S. Yakoubou et al. / Natural Science 2 (2010) 990-997
Copyright © 2010 SciRes. OPEN ACCESS
997
997
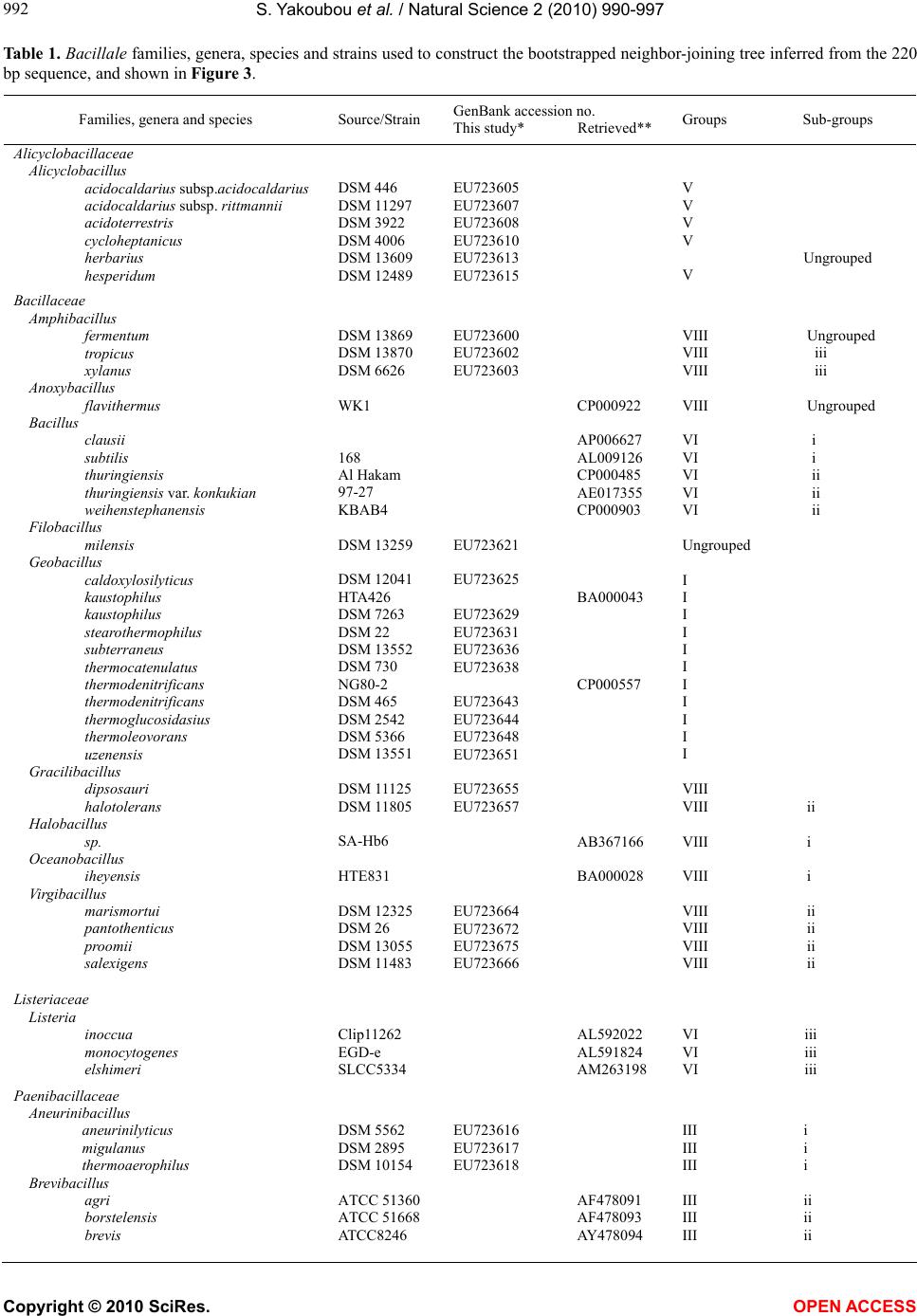
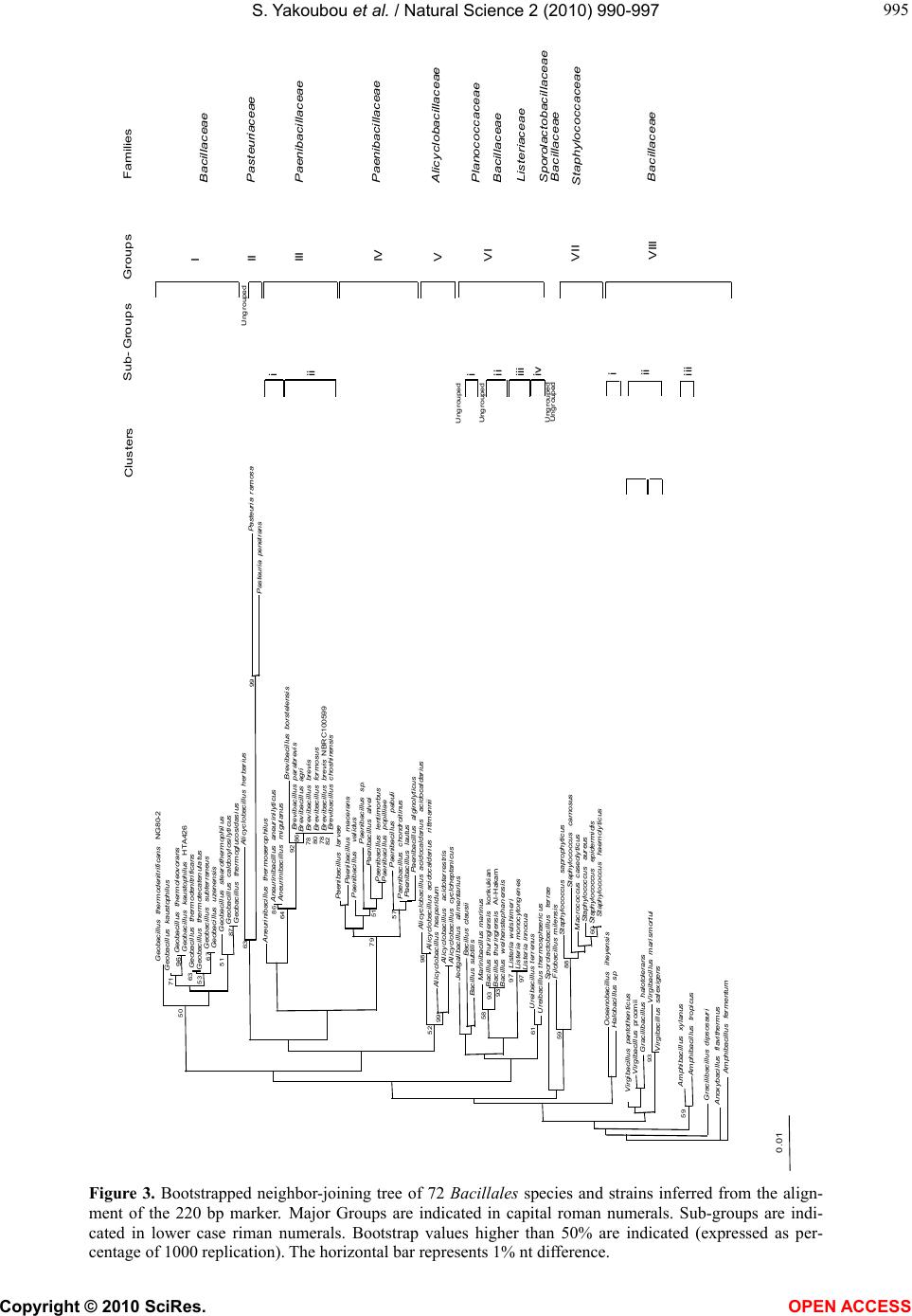
assessing the resolving power of this marker in recon-
structing the phylogeny at a lower taxa level: the Bacil-
lus cereus group, sensu lato.
Recently, in parallel, a similar maker was tested and
shown to be able to reconstruct the phylogeny of the
Class γ-proteobacteria [19].
REFERENCES
[1] Claus, D. and Berkeley, R.C.W. (1986) Genus Bacillus
Cohn, 1872. In: Sneath, P.H.A., Mair, N.S., Sharpe, M.E.
and J.G. Holt., Ed., Bergey’s Manual of Systematic Bac-
teriology, The Williams & Wilkins Co., Baltimore, 2,
1105-1139.
[2] Priest, F.G., Goodfellow, M. and Todd, C. (1988) A nu-
merical classification of the genus Bacillus. Journal of
General Microbiology, 134(7), 1847-1882.
[3] Ash, C., Farrow, A.E., Wallbanks, S. and Collins, M.D.
(1991) Phylogenetic heterogeneity of the genus Bacillus
revealed by comparative analysis of small-subunit-riboso-
mal RNA sequences. Letters in Applied Microbiology,
13(4), 202-206.
[4] Xu, D. and Côté, J.-C. (2003) Phylogenetic relationships
between Bacillus species and related genera inferred from
comparison of 3’ end 16S rDNA and 5’ end 16S-23S ITS
nucleotide sequences. International Journal of System-
atic and Evolutionary Microbiology, 53, 695-704.
[5] Ludwig, W., Schleifer, K-H. and Whitman, W.B. (2009)
Revised road map to the phylum Firmicutes. In: De Vos,
P., et al. Eds., Bergey’s Manual of Systematic Bacteriology,
2nd Edition, The Firmicutes, Springer-Verlag, New York,
3, 1-17.
[6] Garrity, G.M., Bell, J.A. and Lilburn, T. (2005) The Re-
vised Road Map to the Manual. In: Brenner, D.J., Krieg,
N.R., Staley, J.T. and Garrity, G.M., Eds., Bergey’s Man-
ual of Systematic Bacteriology, 2nd Edition, The Pro-
teobacteria, Part A, Introductory Essays. Springer, New
York , 2, 159-220.
[7] Sambrook, J. and Russell, D.W. (2001) Molecular Clon-
ing: A Laboratory Manual. 3rd Edition, Cold Spring
Harbor Laboratory Press, N.Y..
[8] Stephen, D., Jones, C. and Schofield, J.P. (1990) A rapid
method for isolating high quality plasmid DNA suitable
for DNA sequencing. Nucleic Acids Research, 18(24),
7463-7464
[9] Sanger, F., Nicklen, S. and Coulson, A.R. (1977) DNA
sequencing with chain-termination inhibitors. Proceed-
ings of the National Academy of Sciences, USA, 74,
5463-5467.
[10] Saitou, N. and Nei, M. (1987) The neighbor-joining
method: A new method for reconstructing phylogenetic
trees. Molecular Biology and Evolution, 4(4), 406-425.
[11] Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994)
CLUSTAL W: Improving the sensitivity of progressive
multiple sequence alignment through sequence weighting,
positions-specific gap penalties and weight matrix choice.
Nucleic Acids Research, 22(22), 4673-4680.
[12] Kimura, M. (1980) A simple method for estimating evo-
lutionary rate of base substitutions through comparative
studies of nucleotide sequences. Journal of Molecular
Evolution, 16(2), 111-120.
[13] Page, R.D.M. (1996) TreeView: An application to display
phylogenetic trees on personal computers. Computer Ap-
plication in the Biosciences, 12(4), 357-358.
[14] Page, R.D.M. (2000) TreeView—tree drawing software
for Apple Macintosh and Windows.
http://taxonomy.zoology.gla.ac.uk/rod/treeview.html
[15] Lu, J., Nogi, Y. and Takami, H., (2001) Oceanobacillus
iheyensis gen. nov., sp. nov., a deep-sea extremely
halotolerant and alkaliphilic species isolated from a depth
of 1050 m on the Iheya Ridge. FEMS Microbiology Let-
ters, 205(2), 291-297.
[16] Heyrman, J., Logan, N.A., Busse, H.J., Balcaen, A.,
Lebbe, L., et al. (2003) Virgibacillus carmonensis sp.
nov., Virgibacillus necropolis sp. nov. and Virgibacillus
picturae sp. nov., three novel species isolated from dete-
riorated mural paintings, transfer of the species of the
genus Salibacillus to Virgibacillus, as Virgibacillus
marismortui comb. nov. and Virgibacillus salexigens
comb. nov., and emended description of the genus Virgi-
bacillus. International Journal of Systematic and Evolu-
tionary Microbiology, 53, Part 2, 501-511.
[17] Zhilina, T.N., Garnova, E.S., Tourova, T.P., Kostrikina,
N.A. and Zavarzin, G.A., (2001) AmAmphibacillus fer-
mentum sp. nov. and Amphibacillus tropicus sp. nov.,
new alkaliphilic, facultatively anaerobic, saccharolytic
bacilli from Lake Magadi. Mikrobiologiya, in Russian,
70(6), 825-837. English Translation: Microbiology, 70(6),
711-722.
[18] Goto, K., Matsubara, H., Mochida, K., Matsumura, T.,
Hara, Y., et al. (2002) Alicyclobacillus herbarius sp. nov.,
a novel bacterium containing omega-cycloheptane fatty
acids, isolated from herbal tea. International Journal of
Systematic and Evolutionary Microbiology, 52(1), 109-
113.
[19] Yakoubou, S. and Côté, J.-C. (2010) Phylogeny of
γ-proteobacteria inferred from comparisons of 3’ end 16S
rRNA gene and 5’ end 16S-23S ITS nucleotide sequences.
Natural Science, 2(6), 535-543.