V. Singh et al. / Natural Science 2 (2010) 1148-1154
Copyright © 2010 SciRes. OPEN ACCESS
115
1153
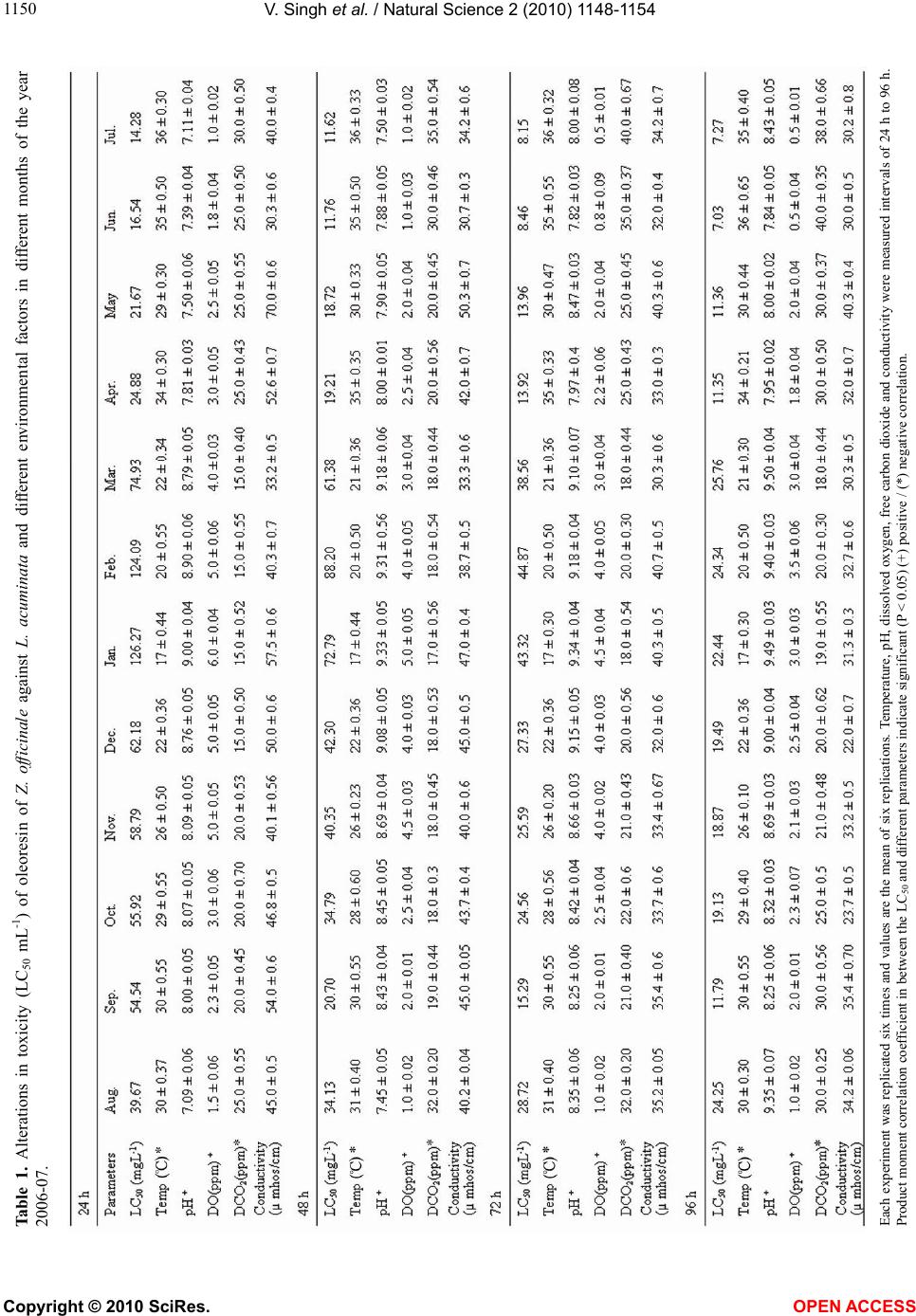
or in the snail body due to variant environmental factors
in the month of June and July. Earlier, it has been shown
that the treatment of oleoresin of Z. officinale caused
significant inhibition of AChE, ALP and ACP activity in
the nervous tissue of L. acuminata [9]. The high anti
AChE and ACP activity of oleoresin of Z. officinale was
observed in months of June-July. The enzyme ALP plays
a critical role in protein synthesis [36] and secretary ac-
tivity [37] is comparatively less inhibited than AChE.
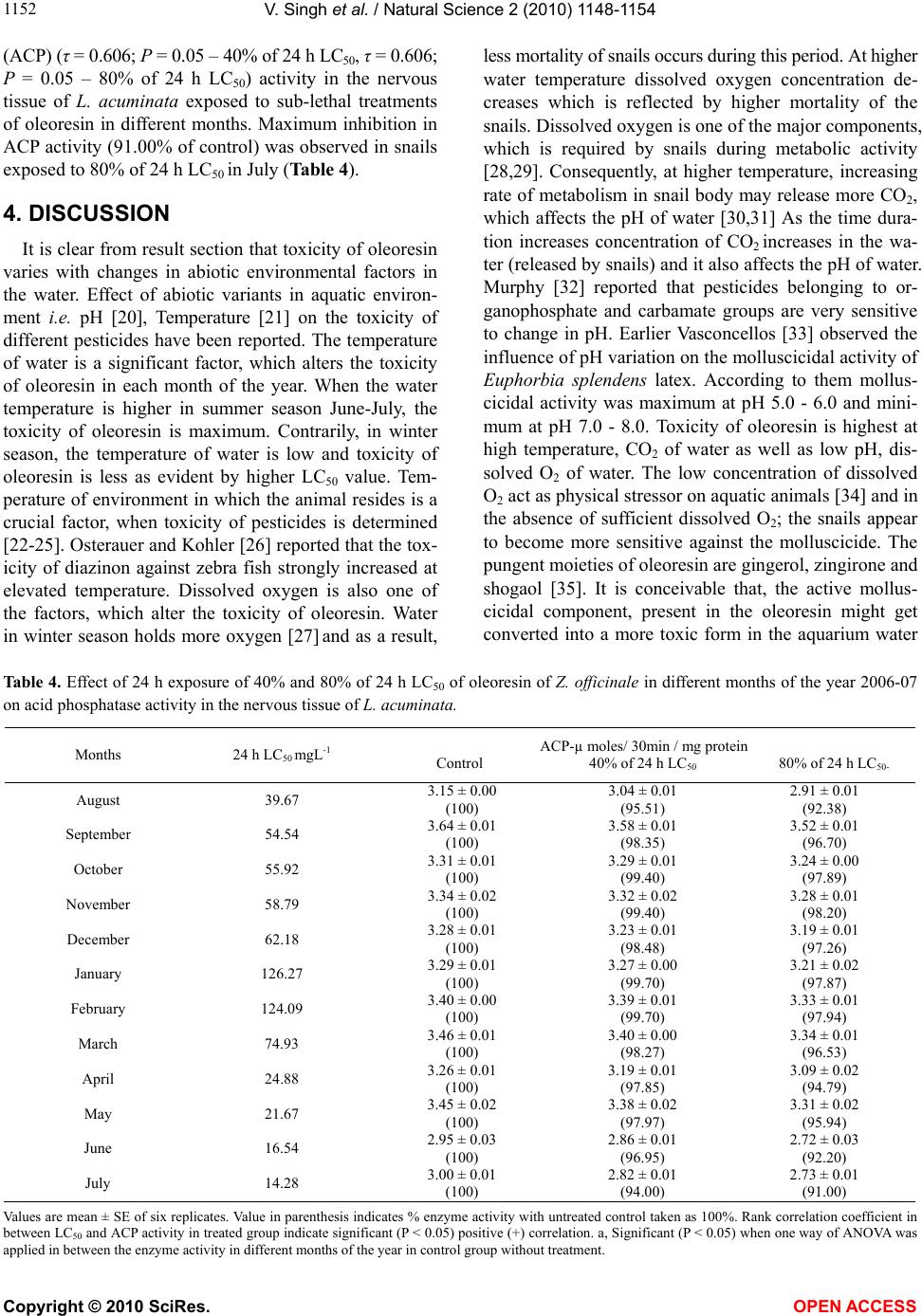
Acid phosphatase (ACP), a lysosomal enzyme [38], plays
an important role in autolysis and phagocytosis, patho-
logical necrosis, and overall catabolism [8,10,39] was
reduced significantly. Earlier, it has been observed that
increased activity of ACP causes breakdown of existing
protein in L. acuminata [18], but inhibition of ACP ac-
tivity in this study indicates that it is not used in break-
down of cellular protein. The rank correlation coefficient
applied between the LC50 values of different months and
the corresponding inhibition in enzyme activity, point
out a positive correlation between the LC50 and the inhi-
bition of AChE and ACP. Whereas there was no correla-
tion in between LC50 and ALP activity indicate that ALP
is not altered by action of oleoresin in different months.
Accurate prediction of molluscicide fate and toxicity
in aqueous environment against snails are hindered due
to lack of information that how abiotic factors of aque-
ous environment affect the biological activity and related
toxicity of molluscicides. Abiotic factors are not only
correlated with the lethality of molluscicide, but with
each other also. The present study conclusively shows
that variant abiotic factors can significantly alter the
toxicity of oleoresin of Z. officinale in L. acuminata. It is
also obvious that the most suitable period for the control
of this snail in India is the month of June, July. It is sug-
gested that the treatment of a water body with oleoresin
of Z. officinale for the control of L. acuminata and ulti-
mately fascioliasis, is not only more potent and cost ef-
fective during these months than spending more money
by using higher concentrations of this molluscicide dur-
ing the rest ten months of the year.
REFERENCES
[1] Singh, S., Singh, V.K. and Singh, D.K. (1997) Mollus-
cicidal activity of some common spice plants. Biological
Agriculture and Horticulture, 14, 237-249.
[2] Shukla, S., Singh, V.K. and Singh, D.K. (2006) The ef-
fect of single, binary, tertiary, combination of few plant
derived molluscicides alone or in combination with syn-
ergist on different enzymes in the nervous tissues of the
freshwater snail Lymnaea (Radix) acuminata (Lamark).
Pesticide Biochemistry and Physiology, 85, 167-173.
[3] Singh, O. and Agarwal, R.A. (1981) Toxicity of certain
pesticides to two economic species of snails in northern
India. Journal of Economic Entomology, 74, 568-571.
[4] World Health Organization. (2006) Report of the WHO
informal meeting on use of triclabendazole in fascioliasis
control. WHO headquarters, Geneva, Switzerland, 17-18
October 2006.
[5] Singh, P., Singh, V.K. and Singh, D.K. (2005) Effect of
binary combination of some plant-derived molluscicides
with MGK-264 or piperonyl butoxide on the reproduc-
tion of the snail Lymnaea acuminata. Pest Management
Science, 61, 204-208.
[6] Kumar, P. and Singh, D.K. (2006) Molluscicidal activity
of Ferula asafoetida, Syzygium aromaticum and Carum
carvi and their active components against the snail Ly m-
naea acuminata. Chemo, 63, 1568-1574.
[7] Kumar, P., Singh, V.K. and Singh, D.K. (2009) Kinetics
of enzyme inhibition by active molluscicidal agents fer-
ulic acid, umbelliferone, eugenol and limonene in the
nervous tissue of snail Lymnaea acuminata. Phytother-
apy Research, 23, 172-177.
[8] Jaiswal, P., Singh, V.K. and Singh, D.K. (2008) Enzyme
inhibition by molluscicidal component of Areca catechu
and Carica papaya in the nervous tissue of vector snail
Lymnaea acuminata. Pesticide Biochemistry and Physiol-
ogy, 92, 164-168.
[9] Singh, K. and Singh, D.K. (2000) Toxicity to the snail
Lymnaea acuminata of plant-derived molluscicides in
combination with synergists. Pest Management Science,
56, 889-898.
[10] Tripathi, S.M., Singh, V.K., Singh, S. and Singh, D.K.
(2004) Enzyme inhibition by the molluscicidal agent Pu-
nica granatum Linn. bark and Canna indica Linn. root.
Phytotherapy Research, 18, 501-506.
[11] Gunther, E. (1975) The essential oils. Robert, E Krieger
Publishing Company, NewYork.
[12] American Public Health Association (APHA). (2005) Stan-
dard methods for the examination of water and wastewa-
ter. 21st Edition, Washington, D.C.
[13] Robertson, J.L., Russell, R.M., Preisler, H.K. and Savin,
N.E. (2007) Bioassay with Arthropods POLO computer
programme for analysis of bioassay data. 2nd Edition,
CRC Press, Talor and Francis, 1-224.
[14] Sokal, R.R. and Rohlf, F.J. (1973) Introduction to biosta-
tistics. W. H. Freeman & Co., Ltd., San Francisco.
[15] Ellman, G.L., Courtney, K.D., Andres, V. and Feather-
stone, R.M. (1961) A new and rapid colorimetric deter-
mination of acetylcholinesterase activity. Bio Pharmacol,
7, 88-95.
[16] Singh, D.K., Singh, O. and Agarwal, R.A. (1982) Com-
parative study of cholinesterase in two snails. Pila glo-
bosa and Lymnaea acuminata. The Journal of Physiology,
78, 467-472.
[17] Bergmeyer, U.H. (1967) Methods of enzymatic analysis.
Academic Press, New York, 1129.
[18] Singh, D.K. and Agarwal, R.A. (1989) Toxicity of pi-
peronyl butoxide carbaryl synergism on the snail Lym-
naea acuminata. Internationale Revue der gesamten Hy-
drobiologie und Hydrographie, 74, 689-699.
[19] Lowry, O.H., Rosebrough, N.J., Farr, A.L. and Randall
R.J. (1951) Protein measurement with folin phenol rea-
gent. The Journal of Biological Chemistry, 193, 265-275.
[20] Watson, S.J. and Maly, E.J. (1987) Thiocyanate toxicity
to Daphina magna: Modified by pH and temperature. Aqua-