% CO2. The growth medium was EBM-2 medium supplemented with 10% fetal bovine serum (Gibco BRL, Grand Island, NY, USA), endothelial growth supplements (Clonetics Cambrex), and antibiotics (100 U·mL−1 penicillin, 100 μg·mL−1 streptomycin and 0.25 μg·mL−1 amphotericin B). Cells were cultured in the growth medium to a cell confluence of ~95%. The medium was replaced with a fresh growth medium with or without serum plus endothelial supplements.
2.2. Transfection with a Small Interfering RNA (siRNA)
Human NOX2 siRNA (#1299001, HSS102523), Human NOX4 siRNA (#1299001, HSS121314), and a negative control siRNA with scrambled sequences (#12935200) were purchased from Invitrogen (Grand Island, CA, USA). The nucleotide sequences of the NOX2 and NOX4 siRNAs were as follows: NOX2 siRNA, 5’-GGG UUU AUG AUA UUC CAC CUA AGU U-3’ (sense) and 5’-AAC UUA GGU GGA AUA UCA UAA ACC C-3’ (antisense); NOX4 siRNA, 5’-CCU CAU GAU CAC AGC CUC UAC AUA U-3’ (sense) and 5’-AUA UGU AGA GGC UGU GAU CAU GAG G-3’ (antisense). For transfection, cells were treated with a mixture of 100 nM siRNA and 1.25 μL mL−1 Lipofectamine RNAiMAX (Invitrogen) in Opti-MEM (Invitrogen) for 4 h and then the cells were fed with the growth medium. After 2 days, the cells were exposed to serum depletion for the indicated time.
2.3. Transfection with a Plasmid DNA
Total RNA from HUVECs was used for the synthesis of first strand cDNA using the Reverse Transcription Master Premix (ELPIS-Biotech, Daejeon, Korea). The full coding sequence of human p67phox was PCR-amplified from the first-strand cDNA using primers 5’-CCG AAG CTT GCC ACC ATG TCC CTG GTG GAG GCC AT-3’ and 5’-GCG TCT AGA CTA GAC TTC TCT CCG AGT GCT TTC C-3’ (HindIII and XbaI sites were incorporated into the sense and antisense primers, respectively, as underlined).The PCR products were gel-purified using the QIA Quick gel extraction kit (Qiagen, Valencia, CA, USA), digested with HindIII and XbaI (New England Biolabs, Ipswich, MA, USA) and ligated with the pcDNA3.1(+) vector (Invitrogen) using the Ligation Mix Kit (TaKaRaBio Inc., Shiga, Japan). The plasmid was cloned. Plasmid clones of human NOX2 in pEZ-M02 vector (#EX-T8138-M02), human NOX4 (#EX-W0105- M02), human p22phox (#EX-C0158-M02), human p47phox (#EX-Z5866-M02) and EGFP (enhanced green fluorescent protein) in pReceiver-M02 vector (#EX-EGFPM02) were purchased from GeneCopoeia (Rockville, MD, USA). The plasmid clones were propagated in the Escherichia coli strain DH5α (TaKaRa Bio Inc., Shiga, Japan) and purified using the Maxi-Prep DNA purification kit (Qiagen, Valencia, CA, USA).The coding sequences of the plasmid clones were verified to be identical to their reference sequences: NOX2 (CYBB, gp91phox) (NM_000397.3), NOX4 (NM_016931.3, NM_001143836.1, NM_001143837.1), CYBA (p22phox) (NM_000101.3), NCF1 (p47phox) (NM_000265.4) and NCF2 (p67phox) (NM_000433.3, NM_001127651.2, NM_001190794.1, NM_001190789.1). Transfection of HUVECs with the plasmid DNA was performed with TrueFectTM (United BioSystems Inc., Rockville, MD, USA). Briefly, cultured cells at ~90% confluence were treated with 1 μg·mL−1 plasmid DNA and 3 μL·mL−1 TrueFectTM in Opti-MEM for 4 h, followed by incubation in a growth medium.
2.4. Cell Viability Assay
Cell viability was determined by the trypan blue exclusion assay. Briefly, the cells that lost attachment were collected by centrifuging (2500 rpm, 5 min) the culture medium, and the attached cells were harvested by trypsinization followed by centrifugation. The cells were collectively suspended in medium. 0.4% trypan blue solution (Sigma, St Louis, MO, USA) was added to the cell suspension at a one to one ratio. The numbers of stained dead cells and unstained live cells were counted on a hemocytometer under a microscope. Cell death (%) was defined as follows: cell death (%) = [(Ad)/(Ad + Av)] × 100%, where Ad is the number of the dead cells and Av is the number of viable cells.
2.5. Detection of ROS
The production of ROS was determined with the oxidant-sensitive probe dehydrorhodamine (DHR) 123. HUVECs were treated with 1.0 μM DHR 123 for 2 h. Images of cells fluorescing due to the oxidation of DHR 123 to rhodamine 123 were obtained with a Nikon Eclipse TE2000-U microscope. The formed rhodamine 123 was extracted from cells using ice-cold 70% ethanol that contained 0.1 N HCl, followed by centrifugation at 13,000 rpm for 15 min. The supernatants were neutralized with 1 M NaHCO3 and centrifuged again to remove precipitates. The fluorescence intensity of clear supernatant aliquots was measured at an emission wavelength of 590 nm (excitation at 485 nm) using the Gemini EM fluorescence microplate reader (Molecular devices, Sunnyvale, CA, USA).
2.6. Quantitative Real Time-Polymerase Chain Reaction (qRT-PCR) Analysis
Total cellular RNA was extracted with the RNeasy kit (Qiagen, Valencia, CA, USA). To prepare the cDNA, 1 μg of cellular mRNA was reverse-transcribed with the High Capacity cDNA Archive Kit (Applied Biosystems, Foster city, CA, USA). The kit utilizes random hexamers primers and MultiScribeTM Reverse Transcriptase. PCR was done with the StepOnePlusTM Real-Time PCR System (Applied Biosystems) in a reaction mixture (20 μL) containing SYBR®Green PCR Master Mix (Applied Biosystems), 500 ng cDNA, and 10 picomoles of genespecific primer sets (Macrogen, Seoul, Korea). The reactions were performed with the following conditions: 50˚C for 2 min, 95˚C for 10 min, 40 cycles of 95˚C for 15 s and 60˚C for 1 min, followed by a dissociation protocol. Single peaks in the melting curve analysis supported the homogeneity of the amplicons. The mRNA expression level relative to the internal control glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was calculated by the comparative threshold cycle method. The sequences of the PCR primers were as follows: NOX2 (CYBB, gp91phox) (GeneBank accession number, NM_ 000397.3) 5’-TGTTAG TGG GAG CAG GGA TTG-3’ (forward) and 5’-TCA GAT TGGTGG CGT TAT TGC- 3’ (reverse); NOX4 (NM_016931.3, NM_001143836.1, NM_001143837.1) 5’-GCA TGT GGC TGC CCA TCT- 3’ (forward) and 5’-GCC AGG AAC AGT TGT GAA GAG A-3’ (reverse); CYBA (p22phox) (NM_000101.3) 5’-ACT TTG GTG CCTACT CCA TTG TG-3’ (forward) and 5’-GGC ACC GAG AGC AGGAGA T-3’ (antisense); NCF1 (p47phox) (NM_000265.4) 5’-CAG CCA GCA CTA TGT GTA CA-3’ (forward) and 5’-GAA CTC GTA GAT CTC GGT GAA-3’ (reverse); NCF2 (p67phox) (NM_000433.3, NM_001127651.2, NM_ 001190794.1, NM_001190789.1) 5’-CCT CCA CCC AGA CCG AAA A-3’ (forward) and 5’-CCG TGG CCC AGT TAT CAT TG-3’ (reverse); GAPDH (NM_ 002046.3) 5’-ATG GGG AAG GTG AAG GTC G-3’ (forward) and 5’-GGG GTC ATT GAT GGC AAC AA-3’ (reverse).
2.7. Western Blotting
Cells were lysed in a lysis buffer (20 mM Tris-Cl, 2.5 mM EDTA, 1.0% SDS, pH 7.5) supplemented with 1 mM phenyl methyl sulfonyl fluoride and protease inhibitor cocktail (Roche, Mannheim, Germany). Proteins were denatured in Laemmli sample buffer for 5 min at 95˚C, resolved on 12% SDS-PAGE gel and transferred to a polyvinylidene difluoride membrane (Amersham Pharmacia, Little Chalfont, UK). The membranes were incubated overnight with a primary antibody at 4˚C, and then with a secondary antibody conjugated to horseradish peroxidase (Cell Signaling, Danvers, MA, USA) for 1 h at room temperature. The immunoreactive bands were detected with a picoEPD Western Reagent kit (ELPISBiotech, Daejeon, Korea) an ECL kit (Amersham Pharmacia, Little Chalfont, UK) and analyzed with the NIH Image program. Rabbit polyclonal caspase-3 antibody and rabbit polyclonal caspase-9 were purchased from Cell Signaling. Mouse monoclonal β-actin antibody was from Sigma-Aldrich.
2.8. Statistical Analysis
Data are presented as the means ± SE. The statistical analyses were done with the Sigma Stat 3.1 software program. Significant differences among the groups were determined with one way ANOVA. Duncan’s multiplerange test was done if differences were identified between the groups with a significance level of p < 0.05.
3. RESULTS
We examined whether serum removal from the cell culture medium induces ROS production and apoptotic cell death of HUVECs. We used DHR123 as a fluorogenic probe for ROS detection. This probe is oxidized to highly fluorescent products by highly reactive ROS such as peroxynitrite. Neither NO, superoxide, nor hydrogen peroxide alone is not able to oxidize DHR123 directly. Thus this probe is useful in detection of ROS more associated with oxidative stress. When cultured HUVECs were exposed to serum depleted conditions, ROS production increased significantly as monitored by conversion of DHR123 to fluorescent compounds (Figures 1(a) and(b)). Serum depletion also caused cell death (Figure 1(c)). The serum depletion-induced cell death was confirmed to involve apoptotic mechanisms such as the activation of caspase-3 and caspase-9 (Figure 1(d)). Effect of serum depletion on the gene expressions of NOX2, NOX4, p22phox, p47phox and p67phox were determined by qRT-PCR analysis. Serum depletion increased the expression levels of NOX2, NOX4, and p47phox (Figure 1(e)), suggesting potential roles of NOX2 and NOX4 complexes in serum depletion-induced ROS production and apoptotic cell death.
In order to examine the roles of the NOX systems, we used the siRNA approach to selectively decrease the expression of NOX2 and NOX4 catalytic subunits. HUVECs pretreated with the siRNA of NOX2 or NOX4 or scrambled negative control siRNA were exposed to serum-provided and serum-depleted conditions for 2 h for ROS measurement (Figure 2(a)) or 24 h for the cell viability assay (Figure 2(b)). Expression of NOX2 and NOX4 at the mRNA levels were examined by qRT-PCR analysis (Figure 2(c)). Serum depletion-induced ROS production was significantly attenuated by NOX4 siRNA but not by NOX2 siRNA (Figure 2(a)). NOX4siRNA also significantly attenuated cell death due to serum depletion, whereas siRNA of NOX2 increased cell death (Figure 2(b)). Specific down regulation of NOX2 and NOX4 expression was verified by quantitative mRNA analysis (Figure 2(c)). In accordance with the cell death data, the level of cleaved caspase-3 and cleaved caspase-9 increased by siNOX2 treatment but decreased by siNOX4 treatment (Figure 2(d)).
We next examined the effect of overexpression of the NOX2 and NOX4 catalytic subunits on ROS production, cell viability and caspase activation. Cells were transfected with an equal amount of plasmid DNA encoding NOX2 or NOX4 and cultured for 48 h, followed by 24 h of serum depletion or not. Differently to our initial expectation, the ROS production, cell death and caspase activation of HUVECs under serum-provided and serumdepleted conditions were not significantly increased by overexpression of the NOX2 subunit alone or NOX4 subunit alone (Figure 3). These result led us to assume that overexpression of NOX2 or NOX4 subunit alone may not enough to increase ROS production causing cell death, and that ROS production may require expression or activation of other protein partners.
This notion was tested in the following experiment by transfecting cells with plasmids encoding NOX2, p22phox, p47phox and p67phox together, or transfecting with plasmids encoding NOX4 and p22phox together to rebuild the whole complexes of NOX2 or NOX4 in cells. Total amount of plasmids were kept equal in all cells by adding the appropriate amount of vector. Coexpression of the subunits of NOX complex (NOX2, p22phox, p47phox and p67phox) or NOX4 complex (NOX4 and p22phox) increased the ROS production (Figure 4(a)) and cell death rate (Figure 4(b)) under serum-depleted conditions, compared to the cells transfected with the EGFP vector. Western blot analysis also showed that caspase-3 and caspase-9 were activated by coexpression of NOX2, p22phox, p47phox and p67phox or NOX4 and p22phox (Figure 4(c)). Abundant mRNA expression of these subunits was verified by qRT-PCR (Figure 4(d)).
4. DISCUSSION
Oxidative stress is a cause of endothelial dysfunction, a prominent feature in the early stages of atherosclerosis [24]. Although increased ROS production and oxidant stress have been found to be involved in serum deprivation-induced endothelial cell injury, the enzymatic sources of ROS production and redox-signaling pathways are far from clear [11,12].
Serum deprivation from the HUVEC culture increased cellular ROS production and cell death via apoptotic mechanisms in the current study (Figure 1). The expression levels of NOX2, NOX4, and p47phox, but not p22phox and p67phox were increased by serum depletion. These results led us to examine the role of NOX2 versus NOX4 in ROS production and apoptotic endothelial cell death due to serum depletion.
Surprisingly, the effects of the siRNAs on NOX2 and NOX4 were quite different from each other (Figure 2). Serum depletion-induced ROS production was inhibited by NOX4 siRNA and no significant change was observed with the NOX2 siRNA. Furthermore, cell death and apoptotic caspase activation due to serum depletion were suppressed by NOX4 siRNA but they were rather increased by NOX2 siRNA. These results show that NOX4 may be the major source of ROS associated with the apoptotic death of endothelial cells under serumdepleted conditions. In contrast, ROS from NOX2, even though its amount is much less compared to that from NOX4, is assumed to play a role in attenuating cell death due to serum depletion.
How do different isotypes of NOX influence the cell fate differently? Of interest, NOX4 is known to generate hydrogen peroxide unlike other NOX isoforms generating superoxide anion radicals [15,16]. Superoxide mediates one-electron signaling while hydrogen peroxide
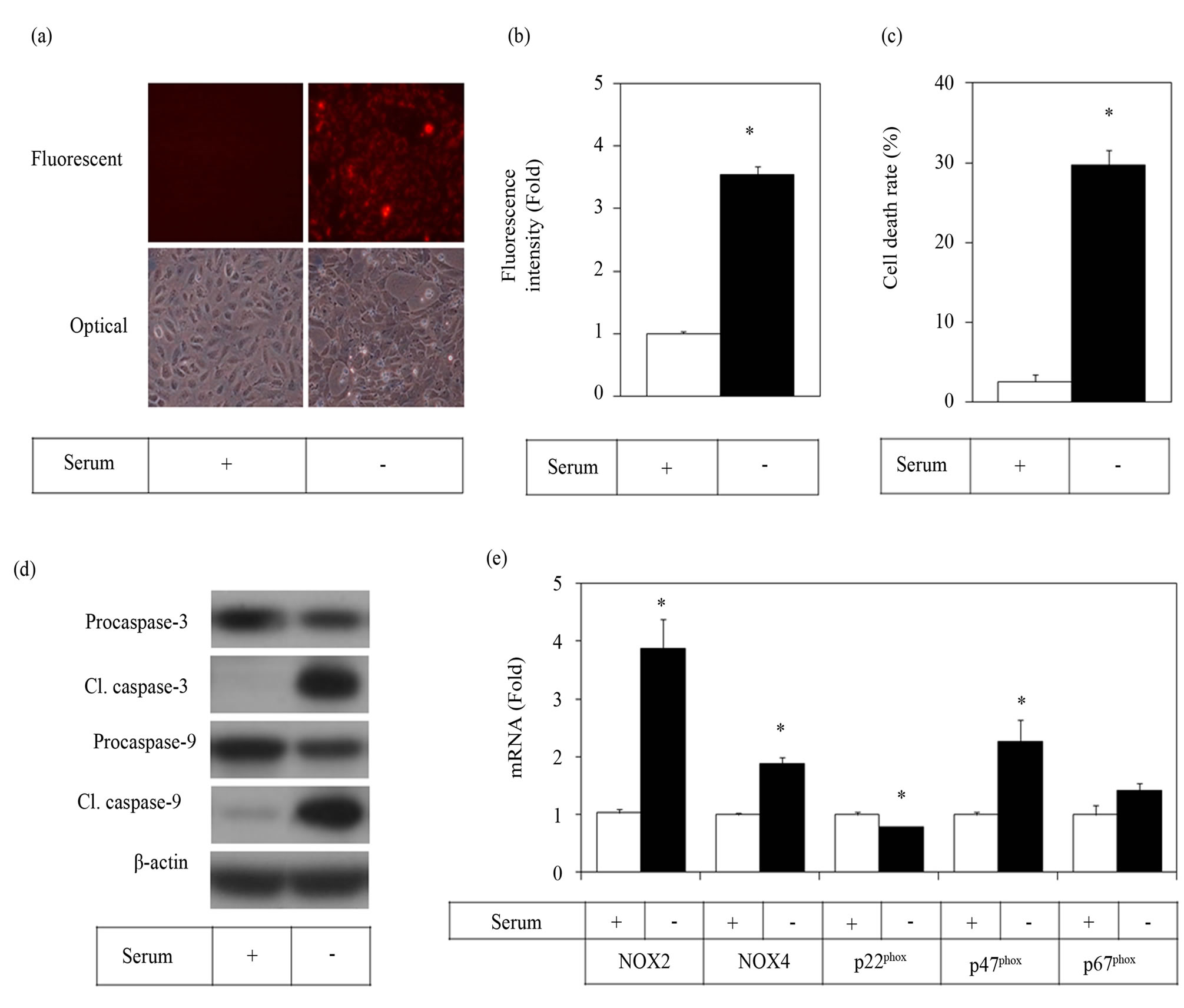
Figure 1. Serum depletion increased the ROS production and apoptotic cell death of endothelial cells. HUVECs were cultured under serum-provided or serum-depleted conditions. ROS production for 2 h was determined using the oxidant-sensitive probe DHR123. Fluorescent microscopic cell images (a) and fluorometric quantitation data are shown (b). Cell viability was assayed with trypan blue method after 24 h of serum depletion (c). Whole cell lysates were used for the Western blot of caspase-3 and caspase-9, both the inactive pro-forms and active cleaved forms (d). The mRNA expression of NOX2, NOX4, p22phox, p47phox and p67phox was determined by qRT-PCR (e). Data are presented as the Mean ± SE (n = 3). *p < 0.05 vs control.
mediates two-electron signaling [25], and their relative concentrations are known to be a key factor in determining cell fates, survival versus death [26]. In this regard, different NOX isoforms producing either superoxide or hydrogen peroxide preferentially would have different effects on cell fates. Thus, it is assumed that the preferential activation of NOX4 relative to NOX2 under serum depleted conditions increases the local concentration of hydrogen peroxide leading to cell death.
Based on the experimental results using siRNA for NOX2 and NOX4, we initially expected that overexpression of NOX4 might increase cell death and overexpression of NOX2 might have cytoprotective effects. However, we could not observe any significant changes in the ROS production and cell death rate by overexpression of exogenous NOX2 or NOX4 subunit alone (Figure 3). These results are different from previous reports [23]. One may argue that the transfection efficiency and assay sensitivity in our experiment were not enough to observe the difference caused by the overexpression of the exogenous NOX2 or NOX4 subunit. However, another thing to consider is that the activation of the exogenous NOX2 or NOX4 subunits may require stable organization by binding to p22phox or other subunits.
Indeed, coexpression of the 2 subunits of the NOX4 complex increased ROS production and cell death under
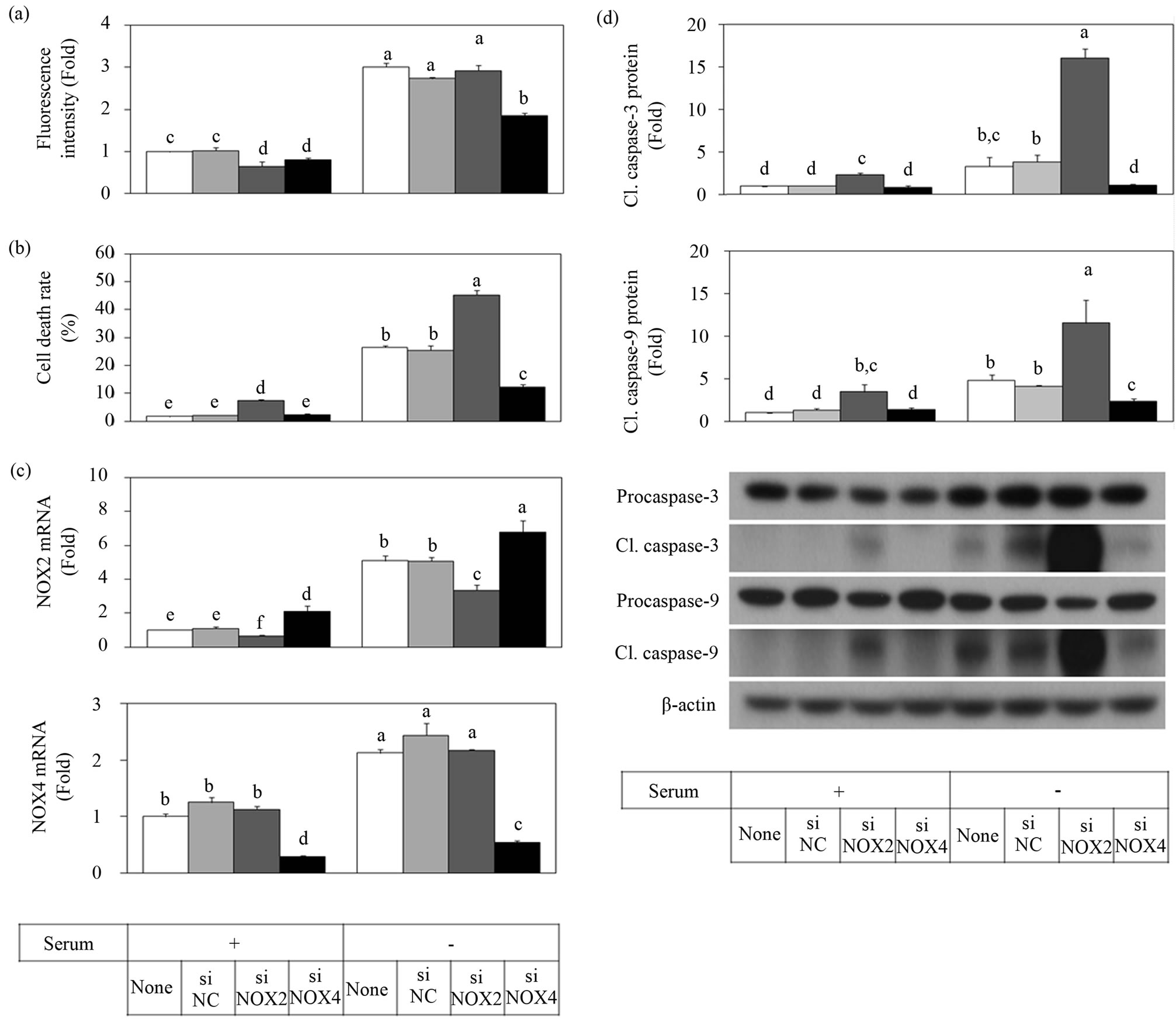
Figure 2. NOX4 siRNA attenuated ROS production and apoptotic cell death due to serum depletion whereas NOX2 siRNA increased apoptotic cell death. HUVECs were transfected with siNC, NOX2 siRNA, or NOX4 siRNA and cultured for 48 h, followed by 24 h serum depletion or not. ROS production for a 2 h serum depletion period was determined with the oxidant-sensitive fluorescence probe DHR123 (a). Cell death for a 24 h serum depletion period was determined with the trypan blue method (b). The mRNA expression of NOX2 and NOX4 was determined by qRT-PCR (c). Protein levels of caspase-3 and caspase-9 were determined by Western blot (d). Data are presented as the Mean ± SE (n = 3). Data not sharing the same letter differ significantly from each other (p < 0.05).
serum-provided and serum-depleted conditions (Figure 4), as we expected based on the data obtained using the NOX4 siRNA. However, coexpression of the 4 subunits of NOX2 complex also increased the ROS production and cell death under serum-provided and serum-depleted conditions (Figure 4), although we initially expected cytoprotective effects. These results show that ROS from both NOX4 and NOX2 would be cytotoxic if they are overproduced although a certain physiological range of superoxide anion radical produced by the NOX2 complex would be cytoprotective.
5. CONCLUSION
In conclusion, the present study showed that different NOX family members can have different effects on endothelial cell survival and death. NOX4-derived ROS appeared to mediate serum depletion-induced apoptotic cell death while a low level of ROS produced by NOX2 was implicated in providing an antiapoptotic mechanism. Thus, isotype-specific inhibition of NOX4 and activation of NOX2 are considered to be potentially useful strategies to enhance endothelial cell survival against apoptotic stimuli. However, special care must be paid in such an
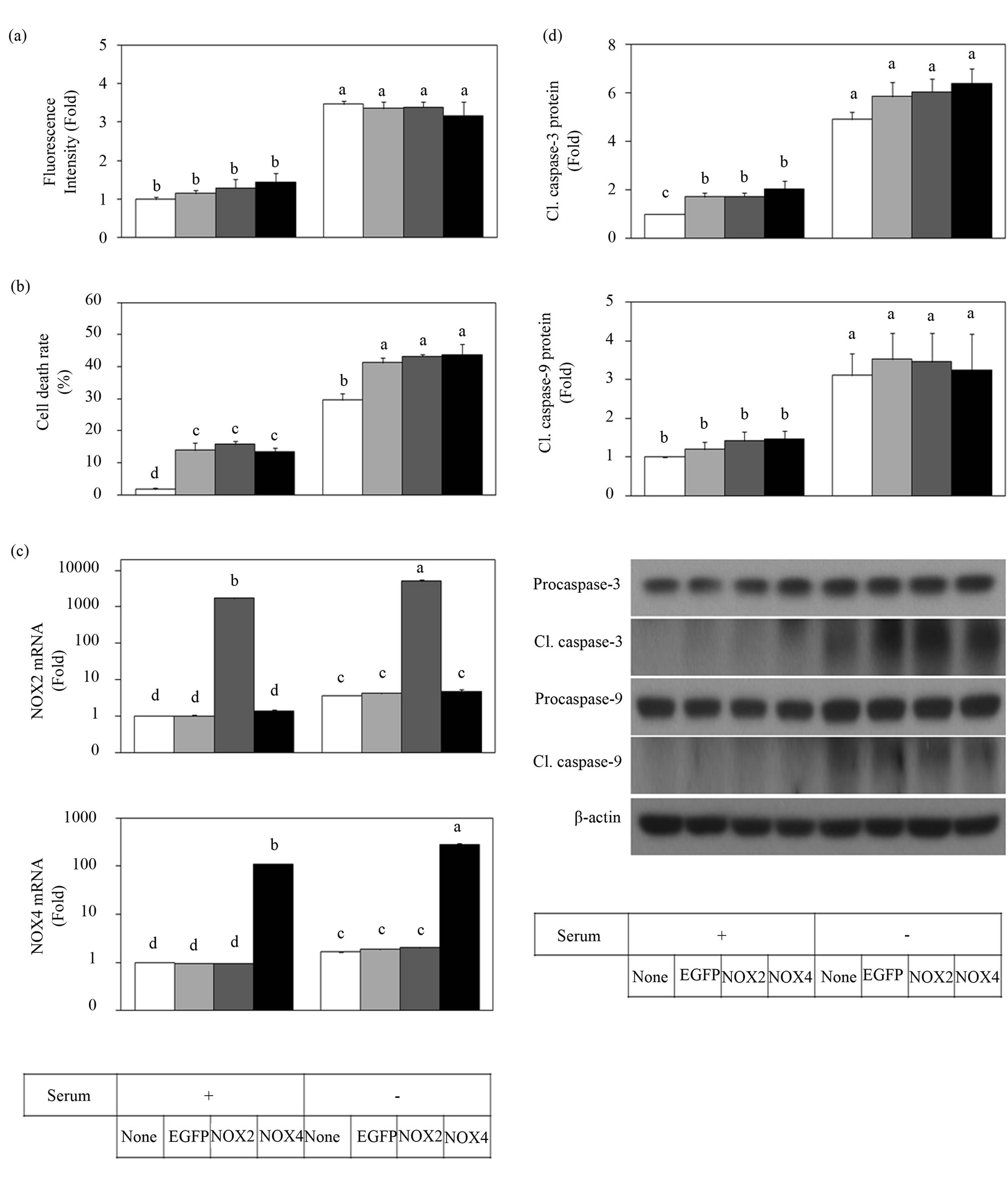
Figure 3. Over-expression of exogenous NOX2 or NOX4 subunit alone did not change ROS production and cell death under serum-provided and serum-depleted conditions. HUVECs were transfected with an equal amount of plasmid DNA encoding EGFP, NOX2 or NOX4, and cultured for 48 h, followed by 24 h of serum depletion or not. ROS production for a 2 h serum depletion period was determined with the oxidant-sensitive fluorescence probe DHR123 (a). Cell death after a 24 h serum depletion period was determined with the trypan blue method (b). The mRNA expression of NOX2 and NOX4 was determined by qRT-PCR and data are presented on a logarithmic scale (c). Protein levels of caspase-3 and caspase-9 were determined by Western blot (d). Data are presented as the Mean ± SE (n = 3). Data not sharing the same letter differ significantly from each other (p < 0.05).

Figure 4. Coexpression of the subunits of NOX2 or NOX4 complex increased ROS production and cell death under serum depletion. HUVECs were transfected with an equal amount plasmid DNAs and cultured for 48 h, followed by 24 h of serum depletion or not. ROS production for a 2 h serum depletion period was determined with the oxidant-sensitive fluorescence probe DHR123 (a). Cell death after a 24 h serum depletion period was determined with the trypan blue method (b). Protein levels of caspase-3 and caspase-9 were determined by western blot (c). The mRNA expression of NOX2, NOX4, p22phox, p47phox and p67phox was determined by qRTPCR and data are presented on a logarithmic scale (d). Data are presented as the Mean ± SE (n = 3). Data not sharing the same letter differ significantly from each other (p < 0.05).
approach because supraphysiological production of ROS from NOX2 as well as NOX4 could have harmful effects on cell survival.
ACKNOWLEDGEMENTS
This research was supported by the Mid-career Researcher Program (2011-0014527) of the National Research Foundation funded by the Ministry of Education, Science and Technology, Republic of Korea.
REFERENCES
- Gimbrone Jr., M.A. and Garcia-Cardena G. (2013) Vascular endothelium, hemodynamics, and the pathobiology of atherosclerosis. Cardiovascular Pathology, 22, 9-15. http://dx.doi.org/10.1016/j.carpath.2012.06.006
- Furchgott, R.F. and Zawadzki, J.V. (1980) The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature, 288, 373-376. http://dx.doi.org/10.1038/288373a0
- Libby, P. (2002) Inflammation in atherosclerosis. Nature, 420, 868-874. http://dx.doi.org/10.1038/nature01323
- Giannotti, G. and Landmesser U. (2007) Endothelial dysfunction as an early sign of atherosclerosis. Herz Kardiovaskuläre Erkrankungen, 32, 568-572. http://dx.doi.org/10.1007/s00059-007-3073-1
- Cai, H. and Harrison, D.G. (2000) Endothelial dysfunction in cardiovascular diseases: the role of oxidant stress. Circulation Research, 87, 840-844. http://dx.doi.org/10.1161/01.RES.87.10.840
- Hadi, H.A., Carr, C.S. and Al Suwaidi, J. (2005) Endothelial dysfunction: Cardiovascular risk factors, therapy, and outcome. Vascular Health and Risk Management, 1, 183-198.
- Dimmeler, S., Hermann C. and Zeiher, A.M. (1998) Apoptosis of endothelial cells. Contribution to the pathophysiology of atherosclerosis? European Cytokine Network, 9, 697-698.
- Duval, H., Harris, M., Li, J., Johnson, N. and Print, C. (2003) New insights into the function and regulation of endothelial cell apoptosis. Angiogenesis, 6, 171-183. http://dx.doi.org/10.1023/B:AGEN.0000021390.09275.bc
- Tricot, O., Mallat, Z., Heymes, C., Belmin, J., Leseche, G. and Tedgui A. (2000) Relation between endothelial cell apoptosis and blood flow direction in human atherosclerotic plaques. Circulation, 101, 2450-2453. http://dx.doi.org/10.1161/01.CIR.101.21.2450
- Kono, Y., Sawada, S., Kawahara, T., Tsuda, Y., Higaki, T., Yamasaki, S., Imamura, H., Tada, Y., Sato, T., Hiranuma, O., Akamatsu, N., Komatsu, S., Tamagaki, T., Nakagawa, K., Tsuji, H. and Nakagawa, M. (2002) Bradykinin inhibits serum-depletion-induced apoptosis of human vascular endothelial cells by inducing nitric oxide via calcium ion kinetics. Journal of Cardiovascular Pharmacology, 39, 251-261. http://dx.doi.org/10.1097/00005344-200202000-00012
- Aoki, M., Nata, T., Morishita, R., Matsushita, H., Nakagami, H., Yamamoto, K., Yamazaki, K., Nakabayashi, M., Ogihara, T. and Kaneda, Y. (2001) Endothelial apoptosis induced by oxidative stress through activation of NFkappaB: Antiapoptotic effect of antioxidant agents on endothelial cells. Hypertension, 38, 48-55. http://dx.doi.org/10.1161/01.HYP.38.1.48
- Li, J.M., Fan, L.M., George, V.T. and Brooks, G. (2007) NOX2 regulates endothelial cell cycle arrest and apoptosis via p21cip1 and p53. Free Radical Biology & Medicine, 43, 976-986. http://dx.doi.org/10.1016/j.freeradbiomed.2007.06.001
- Bedard, K. and Krause, K.H. (2007) The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiological Reviews, 87, 245-313. http://dx.doi.org/10.1152/physrev.00044.2005
- Lassegue, B. and Griendling, K.K. (2010) NADPH oxidases: Functions and pathologies in the vasculature. Arteriosclerosis, Thrombosis, and Vascular Biology, 30, 653- 661. http://dx.doi.org/10.1161/ATVBAHA.108.181610
- Martyn, K.D., Frederick, L.M., von Loehneysen, K., Dinauer, M.C. and Knaus, U.G. (2006) Functional analysis of Nox4 reveals unique characteristics compared to other NADPH oxidases. Cell Signaling, 18, 69-82. http://dx.doi.org/10.1016/j.cellsig.2005.03.023
- Dikalov, S.I., Dikalova, A.E., Bikineyeva, A.T., Schmidt, H.H., Harrison, D.G. and Griendling, K.K. (2008) Distinct roles of Nox1 and NOX4 in basal and angiotensin II-stimulated superoxide and hydrogen peroxide production. Free Radical Biology & Medicine, 45, 1340-1351. http://dx.doi.org/10.1016/j.freeradbiomed.2008.08.013
- Irani, K. (2000) Oxidant signaling in vascular cell growth, death, and survival: A review of the roles of reactive oxygen species in smooth muscle and endothelial cell mitogenic and apoptotic signaling. Circulation Research, 87, 179-183. http://dx.doi.org/10.1161/01.RES.87.3.179
- Basuroy, S., Bhattacharya, S., Leffler, C.W. and Parfenova, H. (2009) NOX4 NADPH oxidase mediates oxidative stress and apoptosis caused by TNF-alpha in cerebral vascular endothelial cells. American Journal of Physiology-Cell Physiology, 296, C422-C432. http://dx.doi.org/10.1152/ajpcell.00381.2008
- Peshavariya, H., Dusting, G.J., Jiang, F., Halmos, L.R., Sobey, C.G., Drummond, G.R. and Selemidis, S. (2009) NADPH oxidase isoform selective regulation of endothelial cell proliferation and survival. Naunyn-Schmiedeberg’s Archives of Pharmacology, 380, 193-204. http://dx.doi.org/10.1007/s00210-009-0413-0
- Deshpande, S.S., Angkeow, P., Huang, J., Ozaki, M. and Irani, K. (2000) Rac1 inhibits TNF-alpha-induced endothelial cell apoptosis: Dual regulation by reactive oxygen species. The FASEB Journal, 14, 1705-1714. http://dx.doi.org/10.1096/fj.99-0910com
- Mochizuki, T., Furuta S., Mitsushita J., Shang, W.H., Ito, M., Yokoo, Y., Yamaura, M., Ishizone, S., Nakayama, J., Konagai, A., Hirose, K., Kiyosawa, K. and Kamata T. (2006) Inhibition of NADPH oxidase 4 activates apoptosis via the AKT/apoptosis signal-regulating kinase 1 pathway in pancreatic cancer PANC-1 cells. Oncogene, 25, 3699-3707. http://dx.doi.org/10.1038/sj.onc.1209406
- Jeon, H. and Boo, Y.C. (2013) Laminar shear stress enhances endothelial cell survival through a NADPH oxidase 2-dependent mechanism. Biochemical and Biophysical Research Communications, 430, 460-465. http://dx.doi.org/10.1016/j.bbrc.2012.12.016
- Petry, A., Djordjevic, T., Weitnauer, M., Kietzmann, T., Hess, J. and Gorlach, A. (2006) NOX2 and NOX4 mediate proliferative response in endothelial cells. Antioxidants & Redox Signaling, 8, 1473-1484. http://dx.doi.org/10.1089/ars.2006.8.1473
- Valgimigli, M., Merli, E., Malagutti, P., Soukhomovskaia, O., Cicchitelli, G., Macri, G. and Ferrari, R. (2003) Endothelial dysfunction in acute and chronic coronary syndromes: Evidence for a pathogenetic role of oxidative stress. Archives of Biochemistry and Biophysics, 420, 255-261. http://dx.doi.org/10.1016/j.abb.2003.07.006
- Buettner, G.R., Ng, C.F., Wang, M., Rodgers, V.G. and Schafer, F.Q. (2006) A new paradigm: Manganese superoxide dismutase influences the production of H2O2 in cells and thereby their biological state. Free Radical Biology & Medicine, 41, 1338-1350. http://dx.doi.org/10.1016/j.freeradbiomed.2006.07.015
- Clement, M.V. and Pervaiz, S. (2001) Intracellular superoxide and hydrogen peroxide concentrations: A critical balance that determines survival or death. Redox Report, 6, 211-214. http://dx.doi.org/10.1016/j.freeradbiomed.2006.07.015