Estimates of Genetic Variability of Mycobacterium tuberculosis Complex and
Its Association with Drug Resistance in Cameroon
59
The population structure of Mycobacterium tuberculo-
sis complex strains from the Centre region was diverse
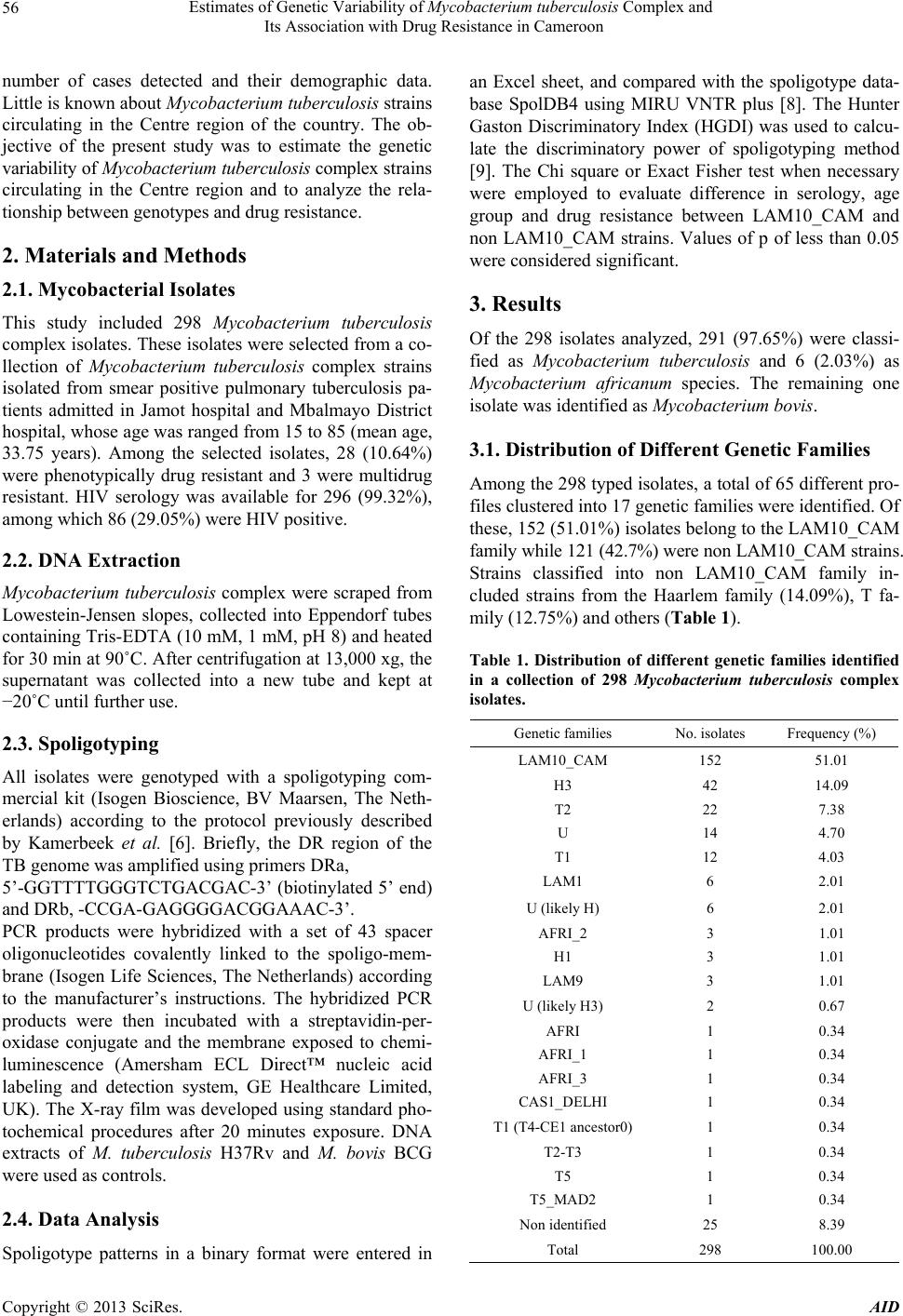
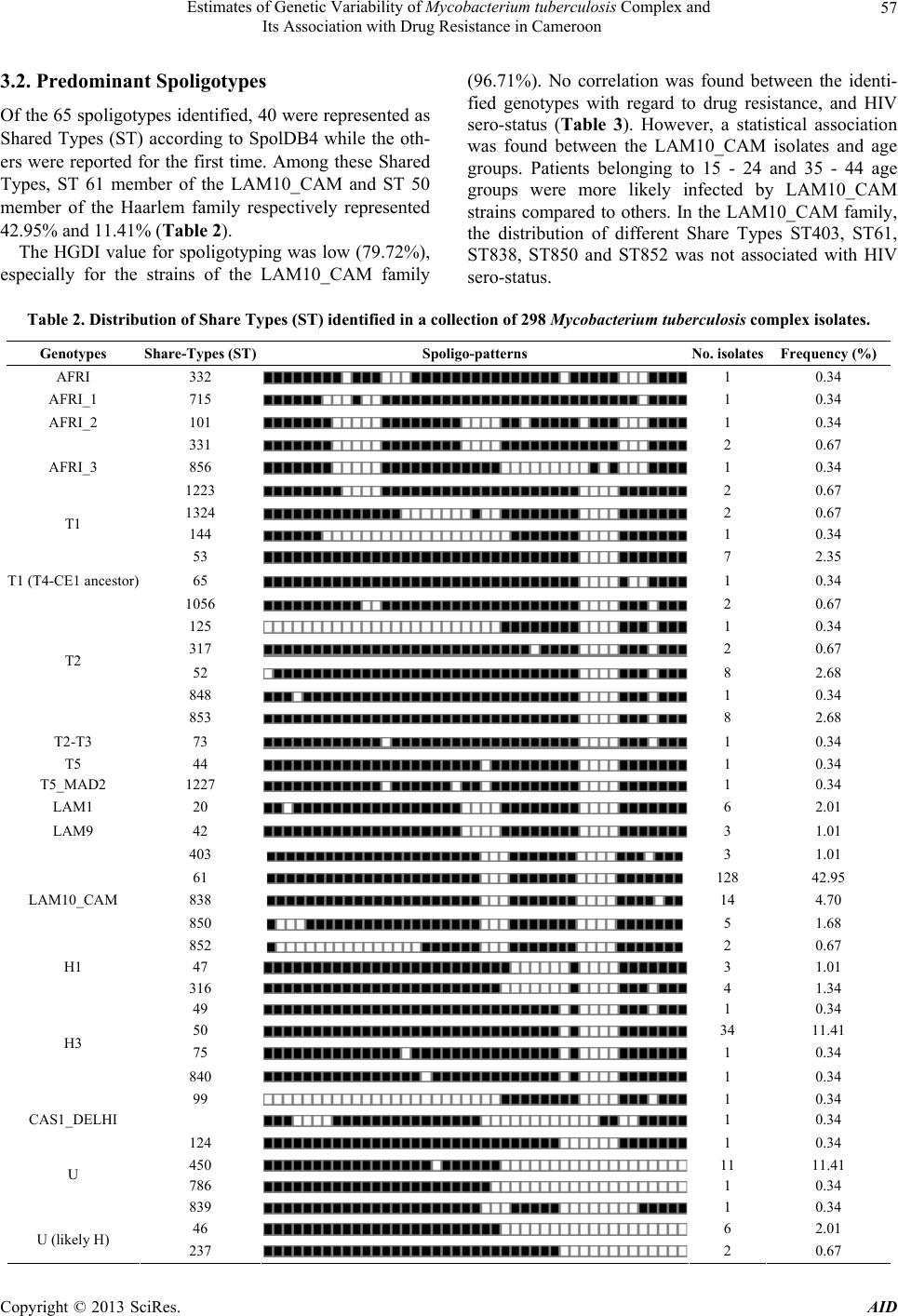
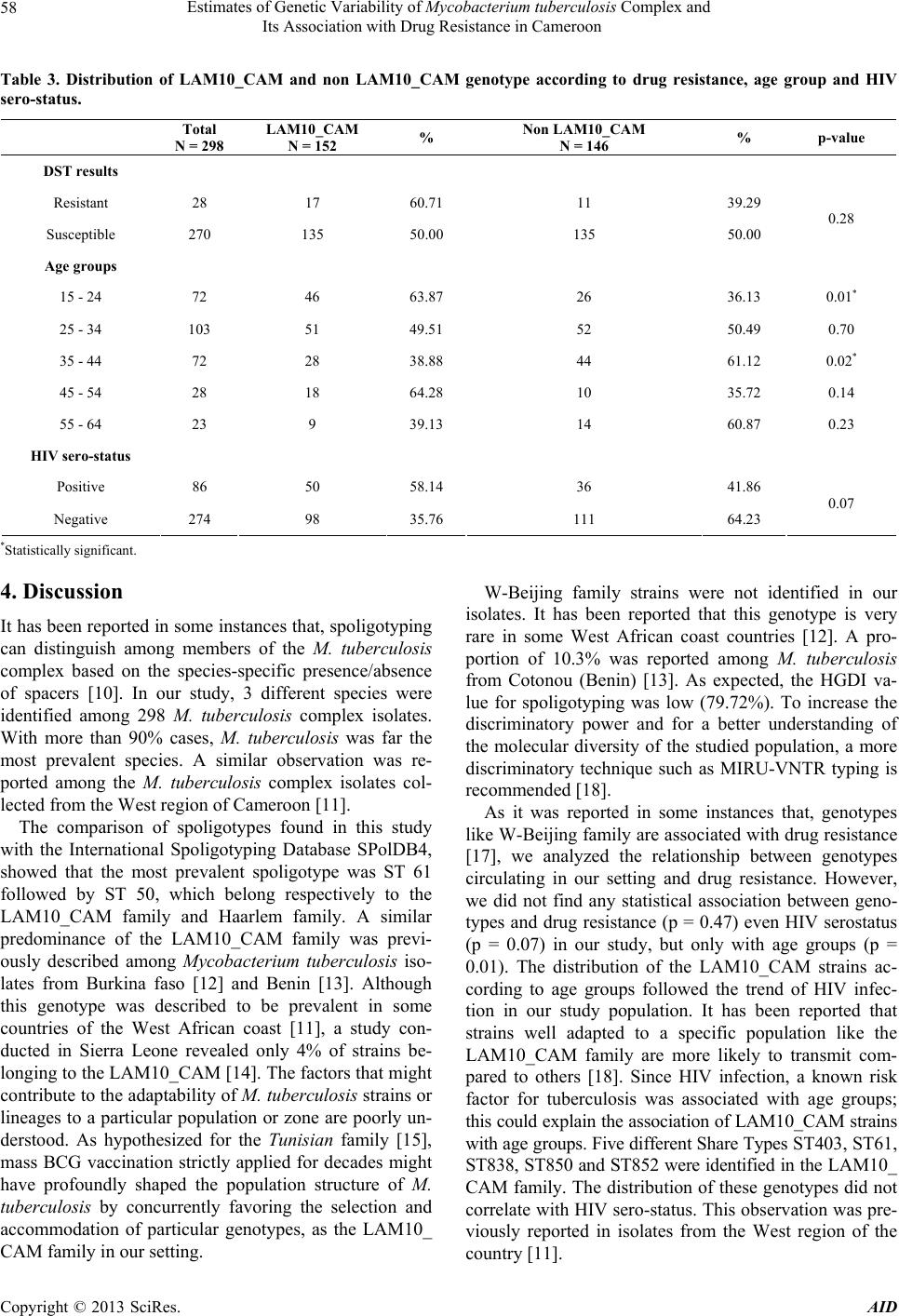
and included 65 different genotypes. The majority of
strains belonged to the LAM10_CAM which can be sub-
divided in 5 spoligotypes. The consequence of this diver-
sity for the TB epidemic are not yet clear and need to be
addressed in further studies.
5. Acknowledgements
This study was financially supported by EDCTP grant
through the CANTAM-TB “Central Africa Network for
Tuberculosis, AIDS/HIV and Malaria” project. Larissa
Kamgue Sidze and Emmanuel Mouafo Tekwu were re-
search fellow students at the Institute for Tropical Medi-
cine in Tübingen (Germany). We thank Mrs. Augusta
Tsasse (Centre Pasteur of Cameroon) and Tanja Ubben
(Research Centre Borstel, Germany) for their technical
assistance.
REFERENCES
[1] C. Dye, “Global Epidezmiology of Tuberculosis,” Lancet,
Vol. 367, No. 9514, 2006, pp. 938-940.
doi:10.1016/S0140-6736(06)68384-0
[2] K. Rowland, “Totally Drug-Resistant TB Emerges in
India,” Nature News and Comment, 13 January 2012.
[3] World Health Organization, “Global Tuberculosis Control:
WHO Report 2011,” Geneva, 2011.
[4] G. D. Van der Spuy, R. M. Warren and P. D. Van Helden,
“The Role of Molecular Epidemiology in Low-Income,
High-Burden Countries,” International Journal of Tuber-
cle and Lung Diseases, Vol. 13, No. 4, 2009, pp. 419-
420.
[5] D. Van Soolingen, P. W. Hermans, P. E. De Haas, D. R.
Soll and J. D. Van Embden, “Occurrence and Stability of
Insertion Sequences in Mycobacterium tuberculosis Com-
plex Strains: Evaluation of an Insertion Sequence-De-
pendent DNA Polymorphism as a Tool in the Epidemiol-
ogy of Tuberculosis,” Journal of Clinical Microbiology,
Vol. 29, No. 11, 1991, pp. 2578-2586.
[6] J. Kamerbeek, L. Schouls, A. Kolk, M. Van Agterveld, D.
Van Soolingen, S. Kuijper, A. Bunschoten, H. Molhuizen,
R. Shaw, M. Goyal, et al., “Simultaneous Detection and
Strain Differentiation of Mycobacterium tuberculosis for
Diagnosis and Epidemiology,” Journal of Clinical Mi-
crobiology, Vol. 35, No. 4, 1997, pp. 907-914.
doi:10.3201/eid0811.020125
[7] I. Filliol, J. R. Driscoll, D. Van Soolingen, B. N. Kreis-
wirth, K. Kremer, G. Valetudie, D. D. Anh, R. Barlow, D.
Banerjee, P. J. Bifani, et al., “Global Distribution of My-
cobacterium tuberculosis Spoligotypes,” Emerging Infec-
tious Diseases, Vol. 8, No. 11, 2002, pp. 1347-1349.
[8] T. Weniger, J. Krawczyk, P. Supply, S. Niemann and D.
Harmsen, “MIRU-VNTR plus: A Web Tool for Polypha-
sic Genotyping of Mycobacterium tuberculosis Complex
Bacteria,” Nucleic Acids Research, Vol. 38, 2010, pp.
W326-W331.
[9] P. R. Hunter and M. A. Gaston, “Numerical Index of the
Discriminatory Ability of Typing Sysems: An Applica-
tion of Simpson’s Index of Diversity,” Journal of Clinical
Microbiology, Vol. 26, No. 1, 1988, pp. 2465-2466.
[10] B. Mathema, N. E. Kurepina, P. J. Bifani and B. N. Krei-
swirth, “Molecular Epidemiology of Tuberculosis: Cur-
rent Insights,” Clinical Microbiology Review, Vol. 19, No.
4, 2006, pp. 658-685. doi:10.1128/CMR.00061-05
[11] S. N. Niobe-Eyangoh, C. Kuaban, P. Sorlin, P. Cunin, J.
Thonnon, C. Sola, et al., “Genetic Biodiversity of Myco-
bacterium tuberculosis Complex Strains from Patients
with Pulmonary Tuberculosis in Cameroon,” Journal of
Clinical Microbiology, Vol. 41, No. 6, 2003, pp. 2547-
2553. doi:10.1128/JCM.41.6.2547-2553.2003
[12] S. Godreuil, G. Torrea, D. Terru, F. Chevenet, S. Diag-
bouga, P. Supply, et al., “First Molecular Epidemiology
Study of Mycobacterium tuberculosis in Burkina Faso,”
Journal of Clinical Microbiology, Vol. 45, No. 3, 2007,
pp. 921-927. doi:10.1128/JCM.01918-06
[13] D. Affolabi, G. Anyo, F. Faihun, N. Sanoussi, I. C. Sham-
puta, L. Rigouts, et al., “First Molecular Epidemiological
Study of Tuberculosis in Benin,” International Journal of
Tubercle and Lung Diseases, Vol. 13, No. 13, 2009, pp.
317-322.
[14] S. Homolka, E. Post, B. Oberhauser, A. G. George, L.
Westman, F. Dafae, S. Rusch-Gerdes and S. Niemann,
“High Genetic Diversity among Mycobacterium tubercu-
losis Complex Strains from Sierra Leone,” BMC Micro-
biology, Vol. 8, No. 103, 2008, pp. 489-495.
[15] A. Namouchi, A. Karboul, B. Mhenni, N. Khabouchi, R.
Haltiti, R. Ben Hassine, et al., “Genetic Profiling of My-
cobacterium tuberculosis in Tunisia: Predominance and
Evidence for the Establishment of a Few Genotypes,”
Journal of Medical Microbiology, Vol. 57, No. 7, 2008,
pp. 864-872. doi:10.1099/jmm.0.47483-0
[16] P. Supply, E. Mazars, S. Lesjean, V. Vincent, B. Gicquel
and C. Locht, “Variable Human Minisatellite-Like Re-
gions in the Mycobacterium tuberculosis Genome,” Mo-
lecular Microbiology, Vol. 36, No. 3, 2000, pp. 762-771.
doi:10.1046/j.1365-2958.2000.01905.x
[17] W. A. Githui, A. M. Jordaan, E. S. Juma, P. Kinyanjui, F.
G. Karimi, J. Kimwomi, et al., “Identification of MDR-
TB Beijing/W and Other Mycobacterium tuberculosis
Genotypes in Nairobi, Kenya,” International Journal of
Tubercle and Lung Diseases, Vol. 8, No. 3, 2004, pp.
352-360.
[18] S. Gagneux and P. M. Small, “Global Phylogeography of
Mycobacterium tuberculosis and Implications for Tuber-
culosis Product Development,” Lancet Infectious Dis-
eases, Vol. 7, No. 5, 2007, pp. 328-337.
doi:10.1016/S1473-3099(07)70108-1
Copyright © 2013 SciRes. AID