Paper Menu >>
Journal Menu >>
 American Journal of Plant Sciences, 2010, 1, 17-23 doi:10.4236/ajps.2010.11003 Published Online September 2010 (http://www.SciRP.org/journal/ajps) Copyright © 2010 SciRes. AJPS 17 Genetic Analysis of Leucine Content in Indica-Japonica Hybrid Rice (Oryza sativa L.) Xiaoming Zhang1,2, Chunhai Shi2*, Jianguo Wu 2, Shenghai Ye1, Genliang Bao1, Wenchao Yan1 1Institute of Crop Research and Atomic Energy Utilization, Zhejiang Academy of Agricultural Sciences, Hangzhou, People’s Repub- lic of China; 2College of Agriculture and Biotechnology, Zhejiang University, Hangzhou, People’s Republic of China. Email: chhshi@zju.edu.cn Received August 10th, 2010; revised September 6th, 2010; accepted September 13th, 2010 ABSTRACT Genetic control of leucine content in indica-japonica hybrid rice (Oryza sativa L.) was studied in 35 crosses of F1 and F2 generations, which were derived from crossing 7 male sterile indica rice lines with 5 restorer japonica rice lines along with their parents. Two g enetic models and their corresponding statistical methods for qua ntitative traits of trip- loid seeds in cereal crops were u sed for the a nalysis. The first was th e unconditional g enetic model, which refers to the analysis of cumulative measurements (from flowering to a specific time) along the develo pmental stages, while the sec- ond was the conditional genetic model, which relates to analysis from one developmental stage to another stage (t - 1→ t). The results showed that leucine content of indica-japonica hybrid rice was controlled by the expression of triploid endosperm effect (endosperm additive effect and endosperm dominant effect), cytoplasm effect, diploid maternal plant effect (maternal additive effect and maternal dominant effect) and their environmental interaction effects. Of these ef- fects, endosperm dominant effect and maternal dominant effect were more important at the earlier stages, while en- dosperm additive effect and maternal additive effect were more important at the later stages of rice grain development under both unco nditional and co nditiona l genetic analyses. D ue to the h igh heritabilities, which came from endosperm , maternal and cytoplasm effects for leucine content at different developmental stages, selection for leucine content of indica-japonica hybrid rice would be more efficient at early generations in breeding programs. Keywords: Indica-Japonica Hybrid Rice, Developmental Genetics, Quality, Leucine Content, Genetic Variance, Conditional Genetic Variance, Heritability 1. Introduction The use of indica-japonica F1 hybrids offers a way of co- mbining the best attributes of both types. Advantages of these hybrids include strong tillering ability, thick culms and larger panicles. However, some disadvantages have also been noted, such as longer growing duration, greater plant height and reduced spikelet fertility. Recently, sign- ificant progress has been made in improving agronomic traits in an indica-japonica hybrid breeding project [1] and breeders are now paying greater attention to improv- ing rice grain quality traits. In most reports, phenotypic values measured at ma- turity are used when analyzing rice quality traits [2-7], but this does not provide information on the develop- mental processes of these traits. Although recent quan- titative trait analysis software makes developmental genetic analysis possible [8], the literatures are limited to indica rice apparent quality and milling quality [9-11]. A little information on nutrition of indica-japonica hy- brid rice has been reported, especially for amino acids. One reason for the lack of information is the difficu lty in obtaining a significant quantity of F1 seeds for analysis due to incompatibility between the indica and japonica rice subspecies when crossed. Some japonica wide compatible restorers developed by our projects make this genetic analysis in indica-japonica hybrid rice pos- sible. Leucine (C6H13NO2) is a neutral, genetically coded essential amino acid in human nutrition, which is very important in human health. The expression of genes for leucine content at different grain-filling stages is yet to be understood. Understanding the dynamics of gene ex- pression for leucine content in contrasting environments may prove helpful for future improvements in the nutri- tional quality of rice. The objective of this study was to clarify the developmental behavior of leucine content *corres p ondin g autho r  Genetic Analysis of Leucine Content in Indica-Japonica Hybrid Rice (Oryza sativa L.) Copyright © 2010 SciRes. AJPS 18 gene expression in indica-japonica hybrid rice. The de- vel op men ta l dynamic expression mechanism an d th e h er i- tability components from different genetic systems were estimated by unconditional and conditional genetic models . 2. Materials and Methods 2.1. Plant Material Seven indica cytoplasmic male sterile lines (A lines) (Zhe 38, Xieqingzao, K17, Zhenshan 97, Zhenong 8010, Jin 23 and Ⅱ-32) and their maintainers (B lines), and 5 jap- onica wide compatible restorers (R lines) (T 748, T 42, R 1252, Linhui 422 and Zhong 9308) with significant var i- ation in leucine contents were used in this experiment (Table 1). 2.2. Field Experiment F1 seeds were obtained by crossing female A lines to male R lines (A R) using a factorial design. The F1s and their parents were sown at the experimental farm, Zhejiang Academy of Agricultural Sciences, China. After 25 days, seedlings were individually transplanted at a spacing of 20 26 cm. There were 36 plants in each plot and two replications. A second exper iment was conducted 7 mon- ths later using the same methods to provide contrasting environmental conditions at Hainan province, China. Se- eds from parental lines and F2 seeds from F1 plants were collected at 7-, 14-, 21-, 28- and 35-day after flowering from the central 16 plants within each plot. The F1 plants resulted from A × R crosses. The grain-filling period was devided into stages, the initial stage (1-7days after flow- ering), early stage (8-14days), middle stage (15-21days), late stage (22-28 days) and ripe stage (29-35days). 2.3. Leucine Content Analysis All seed hulls of F1s, F2s and their parents were removed using a Yanmar ST50 dehuller from Japan, then milled using a sample miller (model JB-20, Zhejiang province, China). Samples were further ground to 100 mesh with a cyclone grinder (model 3010-019, Fort Collin s, Colorado, USA). Leucine content was determined using the amino acid analysis method described by Wu et al. [12] with two replications for each sample. 2.4. Statistical Method Two developmental genetic models, the unconditional genetic model [13] and the conditional genetic model [8], for quantitative traits of endosperm in cereal crops were used to estimate the variance components for ge- netic main effects and genotype- environmental interac- tion effects at different grain-filling stages. For the un- conditional genetic analysis, which refers to the analy- sis of cumulative measurements through progressive developmental stages, the genetic effects were defined as accumulated effects of genes expressed from flow- ering (0) to a particular time (t). The variance compo- nents were divided into endosperm additive variance (VA), endosperm dominance variance (VD), cytoplasmic variance (VC), maternal additive variance (VAm), maternal Table 1. Leucine content (%) of 12 parents at five different developmental stages. Parent Initial stage Early stage Middle stage Late stage Ripening stage Female group Zhe38 1.113a 1.122a 1.274a 1.203ab 1.083ab Xieqingzao 0.769b 0.805d 0.921cd 0.926bc 0.781e K17 0.788b 0.751d 0.906cd 0.934bc 0.867cde Zhenshan97 0.984ab 0.966bc 1.097abc 1.198ab 0.951bcd Zhenong8010 0.738b 0.872cd 0.849d 1.021abc 0.743e Jin23 1.008ab 1.019ab 1.186ab 1.271a 1.120a II-32 0.723b 0.757d 0.860cd 0.991abc 0.891cde Standard deviation 0.156 0.142 0.171 0.143 0.142 Male group T748 0.748b 0.843cd 0.838d 0.875bc 0.860cde T42 1.029ab 1.041ab 1.070abcd 1.080abc 0.959bc R1252 0.852ab 0.829d 0.947bcd 1.024abc 0.824cde Linhui422 0.871ab 0.820d 0.935cd 0.940bc 0.807de Zhong9308 0.734b 0.771d 0.838d 0.863c 0.957bcd Standard deviation 0.119 0.104 0.096 0.094 0.073 a,b,c,d,e denote significance at 0.05 probability level.  Genetic Analysis of Leucine Content in Indica-Japonica Hybrid Rice (Oryza sativa L.) Copyright © 2010 SciRes. AJPS 19 dominance variance (VDm), and their endosperm additive environment interaction variance (VAE), endosperm do- minance interaction variance (VDE), cytoplasm interaction variance (VCE), maternal additive interaction variance (VAmE) and maternal dominance interaction variance (VDmE), and residual variance (Ve). Since some endosperm genes were derived from maternal plants, there were possible covari- ances between endosperm effects and maternal effects in- cluding additive covarian ce CA.Am, dominance covariance CD.Dm, additive interaction covariance CAE.AmE, and dom- inance interaction covariance CDE.DmE. The partitioning for the phenotypic variance (VP(t)) was: 2() 2() P A D CAmDmAEDECEAmEDmE A AmD DmAEAmEDE DmEe VVVVV V V V VVV CCC CV For the condition al genetic analysis, wh ich refers to gr- ain analysis within each stage, th e conditional developm- ental genetic models could be used to estimate condition- al variances during rice grain filling p eriods (t – 1→t) for leucine content. These conditional variance components were VA(t | t - 1) (conditional endosperm additive variance), VD(t | t - 1) (conditional endosperm dominance variance), VC(t | t - 1) (conditional cytoplasmic variance), VAm(t | t - 1) (conditional maternal additive variance), VDm(t | t - 1) (con- ditional maternal dominance variance), VAE(t | t - 1) (cond- itional endosperm additive interaction variance), VDE(t | t - 1) (conditional endosperm dominance interaction variance), VCE(t | t - 1) (conditional cytoplasm interaction variance), VAmE(t | t - 1) (conditional maternal additive interaction variance), VDmE(t | t - 1) (conditional maternal dominance interaction variance), CA.Am(t | t - 1) (conditional additive covariance), CD.Dm(t | t - 1) (conditional dominance cova- riance), CAE.AmE(t | t - 1) (conditional additive interaction covariance), CDE.DmE(t | t - 1) (conditional dominance in- teraction covariance) and Ve(t | t - 1) (conditional residual variance). The partitio ning fo r the cond ition al pheno typic variance (VP(t | t - 1)) was: )1()1()1( )1()1()1( )1()1()1()1( )1()1()1()1()1()1( )(2 )(2 ttettDmEDEttAmEAE ttDmDttAmAttDmE ttAmEttCEttDEttAE ttDmttAmttCttDttAttP VCC CCV VVVV VVVVVV Heritabilities (2 h) can be further divided into general heritability (2 G h) and environment interaction heritability (2 GE h) by unconditional genetic analysis. The general heritability (2 G h), which was controlled by genetic main effects, could be further divided into endosperm general heritability (PAmAAGo VCVh )( 2 ), cytoplasm heritabil- ity (PCGc VVh 2) and maternal general heritability (2 Gm h PAmAAm VCV )( ). The interaction heritability (2 GE h), which was controlled by genotype environmental inter- action effects, also could further be divided into endosp- erm environment interaction heritability (AEGoE Vh ( 2 PAmEAE VC/) ), cytoplasmic environment interaction heritability (PCEGcEVVh 2) and maternal environment interaction heritability (PAmEAEAmEGmEVCVh )( 2 ). The partitioning for the total narrow-sense heritability was: )()( 222222222GmEGcEGoEGmGcGoGEG hhhhhhhhh The Jackknife re-sampling method was used by sam- pling generation means of entries for estimatin g standard errors of variances components, covariance and herita- bilities [14-15]. 3. Results 3.1. Difference of Leucine Content in Parents and their Descendants There was significant variation for leucine content both in female parents and in male parents at five different fi- lling stages (Table 1). Of the A lines, Zhe38 and Jin23 had high leucine content at the initial stage, while of the B lines, T42 had the highest leucine content at the initial stage. Zhenong 8010 and T748 were developed quickly than others at early filling stage, Xieqingzao, Zhen- shan97, R1252 and Linhui422 were developed faster at middle filling stage. All the parents except Zhong 9318 reached their highest level of leucine content at the late grain-filling stage, and the results showed that Jin 23 was the highest (1.271%) and K17 was the lowest (0.934%). At the ripe stage, leucine content of Jin 23 was signifi- cant high than that of II-32, K17, T748, R1252, Linhui 422, Xieqingzao and Zhenong 8010 (Table 1). The mean leucine contents of indica-japonica hybrids of females, males and their F1 and F2 descendants are sh- own in Figure 1. There were continuous increases after 0.6 0.7 0.8 0.9 1.0 1.1 LC % 7d 14d 21d 28d35d Female Male F 1 F 2 Figure 1. Leucine contents of 4 generations at five different filling stages. 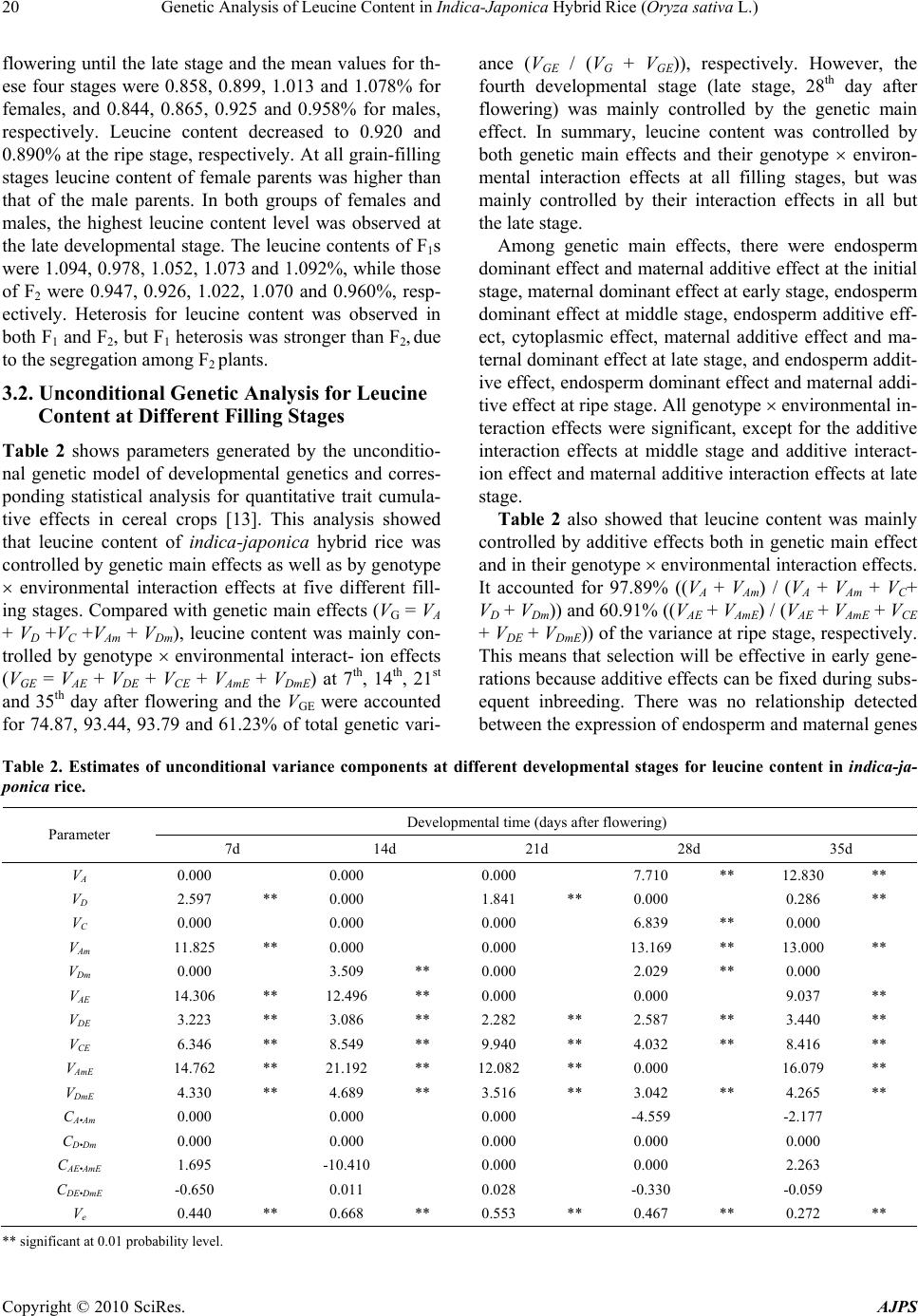 Genetic Analysis of Leucine Content in Indica-Japonica Hybrid Rice (Oryza sativa L.) Copyright © 2010 SciRes. AJPS 20 flowering until the late stage and the mean values for th- ese four stages were 0.858, 0.899, 1.013 and 1.078% for females, and 0.844, 0.865, 0.925 and 0.958% for males, respectively. Leucine content decreased to 0.920 and 0.890% at the ripe stage, respectively. At all grain-filling stages leucine content of female parents was higher than that of the male parents. In both groups of females and males, the highest leucine content level was observed at the late developmental stage. The leucine conten ts of F1s were 1.094, 0.978, 1.052, 1.073 and 1.092%, while those of F2 were 0.947, 0.926, 1.022, 1.070 and 0.960%, resp- ectively. Heterosis for leucine content was observed in both F1 and F2, but F1 heterosis was strong er than F2, due to the segregation among F2 plants. 3.2. Unconditional Genetic Analysis for Leucine Content at Different Filling Stages Table 2 shows parameters generated by the unconditio- nal genetic model of developmental genetics and corres- ponding statistical analysis for quantitative trait cumula- tive effects in cereal crops [13]. This analysis showed that leucine content of indica-japonica hybrid rice was controlled by genetic main effects as well as by genotype environmental interaction effects at five different fill- ing stages. Compared with genetic main effects (VG = VA + VD +VC +VAm + VDm), leucine content was mainly con- trolled by genotype environmental interact- ion effects (VGE = VAE + VDE + VCE + VAmE + VDmE) at 7th, 14th, 21st and 35th day after flowering and the VGE were accounted for 74.87, 93.44, 93.79 and 61.23% of total genetic vari- ance (VGE / (VG + VGE)), respectively. However, the fourth developmental stage (late stage, 28th day after flowering) was mainly controlled by the genetic main effect. In summary, leucine content was controlled by both genetic main effects and their genotype environ- mental interaction effects at all filling stages, but was mainly controlled by their interaction effects in all but the late stage. Among genetic main effects, there were endosperm dominant effect and maternal additive effect at the initial stage, maternal dominant effect at early stage, endosperm dominant effect at middle stage, endosperm additive eff- ect, cytoplasmic effect, maternal additive effect and ma- ternal dominant effect at late stage, and endosperm addit- ive effect, endosperm dominant effect and maternal addi- tive effect at ripe stage. All genotype environmental in- teraction effects were significant, except for the additive interaction effects at middle stage and additive interact- ion effect and maternal additive interaction effects at late stage. Table 2 also showed that leucine content was mainly controlled by additive effects both in genetic main effect and in their genotype environmental interaction effects. It accounted for 97.89% ((VA + VAm) / (VA + VAm + VC+ VD + VDm)) and 60.91% ((VAE + VAmE) / (VAE + VAmE + VCE + VDE + VDmE)) of the varia nce at ripe stage, respectively. This means that selection will be effective in early gene- rations because additive effects can be fixed during subs- equent inbreeding. There was no relationship detected between the expression of endosperm and maternal genes Table 2. Estimates of unconditional variance components at different developmental stages for leucine content in indica-ja- ponica rice. Developmental time (days after flowering) Parameter 7d 14d 21d 28d 35d VA 0.000 0.000 0.000 7.710 ** 12.830 ** VD 2.597 ** 0.000 1.841 ** 0.000 0.286 ** VC 0.000 0.000 0.000 6.839 ** 0.000 VAm 11.825 ** 0.000 0.000 13.169 ** 13.000 ** VDm 0.000 3.509 ** 0.000 2.029 ** 0.000 VAE 14.306 ** 12.496 ** 0.000 0.000 9.037 ** VDE 3.223 ** 3.086 ** 2.282 ** 2.587 ** 3.440 ** VCE 6.346 ** 8.549 ** 9.940 ** 4.032 ** 8.416 ** VAmE 14.762 ** 21.192 ** 12.082 ** 0.000 16.079 ** VDmE 4.330 ** 4.689 ** 3.516 ** 3.042 ** 4.265 ** CA•Am 0.000 0.000 0.000 -4.559 -2.177 CD•Dm 0.000 0.000 0.000 0.000 0.000 CAE•AmE 1.695 -10.410 0.000 0.000 2.263 CDE•DmE -0.650 0.011 0.028 -0.330 -0.059 Ve 0.440 ** 0.668 ** 0.553 ** 0.467 ** 0.272 ** ** significant at 0.01 probability level.  Genetic Analysis of Leucine Content in Indica-Japonica Hybrid Rice (Oryza sativa L.) Copyright © 2010 SciRes. AJPS 21 since the genetic main covariances (CA.Am or CD.Dm) and ge- notype environmental interaction covariances (CAE•AmE or CDE•DmE) between the endosperm and maternal effects were not significant. Leucine content may be influenced by sampling errors according to the significant residual variances (Ve). But, in comparison with other genetic parameters, the values of Ve were lower. 3.3. Conditional Genetic Analysis for Leucine Content at Different Filling Stages The genetic variances estimated by unconditional genetic analysis revealed accumulated genetic effects which we- re expressed from flowering to time t (0→t), and the res- ults could not clarify gene expression at each special dev- elopmental stage (t – 1→t). Therefore, conditional gene- tic analysis was used to clearly explain the dynamic gene expression at each filling stage (t – 1→t). Table 3 shows the parameters generated by the condi- tional genetic model of developmental genetics and cor- responding statistical analysis for quantitative trait p erio- dical effects of triploid in cereal crops [8]. There was no detection of new gene expression for conditional en- dosperm additive main effects (VA (7|0), VA (14|7) and VA (21|14)) from initial- to middle-filling stages, conditional maternal additive main effects (VAm(14|7) and VAm(21|14)) at early- and middle-filling stages, conditional endosperm dominant main effects (VD(21|14) and VD(28|21)) at middle- and late-stages, conditional maternal dominant main eff- ects (VDm(7|0) and VDm(28|21)) at initial- and late-filling st- ages and conditional cytoplasmic main effects (VDm(7|0) and VC(35|28)) at initial- and ripe stages, since the condi- tional genetic main variances at these filling stages were not significant in this experiment. The unconditional ma- ternal dominant main effect at late filling stage in Table 2 might be due to the dominant effect expressed at the former stage, i.e. at middle stage (14-21 days after flow- ering), and then continual expression from activated genes at this stage. Similarly, the endosperm dominant main effect was significant at 21 days after flowering by unconditional genetic analysis, but was not significant us- ing conditional genetic analysis. In contrast, conditional maternal dominant effects at the ripe stage were found by conditional genetic analysis, but not found by uncondit- ional genetic analysis in Table 2. This may reflect inter- ruption of gene expression during different filling stages or the gradual activation of quantitative genes through the filling period(s). It may also indicate that there were differences in regulation of gene expression in triploid endosperm, cytoplasm and diploid maternal plant genetic systems. These results were not detected by the uncondi- tional genetic variance analysis. For the conditional interaction analysis, new genetic effects from gene expression was not detected for the con- Table 3. Estimates of conditional variance components for leucine content at different developmental stages in indica-jap- onica rice. Developmental stage of grain (t|t-1) Parameter 7d|0d 14d|7d 21d|14d 28d|21d 35d|28d VA (t|t-1) 0.000 0.000 0.000 4.324 ** 9.703 ** VD (t|t-1) 2.597 ** 4.700 ** 0.000 0.000 1.279 ** VC (t|t-1) 0.000 7.137 ** 3.621 ** 2.848 ** 0.000 VAm (t|t-1) 11.825 ** 0.000 0.000 9.158 ** 6.559 ** VDm (t|t-1) 0.000 3.277 ** 6.975 ** 0.000 1.076 ** VAE (t|t-1) 14.306 ** 13.463 ** 10.171 ** 0.000 8.072 ** VDE (t|t-1) 3.223 ** 0.000 4.543 ** 2.809 ** 2.503 ** VCE (t|t-1) 6.346 ** 0.000 0.000 0.000 0.000 VAmE (t|t-1) 14.762 ** 7.358 ** 3.852 ** 0.000 11.931 ** VDmE (t|t-1) 4.330 ** 6.009 ** 0.000 2.290 ** 3.941 ** CA•Am (t|t-1) 0.000 0.000 0.000 -2.819 -8.654 CD•Dm (t|t-1) 0.000 -0.080 0.000 0.000 3.889 ** CAE•AmE (t|t-1) 1.695 -46.510 -2.301 0.000 2.404 CDE•Dm E(t|t-1) -0.650 0.000 0.000 -0.436 -0.290 Ve(t|t-1) 0.440 ** 0.738 ** 0.794 ** 0.413 ** 0.257 ** ** was signif icant at 0.01 probability level.  Genetic Analysis of Leucine Content in Indica-Japonica Hybrid Rice (Oryza sativa L.) Copyright © 2010 SciRes. AJPS 22 ditional endosperm additive interaction effect (VAE (28|21)), conditional maternal additive interaction effect (VAmE (28|21)) at late filling stage, conditional endosperm domin- ant interaction effect (VDE(14|7)) at early stage, conditional maternal dominant interaction effect (VDmE (21|14)) at middle stage and conditional cytoplasm interac- tion effect (VCE (14|7), VCE (21|14), VCE (28|21) and VCE (35|28)) at early-, middle- and ripe-stage. The VDE at early stage, VDmE at middle stage and VCE at early-, middle- and ripe-stage were significant using the unconditional ge- netic analysis in Table 2, but were not significant using the conditional genetic analysis in Table 3. It is likely that there was continual expression of activated genes at these stages. There was significant positive conditional dominant interaction covariance (CD•Dm(35\28)=3.889**) at ripe stage, which showed that the new expression for dominance effect from endosperm nuclear genes was closely related with that from maternal nuclear genes at the ripe stage. Significant conditional residual variances (Ve(t|t-1)) sho- wed that new expression of genes for leucine content at the ripe stage could be influenced by sampling errors, but these values were lower than the genetic variances. 3.4. Estimation of Heritability at Different Filling Stages The results in Table 4 showed that general heritability components from cytoplasm and endosperm were not si- gnificant at first three stages, but were significant at the next two developmental stages for endosperm and the fo- urth stage for cytoplasm, while their interaction heritabil- ity components were all significant except for endosperm interaction heritabilities at middle- and late-stage. Ma- ternal general heritabilities were significant except for early- and middle-stage and its interaction heritabilities except for late filling stage. The interaction heritabilities including endosperm, maternal and cytoplasm interaction heritabilities were larger than their general heritability components at the first three stages and the last filling st- age, i.e. initial-, early-, middle- and ripe stage, while general heritability was larger than the in teraction herita- bility at late filling stage. The total narrow sense herita- bility values (2 h) at most filling stages for leucine con- tent were over 75% (h(7d)=84.50, h(14d)= 64.14, h(21d)= 72.75, h(28d)= 75.20 and h (35d)= 87.97%). The general heritabilities were 19.70, 0.00, 0.00, 61.80 and 31.73% and interaction heritabilities were 64.80, 64.14, 72.75, 13.40 and 56.23%, respectively. With regard to the components of heritability, maternal and cytoplasm gen- eral heritabilities and interaction heritabilities were more important for leucine content (2222 GcEGmEGcGm hhhh = 57.80, 57.89, 72.75, 75.20 and 55.53%, respectively) in this experiment. These data suggest that improving leu- cine content would be more efficient when selection is based on maternal plants in early indica-japonica gen- erations. 4. Discussion The genetic behav ior of rice q uantitative traits is simulta- neously controlled by endosperm, maternal and cytopla- sm effects. Genes controlling the performance of comp- lex quantitative traits, were expressed at various times during different developmental stages [8-10]. Analysis for quantitative traits has been gradually emphasized in developmental genetics. This research revealed that gene expression varied du- ring most filling stages in endosperm, cytoplasm and ma- ternal genetic systems across environments. Leucine con- tent was mainly controlled by additive effects, maternal effects and cytoplasmic effects, both by variance analysis and by heritabilities analysis. This means that selection for leucine content would be more efficient at early gen- erations in a rice breeding program. The results of condi- tional genetic variance analysis for leucine content fur- ther indicated that there was a phenomenon of interval expression for some genes among rice filling stages. For example, the conditional genetic analysis, found a sig- nificant maternal additive effect at the initial stage, which was non-significant at the early- and middle- stage, but Table 4. Estimates of heritability components (%) for leucine content at different developmental stages in indica-japonica rice. Heritability Parameter 7d 14d 21d 28d 35d 2 Go h 0.000 0.000 0.000 10.470 15.740 2 Gc h0.000 0.000 0.000 22.724 ** 0.000 2 Gm h19.700 ** 0.000 0.000 28.608 ** 15.992 + 2 GoE h 26.700 6.249 0.000 0.000 16.697 * 2 GcE h10.600 25.603 ** 32.837 ** 13.397 ** 12.435 * 2 GmE h27.500 32.289 ** 39.914 ** 0.000 27.101 ** +, * and ** denote signific ance at 0. 10 , 0.05 and 0.01 probability levels, respectively.  Genetic Analysis of Leucine Content in Indica-Japonica Hybrid Rice (Oryza sativa L.) Copyright © 2010 SciRes. AJPS 23 was again significant at the late- and ripe- stage. Genotype environment interaction effects including endosperm additive and dominant interaction effects, ma- ternal additive and dominant interaction effect, and cyto- plasmic interaction effects were found at most filling st- ages by both unconditional genetic analysis and condi- tional genetic analysis but cytoplasmic interaction effect in this study. Significant genotype environment intera- ction effects also indicated that the sequential expression of genes from endosperm, cytoplasm and maternal gen- etic systems was also influenced by the environmental conditions. The geno type environment interaction effe- ct was the main cause of genetic differences across envi- ronments. It is necessary to consider the variation of rice quantitative traits in different environments because of the varied climatic conditions to which rice is exp osed. The genetic models and statistical analysis methods used in this experiment, which include genetic main eff- ects and genotype environment interaction effects, can be analyzed with only three generations, such as parents, F1 and F2 at different filling stages across a range of en- vironments. Evidence of varying gene expression for leu- cine content during grain filling in rice serves to under- score the need to study developmental behavior of gene expression for important quantitative traits in o ther cereal crops. 5. Acknowledgements The project was supported by the Science and Technol- ogy Office of Zhejia ng P ro vi nce, C hina (N o . 20 08C 1 40 71 and 011102471), Zhejiang Provincial Natural Science Foundation of China, 151 Foundations for the Talents of Zhejiang Province and the Exchange Program of Friend- ship between Zhejiang Government of China and Fukui of Japan. REFERENCES [1] W. Q. Dong, S. H. Shi and Y. J. Dong, “Breeding and Utility of a New Hybrid Rice Xieyou9516,” Zhejiang Ag- ricultural Science, Vol. 5, No. 5, 1999, pp. 211-213. [2] J. S. Chauhan, V. S. Chauhan and S. B. Lodh, “Environ- ment Influence on Genetic Parameters of Quality Com- ponent in Rainfed Upland Rice,” Indian Journal of Agri- cultural Science, Vol. 62, No. 5, 1992, pp. 773-775. [3] K. F. Osato, Y. Hamachi and Y. Matsue, “Genotype Environment Interaction of Palatability in Rice,” Japa- nese Journal of Crop Science, Vol. 65, No. 5, 1996, pp. 585-589. [4] C. H. Shi, J. M. Xue, Y. G. Yu, X. E. Yang and J. Zhu, “Analysis of Genetic Effects for Nutrient Quality Traits in Indica Rice,” Theoretical and Applied Genetics, Vol. 92, No. 8, 1996, pp. 1099-1102. [5] C. H. Shi, J. Zhu, X. E. Yang, Y. G. Yu and J. G. Wu, “Genetic Analysis for Protein Content in Indica Rice,” Euphytica, Vol. 107, No. 2, 1999, pp. 135-140. [6] Y. F. Tan, J. X. Li and C. G. Xu, “Genetic Bases of Ap- pearance Quality of Rice Grains in Shanyou 63, an Elite Rice Hybrid,” Theoretical and Applied Genetics, Vol. 101, No. 5-6, 2000, pp. 823-829. [7] C. W. Xu, A. H. Zhang and Q. S. Zhu, “Genetic Analysis of Quality Traits in Rice Crosses between Indica and Ja- ponica,” Acta Agronomica Sinica, Vol. 22, No. 5, 1996, pp. 530-534. [8] J. Zhu, “Analysis of Conditional Genetic Effects and Variance Components in Developmental Genetics,” Ge- netics, Vol. 141, No. 4, 1995, pp. 1633-1639. [9] C. H. Shi, J. G. Wu and P. Wu, “A Developmental Be- havior of Gene Expression for Brown Rice Thickness under Different Environments,” Genesis, Vol. 33, No. 4, 2002, pp. 185-190. [10] C. H. Shi, J. G. Wu, X. B. Lou, J. Zhu and P. Wu, “Ge- netic Analysis of Transparency and Chalkiness Area at Different Filling Stages of Rice (Oryza sativa L.),” Field Crops Research, Vol. 76, No. 1, 2002, pp. 1-9. [11] C. H. Shi, J. G. Wu, X. M. Zhang and P. Wu, “Develop- mental Analysis on Genetic Behavior of Brown Rice Re- covery in Indica Rice across Environments,” Plant Sci- ence, Vol. 163, No. 3, 2002, pp. 555-561. [12] J. G. Wu, C. H. Shi and X. M Zhang, “Estimating the Amino Acid Composition in the Milled Rice Powder by Near-Infrared Reflectance Spectroscopy,” Field Crops Research, Vol. 75, No. 1, 2002, pp. 1-7. [13] J. Zhu and B. S. Weir, “Analysis of Cytoplasm and Ma- ternal Effects: II. Genetic Models for Triploid Endospe- Rms,” Theoretical and Applied Genetics, Vol. 89, No. 2-3, 1994, pp. 160-166. [14] R. G. Miller, “The Jackknife- A review,” Biometrika, Vol. 61, No. 1, 1974, pp. 1-15. [15] J. Zhu and B. S. Weir, “Diallel analysis for sex-linked and maternal effects,” Theoretical and Applied Genetics, Vol. 92, No. 1, 1996, pp. 1-9. |