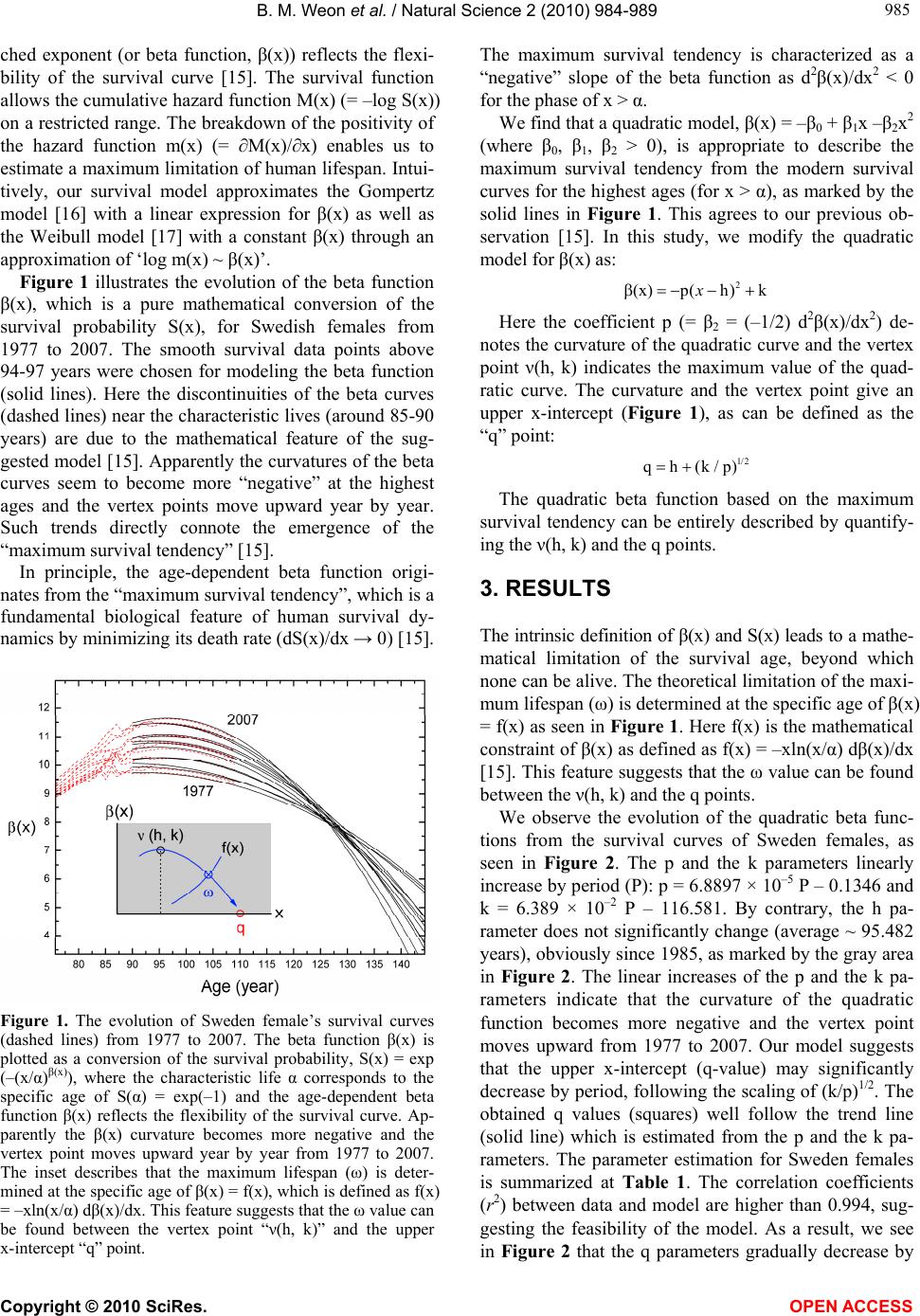
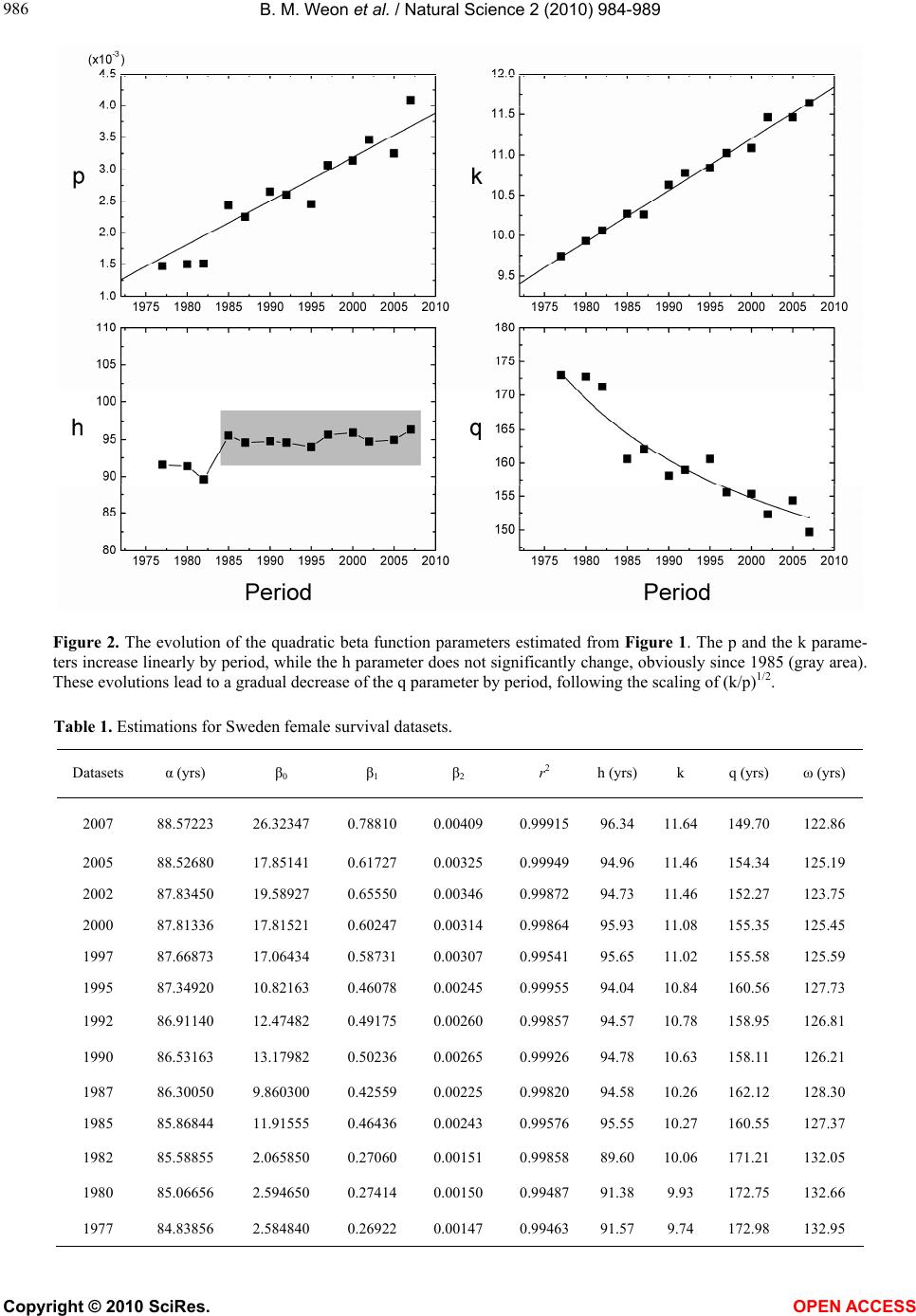
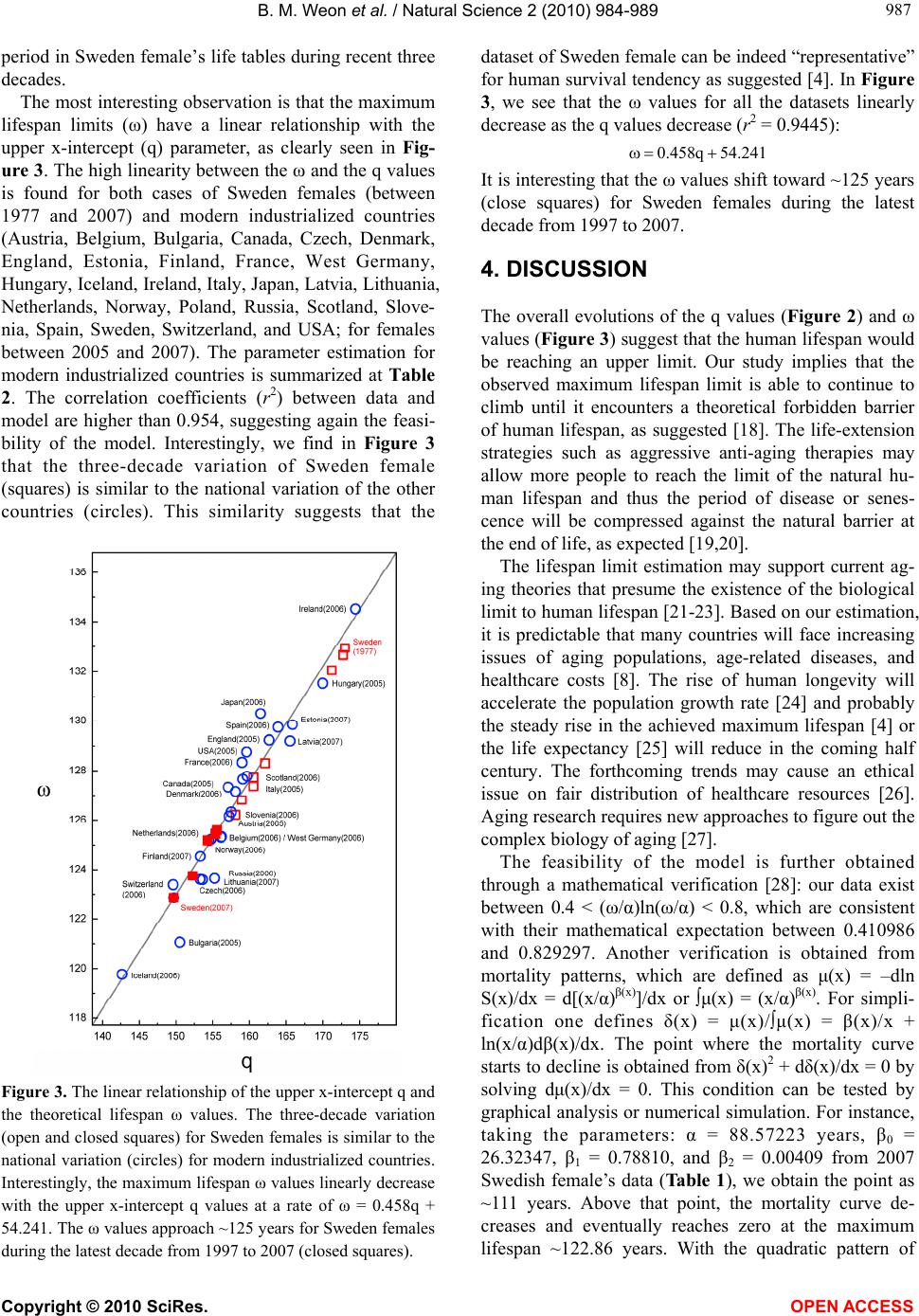
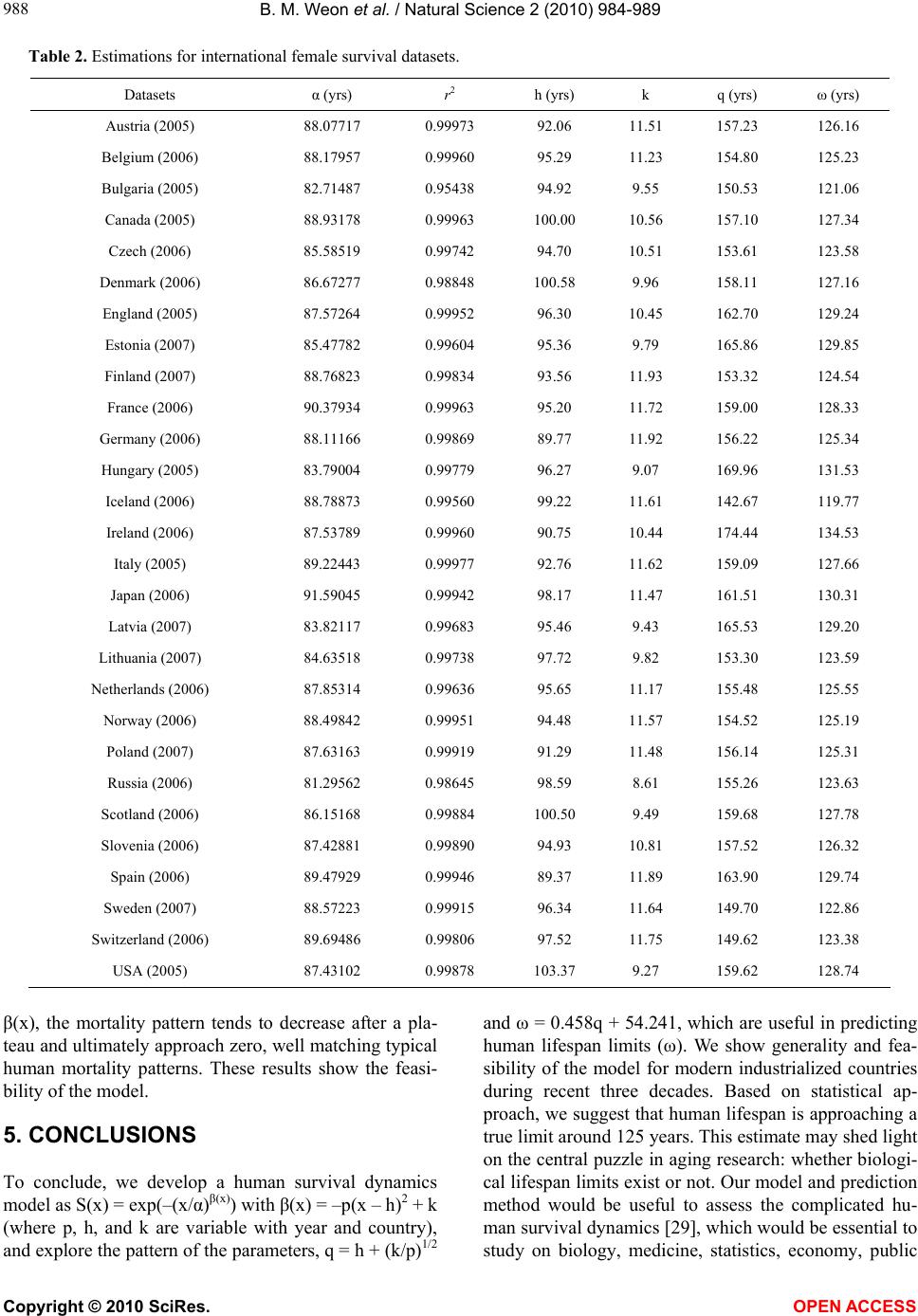
B. M. Weon et al. / Natural Science 2 (2010) 984-989
Copyright © 2010 SciRes. OPEN ACCESS
989
989
policy, and culture.
6. ACKNOWLEDGEMENTS
We are grateful to the Human Mortality Database (http://www.mortality.
org) for allowing anyone to access the demographic data for research.
This work was supported by the Creative Research Initiatives (Func-
tional X-ray Imaging) of MEST/NRF.
REFERENCES
[1] Vaupel, J.W. (1997) The remarkable improvements in
survival at older ages. Philosophical Transactions of the
Royal Society of London B, 352(1363), 1799-1804.
[2] Vaupel, J.W., Carey, J.R., Christensen, K., Johnson, T.E.,
Yashin, A.I., Holm, N.V., Iachine, I.A., Kannisto, V.,
Khazaeli, A.Z., Liedo, P., Longo, V.D., Zeng, Y., Man-
ton, K.G. and Curtsinger, J.W. (1998) Biodemographic
tra- jectories of longevity. Science, 280(5365), 855-860.
[3] Wilmoth, J.R. and Horiuchi, S. (1999) Rectangularization
revisited: Variability of age at death within human popu-
lations. Demography, 36(4), 475-495.
[4] Wilmoth, J.R., Deegan, L.J., Lundstrom, H. and Horiuch,
S. (2000) Increase of maximum life-span in Sweden,
1861-1999. Science, 289(5488), 2366-2368.
[5] Oeppen, J. and Vaupel, J.W. (2002) Broken limits to life
expectancy. Science, 296(5570), 1029-1031.
[6] Nemoto, S. and Finkel, T. (2004) Ageing and the mystery
at Arles. Nature, 429(6988), 149-152.
[7] Vijg, J. and Campisi, J. (2008) Puzzles, promises and a
cure for ageing. Nature, 454(7208), 1065-1071.
[8] Couzin, J. (2005) How much can human life span be
extended. Science, 309(5731), 83.
[9] Butler, R.N., Miller, R.A., Perry, D., Carnes, B.A., Wil-
liams, T.F., Cassel, C., Brody, J., Bernard, M.A., Par-
tridge, L., Kirkwood, T.B.L., Martin, G.M. and Ol-
shansky, S.J. (2008) New model of health promotion and
disease prevention for the 21st century. British Medical
Journal, 337(7662), 149-150.
[10] Mair, W. and Dillin, A. (2008) Aging and survival: The
ge- netics of life span extension by dietary restriction.
Annual Review of Biochemistry, 77(1), 727-754.
[11] Hayflick, L. (2000) The future of ageing. Nature,
408(6809), 267-269.
[12] Takahashi, Y., Kuro-o, M. and Ishikawa, F. (2000) Aging
mechanisms. Proceedings of the National Academy of
Sciences USA, 97(23), 12407-12408.
[13] Weon, B.M., Lee, J.L. and Je, J.H. (2005) A unified de-
cay formula for luminescence decays. Journal of Applied
Physics, 98(9), 096101.
[14] Weon, B.M., Je, J.H. and Lee, J.L. (2007) Lifetime dis-
persion in a single quantum dot. Applied Physics A, 89(4),
1029-1031.
[15] Weon, B.M. and Je, J.H. (2009) Theoretical estimation of
maximum human lifespan. Biogerontology, 10(1), 65-71.
[16] Gompertz, B. (1825) On the nature of the function ex-
pressive of the law of human mortality and on a new
mode of determining life contingencies. Philosophical
Transactions of the Royal Society of London A, 115
(1825), 513-585.
[17] Weibull, W.A. (1951) A statistical distribution function of
wide applicability. Journal of Applied Mechanics, 18(3),
293-297.
[18] Finch, C.E. and Pike, M.C. (1996) Maximum life span
predictions from the Gompertz mortality model. Journals
of Gerontology Series A: Biological Sciences and Medi-
cal Sciences, 51(3), B183-B194.
[19] Fries, J.F. (1980) Aging, natural death, and the compres-
sion of morbidity. New England Journal of Medicine,
303(3), 130-135.
[20] Vita, A.J., Terry, R.B., Hubert, H.B. and Fries, J.F. (1998)
Aging, health risks, and cumulative disability. New Eng-
land Journal of Medicine, 338(15), 1035-1041.
[21] Gavrilov, L.A. and Gavrilova, N.S. (2001) The reliability
theory of aging and longevity. Journal of Theoretical Bi-
ology, 213(4), 527-545.
[22] Carnes, B.A., Olshansky, S.J. and Grahn, D. (2003) Bio-
logical evidence for limits to the duration of life. Bio-
gerontology, 4(1), 31-45.
[23] Kirkwood, T.B.L. (2005) Understanding the odd science
of aging. Cell, 120(4), 437-447.
[24] Cohen, J.E. (2003) Human population: The next half
century. Science, 302(5648), 1172-1175.
[25] Olshansky, S.J., Passaro, D.J., Hershow, R.C., Layden, J.,
Carnes, B.A., Brody, J., Hayflick, L., Butler, R.N., Alli-
son, D.B. and Ludwig, D.S. (2005) A potential decline in
life expectancy in the United States in the 21st century.
New England Journal of Medicine, 352(11), 1138-1145.
[26] Farrant, A. (2009) The fair innings argument and in-
creasing life spans. Journal of Medical Ethics, 35(1),
53-56.
[27] Kirkwood, T.B.L. (2008) Gerontology: Healthy old age.
Nature, 455, 739-740.
[28] Lakshminarayanan, E.S. and Sumathi, M. On extended
Weibull model. International Journal of Applied Mathe-
matics and Applications, in press.
[29] Kaplan, H.S. and Robson, A.J. (2009) We age because
we grow. Proceedings of the Royal Society B, 276(1663),
1837-1844.