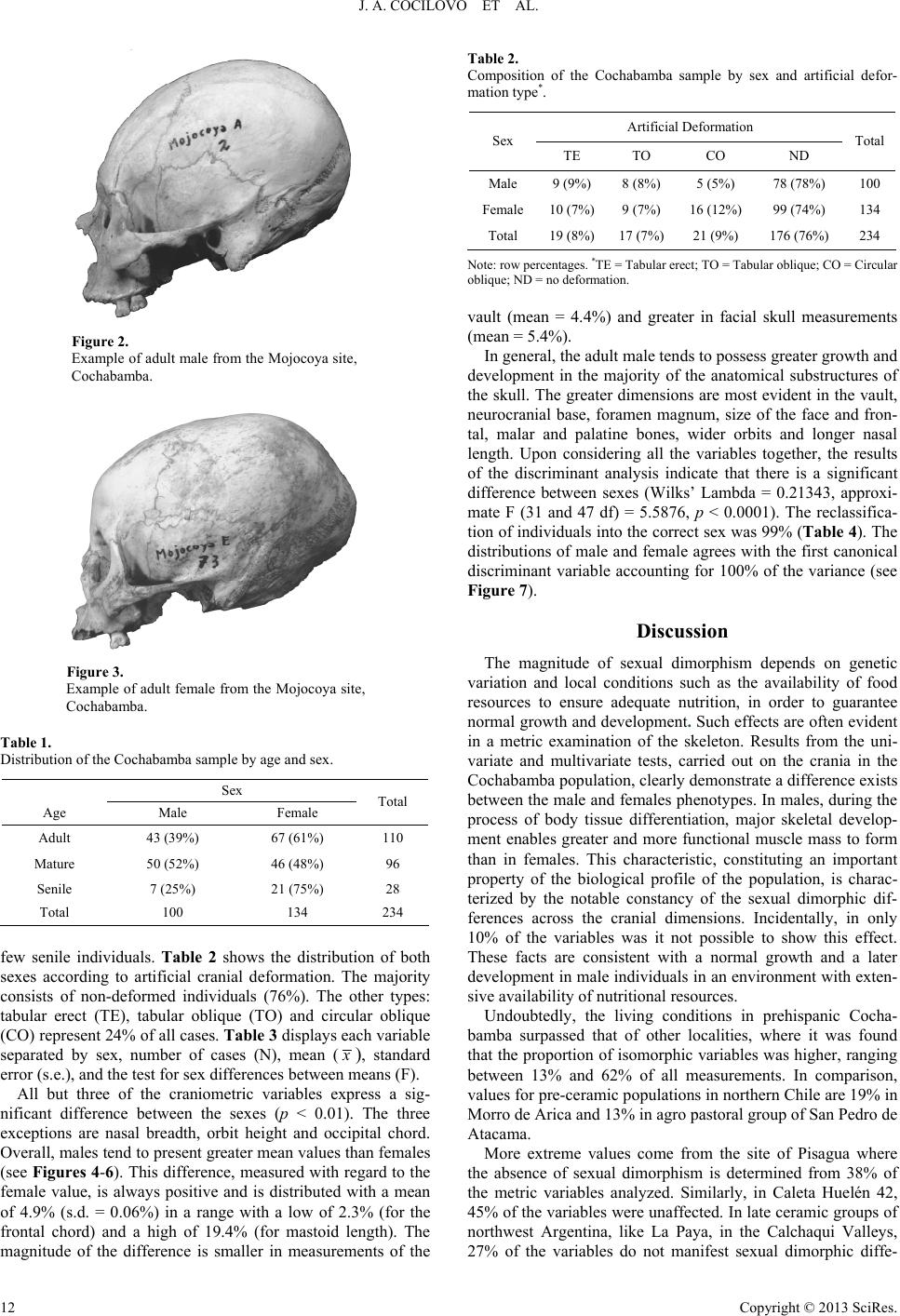
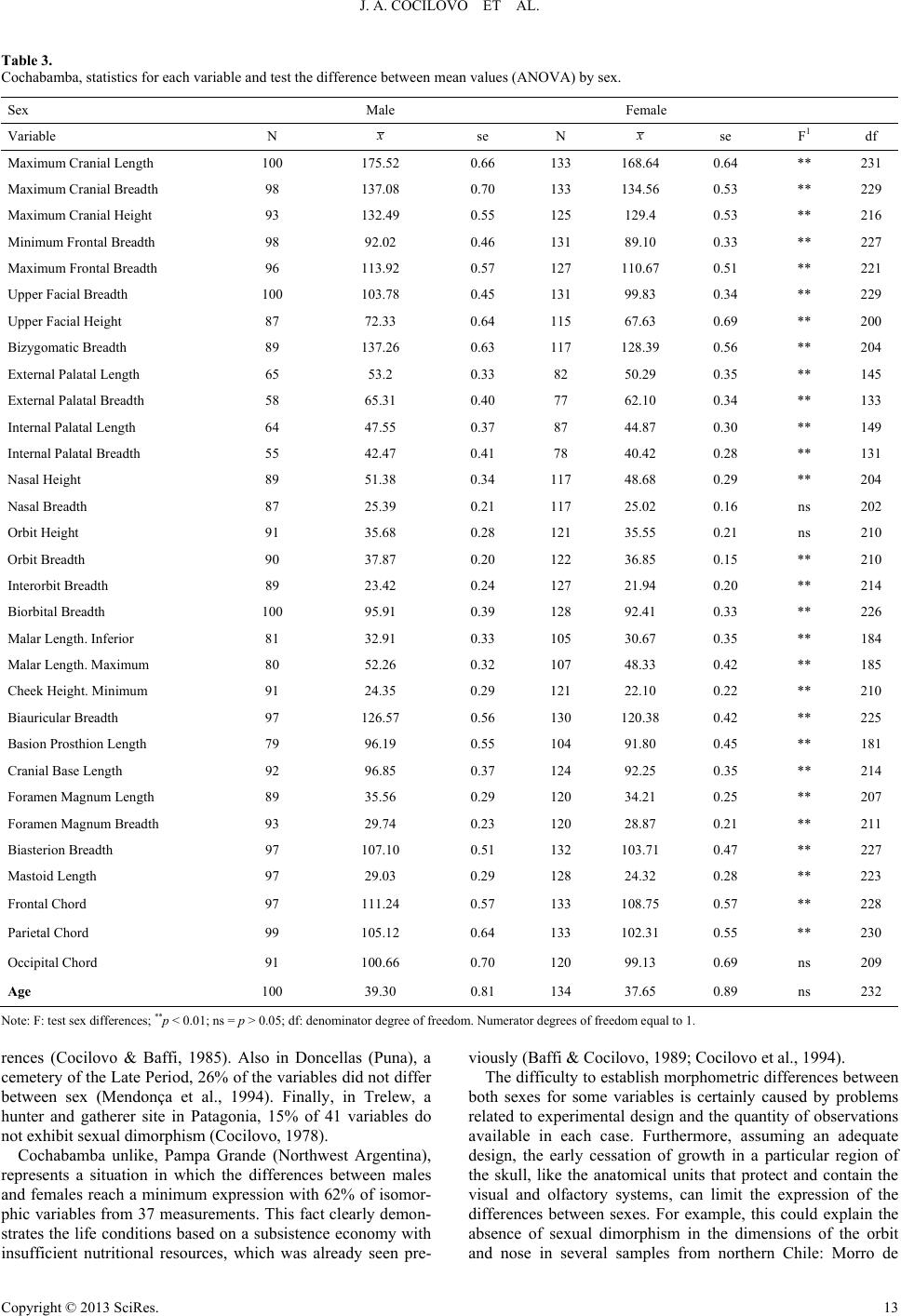
J. A. COCILOVO ET AL.
Copyright © 2013 SciRes. 15
ciate the access to the skeletal collections granted by David
Pereira and Ramon Sanzetenea (Museo Arqueológico del Uni-
versidad Mayor de San Simon, Cochabamba, Bolivia) and Ed-
mundo Salinas (Museo Arqueológico de la Universidad de
Chuquisaca, Sucre, Bolivia).
REFERENCES
Acsádi, G., & Nemeskéri, J. (1970). History of human life span and
mortality. Budapest: Akadémiai Kiadó.
Bass, W. M. (1987). Human osteology: A laboratory and field manual
of the human skeleton. Springfield: University of Missouri.
Buikstra, J. E., & Ubelaker, D. H. (1994). Standards for data collection
from human skeletal remains. Fayetteville: Arkansas Archaeological
Survey.
Baffi, E. I., & Cocilovo, J. A. (1989). Evaluación del impacto ambiental
en una población prehistórica: El caso de Las Pirguas (Salta, Argen-
tina). Revista Argentina de Antropologia Biologica, 4, 39-43.
Baffi, E. I., Torres, M. F., & Cocilovo, J. A. (1996). La población
prehispánica de Las Pirguas (Salta, Argentina). Un enfoque integral.
Revista Argentina de Antropologia Biologica, 1, 204-218.
Cocilovo, J. A. (1975). Estudio de dos factores que influencian la
morfología craneana en una colección andina: El sexo y la defor-
mación artificial. Revista Instituto de A nt ro po l og ía , 2 , 197-212.
Cocilovo, J. A. (1978). Estudio de dos factores que influyen en la
morfología craneana en una colección patagónica: El sexo y la de-
formación artificial. Arquivos de Anatomía e Arqueología, 3, 113-141.
Cocilovo, J. A., & Baffi, E. I. (1985). Contribución al conocimiento de
las características biológicas de la población prehistórica de Puerta
de La Paya (Salta). Runa, 15, 153-178.
Cocilovo, J. A., & Marcellino, A. J. (1974). Determinación del sexo en
una serie craneana aborigen por medio de las funciones discri-
minantes. Revista Instituto de Antropología, 5, 17-24.
Cocilovo, J. A., Varela, H. H., & O’Brien, T. G. (2009). La divergencia
genética entre poblaciones del Área Andina Centro Meridional
evaluada mediante rasgos no métricos del cráneo. Revista Argentina
de Antropologia Biologica, 11, 43-59.
Cocilovo, J. A., Zavattieri, M. V., & Costa Junqueira, M. A. (1994).
Biología del grupo prehistórico de Coyo Oriental (San Pedro de
Atacama, Norte de Chile): I Dimorfismo sexual y variación etaria.
Estudios Atacameños, 11, 121-134.
Cocilovo, J. A., Varela, H. H., Costa-Junqueira, M. A., & Quevedo, S.
G. (2005). Los pescadores arcaicos de la desembocadura del río Loa
(Norte de Chile): El sitio Caleta Huelén 42. Chungara, 37, 5-19.
Cocilovo, J. A., Rothhammer, F., Quevedo, S., & Llop, E. (1982).
Microevolución en poblaciones prehistóricas del área andina. 3. La
población del Morro de Arica. Craneometría. Revista de la Uni-
versidad Nacional de Río Cuarto, 2 , 91-111.
Cocilovo, J. A., Quevedo, S., Varela, H. H., Valdano, S., & Castro, M.
(1999). Biología del grupo prehistórico de Pisagua, costa norte de
Chile. Estudios Atacameños, 1 7, 207-235.
Dembo, A., & Imbelloni, J. (1938). Deformaciones intencionales del
cuerpo humano. Buenos Aires: Humanior.
Dingwall, E. J. (1931). Artificial cranial deformation—A contribution
to the study of ethnic mutilation. London: John Bale, Sons & Dan-
ielsson, Ltd.
Dettwyler, K. A. (1992). Nutritional status of adults in rural mali.
American Journal of Physical Anthropology, 88, 309-321.
doi:10.1002/ajpa.1330880306
Frayer, D. W., & Wolpoff, M. H. (1985). Sexual dimorphism. Annual
Review of Anthropology, 14, 429-473.
doi:10.1146/annurev.an.14.100185.002241
Giles, E., & Elliot, O. (1963). Sex determination by discriminant func-
tion analysis of crania. American Journal of Physical Anthropology,
21, 53-68. doi:10.1002/ajpa.1330210108
Guntupalli, A. M., & Baten, J. (2009). Measuring gender well-being
with biological living-standard indicators. In B. Harris, L. Gálvez, &
H. Machado (Eds.), Gender and well-being in Europe: Historical
and contemporary perspectives (pp. 43-58). London: Ashgate Ed.
Gustafsson, A., & Lindenfors, P. (2004). Human size evolution: No
evolutionary allometric relationship between male and female stature.
Journal of Human Evolution, 4 7, 253-266.
doi:10.1016/j.jhevol.2004.07.004
Gustafsson, A., & Lindenfors, P. (2009). Latitudinal patterns in human
stature and sexual stature dimorphism. Annals of Human Biology, 36,
74-87. doi:10.1080/03014460802570576
Gustafsson, A., Werdelin, L., Tullberg, B. S., & Lindenfors, P. (2007).
Stature and sexual stature dimorphism in Sweden, from the 10th to
the end of the 20th Century. American Journal of Human Biology, 19,
861-870. doi:10.1002/ajhb.20657
Keen, J. A. (1950). A study of the differences between male and female
skulls. American Journal of Physical Anthropology, 8, 65-78.
doi:10.1002/ajpa.1330080113
Lovejoy, C. O. (1985). Dental wear in the Libben population: Its func-
tional pattern and role in the determination of adult skeletal age and
death. American Journal of P hys ica l Anthropology, 68, 47-56.
doi:10.1002/ajpa.1330680105
Masset, C. (1989). Age Estimation on the basis of cranial sutures. In M.
Y. Iscan (Eds.), Age markers in the human skeleton (pp. 71-103).
Springfield: Charles C Thomas.
Meindl, R. S., & Lovejoy, C. O. (1985). Ectocranial suture closure: A
revised method for the determination of skeletal age at death based
on the lateral-anterior sutures. American Journal of Physical An-
thropology, 68, 57-66. doi:10.1002/ajpa.1330680106
Méndez, M. G., & Ferrarini, S. O. (2006). Dimorfismo sexual y
cefalometría en etnias chaqueñas: Una lectura desde la antropología
evolutiva. Revista Española de Antropología Física, 2 6, 75-92.
Mendonça, O. J., Valdano, S. G., & Cocilovo, J. A. (1994). Evaluación
del Dimorfismo Sexual y de la Deformación Artificial en una
muestra craneana del borde oriental de la Puna Jujeña. Revista
Argentina de Antropologia Biologica, 2, 25-37.
O’Brien, T. G. (2003). Cranial microvariation in prehistoric south cen-
tral andean populations: An assessment of morphology in the
Cochabamba Collection, Bolivia. Ph.D. Thesis, Binghamton: Bing-
hamton University, State University of New York.
Pinotti, L. V., Rasines, C. H., & Unsain, R. F. (2005). Tehuelches y
mapuches, dimorfismo sexual en condiciones de vulnerabilidad
social. Studies in Historical Anthropology, 2, 17-29.
Priotto, J. M. (1990). Diferenciación biológica y dimorfismo sexual
entre las poblaciones aborígenes del Chaco Occidental (Chiriguanos,
Chorotes, Matacos y Tobas). Degree thesis, Río Cuarto: Universidad
Nacional de Río Cuarto.
Pucciarelli, H. M., Carnese, F. R., Pinotti, L. V., Guimarey, L. M., &
Goicoechea, A. S. (1993). Sexual dimorphism in schoolchildrem of
the Villa IAPI neighborhood (Quilmes, Buenos Aires, Argentina).
American Journal of Physical Anthropology, 92, 165-172.
doi:10.1002/ajpa.1330920206
Purkait, R., & Chandra, H. (2002). Sexual dimorphism in femora. In-
dian Study Forensic Science Communications Research and Tech-
nology, 4. URL (last checked 7 November 2012).
http://www2.fbi.gov/hq/lab/fsc/backissu/july2002/purkait.htm
Purkait, R., & Chandra, H. (2004). A study of sexual variation in Indian
femur. Forensic Science Inter na t i on a l, 146, 25-33.
doi:10.1016/j.forsciint.2004.04.002
Saunders, S. R. (1992). Subaldult skeletons and growth related studies.
In S. R. Saunders, & A. Katzenberg (Eds.), Skeletal biology of past
peoples: Research method (pp. 1-20). New York: Wiley-Liss.
Seber, G. A. F. (1984). Multivariate observations. New York: Wiley.
doi:10.1002/9780470316641
Stini, W. A. (1969). Nutritional stress and growth: Sex difference in
adaptive response. American Journal of Physical Anthropology, 31,
417-426. doi:10.1002/ajpa.1330310316
Stinson, S. (1985). Sex differences in environmental sensitivity during
growth and development. Yearbook of Physical Anthropology, 28,
123-147. doi:10.1002/ajpa.1330280507
Varela, H. H., Priotto, J. W., & Cocilovo, J. A. (1990a) Evaluación del
dimorfismo sexual en una muestra de nativos de la Puna Jujeña.
Revista de la Universidad Nacional de Río Cuarto, 10, 33-42.
Varela, H. H., Cocilovo, J. A., & Costa Junqueira, M. A. (1990b). El
dimorfismo sexual en la población prehistórica de San Pedro de
Atacama, Chile. Chungara, 24-25, 159-166.