Paper Menu >>
Journal Menu >>
 Engineering, 2012, 5, 30-34 doi:10.4236/eng.2012.410B008 Published Online October 2012 (http://www.SciRP.org/journal/eng) Copyright © 2012 SciRes. ENG Mutations in Caprine DGAT1 and STAT5A Genes were Associated with Milk Production Traits ——Combined Effects of DGAT1 and STAT5A Genes on Milk Yield and Fat Xiaopeng An, Jinxing Hou, Haibo Zhao, Chunmei Zhu, Quanmei Yan, Y uxuan Song, Jiangang Wang, Binyun Cao College of Animal Science and Technology, Northwest A&F University, Yangling, Shaanxi P.R. China Email: caobinyun@yahoo.com.cn Received 2012 ABSTRACT In this study, polymorphisms of the DGAT1 and STAT5A genes were detected in 528 individuals from Xinong Saanen and Gua- nzhong goat breeds by PCR-RFLP, PCR-SSCP and DNA sequencing methods. Three allelic variants were identified: DQ380250: g.407_408insC, AJ237937: g.6798C>T and g.6852C>T in both breeds. At g.407_408insC locus, the frequencies of C1 allele were 0.79–0.85, and frequencies of C2 allele were 0.21–0.15. At g.6852C>T locus, frequencies of C3 allele were 0.70–0.72, and frequen- cies of T3 allele were 0.30–0.28. Compared with goats with C1C1 and C3C3, those with C1C2 and C3T3 genotypes had significant ef- fects on milk yield and fat percentage (P<0.05), respectively. The result showed that does with C1C1C3T3 and C1C2C3T3 had higher milk yield than those with C1C2C3C3 (P < 0.05). In addition, the combined effect of C1C2C3T3 on milk fat percen tage was th e highest in comparison with other combination genotypes (P<0.05). Keywords: Dairy Goat; Milk Production Traits; Fat Percentage; Pedi g r e e 1. Introduction Milk production traits are of fundamental importance in live- stock production and the related economy [1]. Selection aimed at increas ing th e frequency of al leles with a p osit ive effect on a given trait was initiated by geneticists [2]. Meanwhile, variation of either candidate genes for production traits or linked genetic markers h as informed t he basic biolo gy of milk production and composition, and encouraged the use of gene for marker as- sisted selection (MAS) in livestock [3]. In general, identifying and validating genetic markers for milk production traits is the initial and crucial step to establish a MAS system. Diacylglycerol acyltransferases (DGATs) catalyse the final step of the triacylglycerol (TAG) biosynthesis of the Kennedy pathway [4]. Two genes (DGAT1 and DGAT2) have been shown to encode DGATs. Both genes encode membrane-bound proteins, with no sequence homology to each other [5]. DGAT1 gene was the first identified gene encoding a protein with DGAT activity [6]. Diacylglycerol acyltransferase1 (DGAT1) was identified as one underlying quantitative trait locus (QTL) for milk production traits in the centromeric region of the bo- vine chromosome 14 [7, 8]. The signal transducers and activa- tors of transcription (ST ATs ), a family of transcription factors, mediate the actions of a variety of peptide hormones and cyto- kines [9]. STAT5, also known as mammary gland factor ( MGF), was discovered initially as a PRL-induced transcription factor [10]. It is a key intracellular mediator of prolactin signalling and can activate trans cription of milk p rotein genes in response to prolactin [10, 11]. STAT5 exists in two isoforms – A and B, which differ by a few amino acids in the carboxylic end of the protein molecule; separate genes code both of them [12]. In cattle, the STAT5A and STAT5B genes wer e located close to each other (within 40 Kb) at chromosome 19 [13]. Ant o ni ou et al. (1999) described two SSCP variants of the gene fragment that encodes the SH2 domain in bovine STAT5A protein [14]. Brym et al. (2004) detected a new SNP (A/G) located in the intron 9 of STAT5A gene at position 9501 [15]. The aim of this study was to investigate SNPs in DGAT1 and STAT5A genes, and anal yze the combined effect of DGAT1 and STAT5A genes on milk production traits to provide the theoretical basis for goat breeding. 2. Materials and Methods 2.1. Animals and Genomic DNA Isolation Blood samples were obtained from 528 goats belonging to two breeds: Xinong Saanen (SN, n=285) and Guanzhong (GZ, n=243). They were reared in Qianyang county and Zhouzhi county of Shaanxi province, respectively. Health, fertility and milk recording was carried out by dairymen and veterinarians. Data was recorded in winter and spring parturitions of 2008 to 2011. Milk yi e ld s fr o m first to third lactation were standard ized to 30 0 days in milk. For milk analysis, a milk sample was taken from each animal once per month throughout the third lactation, sampling first at least 20 days after parturition to exclude the risk of contamination with colostrum. Goats were milked twice a day at constant intervals an d a 10 ml sa mple from each milk- ing session was mixed for the analysis. Milk constituents (pro- tein, lactose and fat) were determined with an ultrasonic S60SEC milk analyzer (Milkotronic Company, Nova Zagora, Bulgaria). Five milliliters blood per goat were collected asep ti-  X. P. AN ET AL. Copyright © 2012 SciRes. E NG 31 cally from the jugular vein and kept in a tube containing anti- coagulant ACD (citric acid:sodium citrate:dextrose – 10: 27: 38). The genomic DNA was extracted from white blood cells using standard phenol-chloroform extraction protocol [16]. 2.2. PCR Amplification According to bovine DGAT1 and STAT5A genes (GenBank accession no. AJ318490 and AJ237937), fourteen pairs of pri- mers were designed to amplify goat DGAT1 and STAT5A genes. Pairs of primer 1 and 2 are shown in Table 1. Other primer pairs with no polymorphism detected in their amplifica- tion regions are not listed. The 25 μL volume contained 50 ng genomic DNA, 12.5 µL 2 × reaction mix (including 500 µM dNTP each; 20 mM Tris–HCl; pH 9; 100 mM KCl; 3 mM MgCl2 ), 0.5 µM of each primer, and 0.5 units of Taq DNA polymerase. The cycling protocol was 5 min at 95°C, 35 cycles of denaturing at 94°C for 30 s, annealing at 59°C (primer pair 1) and 63°C (primer pair 2) for 30 s, extending at 72°C for 30 s, with a final extension at 72°C for 10 min. 2.3. SNP Genotyping and Sequencing The SSCP analysis of PCR products of primer pair 2 refers to An et al. (2011) [17]. In addition , PCR products (5μl) o f pri mer pair 2 wer e mixed with 1 μl 10 × buffer, 3 U Eco81Ⅰ(TaKaRa, Dalian, China) and 3.5 μl sterilized ddH2O, and then incubated for 1.5 h at 37°C. Digested products were subjected to PAGE (80 × 73 × 0.75 mm) in 1 × TBE buffer and constant voltage (110 V) fo r 1.5 h. After the polymorphisms wer e detected, am- plicons representing unique banding patterns were sequen ced in both directions in ABI 377 DNA analyzer (Applied Biosystems , Foster , California, U SA) and the sequences were analyz e d with DNAstar software (version 7.1) and Blast in NCBI (National Center for Biotechnology Information). 2.4. Statistical Analysis The allelic frequenci es, heterozygosi ty (He) and polymorphism information content (PIC) were calculated using Cluster-analy- sis so ft wa r e (version 1.2). Milk production traits analyzed in the current study included milk yield, milk protein, lactose and fat. Stati stical analysis was perfo rmed using univariate an alysis in the general linear model procedure of SPSS 16 statistical softwa r e. The linear model applied was: Yiknjlm = µ + Gi + Bk + Pn + Nj + (PG) ni+ Sl + Eiknjlm (model 1) where Yik njlm is the trait measured on each of the iknjlmth animal, µ is the overall population mean, Gi is the fixed effect asso- ciated with the ith genotype, Bk is the fixed effect associated with the kth breed, Pn is the fixed effect associated with the nth parity, Nj is the fixed effect associated with the jth number of kids born, (PG)ni is the in teractio n bet ween the nth parity and ith genotype. Sl is the random effect ass ociated with the lth sire, and Eiknjlm is the random error. The combined effects of DGAT1 and STAT5A genes o n mil k p ro du ction trai ts were an al yzed with the following model: Yik njlm = µ + Ci + Bk + Pn + Nj + (PC) ni + Sl + Eiknjlm (model 2) where Yiknjlm, µ, Bk, Pn, Nj and Sl are the same as shown for model 1, Ci is the fixed effect associated with the ith combina- tion genotype, and (PC)ni is the in teracti on b etween the nth par- ity and ith combination genotype. 3. Results 3.1. SNPs Identification and Genot ypes The bands of different genotypes are shown in Figure 1A and 1B. Comparisons among these nucleotide sequences of differ- ence genotypes indicated that one base insertion (g.407_ 408insC, GenBank accession no. JF781126) was detected i n th e Table 1. Primer sequences and information on goat DGAT1 and STAT5A genes. Gene Pri me r Sequence (bp) Ta (℃) Amplicon Product size (bp) DGAT1 P1 F: 5-AGGAACTCGG AGTCCATCAC-3 59 Exon 14-16 328 R: 5- TGAAGGCCCAGAGGCGGAAC-3 STAT5A P2 F: 5- CTGCAGGGCTGTTCTGAGAG-3 63 Exon 7 215 R: 5- TGGTACC AGGACTGTAGCACAT-3 Note: Fragments includ ing 36 bp of C3T3 genotype were invisib le Figure 1. SNP d et ect i on o f PCR products at g.407_408insC (A) and g.6852C>T (B) loci for tw o goat breeds. 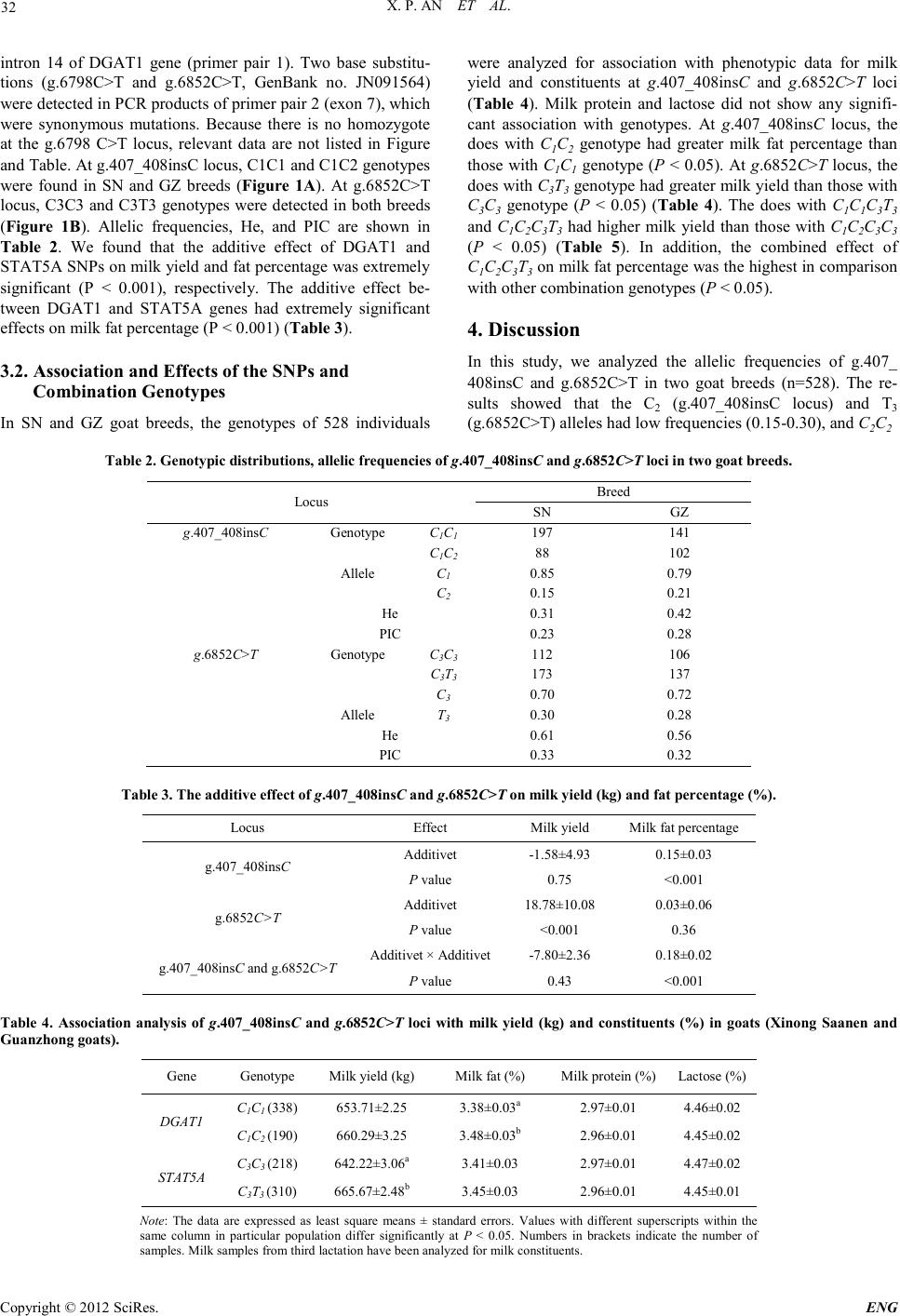 X. P. AN ET AL. Copyright © 2012 SciRes. ENG 32 intron 14 of DGAT1 gene (primer pair 1). Two base substitu- tions (g.6798C>T and g.6852C>T, GenBank no. JN091564) were detected in PCR products of primer pair 2 (exon 7), which were synonymous mutations. Because there is no homozygote at the g.6798 C>T locus, relevant data are not listed in Figure and Table. At g.407_408insC locus, C1C1 and C1C2 genotypes were found in SN and GZ breeds (Figure 1A). At g.6852C>T locus, C3C3 and C3T3 genotypes were detected in both breeds (Figure 1B). Allelic frequencies, He, and PIC are shown in Table 2. We found that the additive effect of DGAT1 and STAT5A SNP s on milk yield and fat p ercentag e was extr emely significant (P < 0.001), respectively. The additive effect be- tween DGAT1 and STAT5A genes had extremely significant effects on milk fat per centage (P < 0.001) (Table 3 ). 3.2. Association and Effects of the SNPs and Comb ination Genotypes In SN and GZ goat breeds, the genotypes of 528 individuals were analyzed for association with phenotypic data for milk yield and constituents at g. 407_408insC and g.6852C>T loci (Table 4). Milk protein and lactose did not show any signifi- cant association with genotypes. At g.407_408insC locus, the does with C1C2 genotype had greater milk fat percentage than those with C1C1 genotype (P < 0.05). At g.6852C>T locus, the does with C3T3 genotype had greater milk yield than those with C3C3 genotype (P < 0.05) (Table 4). The does with C1C1C3T3 and C1C2C3T3 had higher milk yield than those with C1C2C3C3 (P < 0.05) (Table 5). In addition, the combined effect of C1C2C3T3 on milk fat percen tage was the highest in comparison with other combination genotypes (P < 0.05). 4. Discussion In this study, we analyzed the allelic frequencies of g.407_ 408insC and g.6852C>T in two goat breeds (n=528). The re- sults showed that the C2 (g.407_408insC locus) and T3 (g.6852C>T) alleles had low frequencies ( 0.15-0.30), and C2C2 Table 2. Genotypic d istributions, allelic frequencies of g.407_408insC and g.6852C>T lo ci in two g o at breeds. Loc us Breed SN GZ g.407_408insC Genotype C1C1 197 141 C1C2 88 102 Allele C1 0.85 0.79 C2 0.15 0.21 He 0.31 0.42 PIC 0.23 0.28 g.6852C>T Genotype C3C3 112 106 C3T3 173 137 C3 0.70 0.72 Allele T3 0.30 0.28 He 0.61 0.56 PIC 0.33 0.32 Table 3. The additive effect of g.407_408insC and g.6852C>T on milk yield (kg) and fat percentage ( %). Loc u s Effect Milk yield Milk fat percentage g.407_408insC Additivet -1.58±4.93 0.15±0.03 P value 0.75 <0.001 g.6852C>T Additivet 18.78±10.08 0.03±0.06 P value < 0.001 0.36 g.407_408insC and g.6852C>T Additivet × Additivet -7.80±2.36 0.18±0.02 P value 0.43 <0.001 Table 4. Association analysis of g.407_408insC and g.6852C>T loci with milk yield (kg) and constituents (%) in goats (Xinong Saanen and Guanzhong goats). Gene Genotype Milk yield (kg) Milk fat (%) Milk protein (%) Lactose (%) DGAT1 C1C1 (338) 653.71±2.25 3.38±0.03a 2.97±0.01 4.46±0.02 C1C2 (190) 660. 29±3.25 3.48±0.03b 2.96±0.01 4.45±0.02 STAT5A C3C3 (218) 642.22±3.06a 3.41±0.03 2.97±0.01 4.47±0.02 C3T3 (310) 665. 67±2.48b 3.45±0.03 2.96±0.01 4.45±0.01 Note: The data are expressed as least square means ± standard errors. Values with different superscripts within the same column in particular population differ significantly at P < 0.05. Numbers in brackets indicate the number of samples. Milk samples from third lactation have been analyzed for milk constituents.  X. P. AN ET AL. Copyright © 2012 SciRes. E NG 33 Table 5. Combined effects of D GAT1 and S TA T5A genes on milk yield (kg) a nd fat percentage ( %) in goats (Xinong Saanen and Guanzhong goats). Genotypic combination Milk yield (kg) Milk fat (%) Milk protein (%) Lactose (%) C1C1C3C3 (147) 642.10±3.65a 3.44±0 .03a 2.96±0.0 1 4.46±0.03 C1C1C3T3 (191) 664.35±3.32b 3.38±0.03a 2.95±0.01 4.42±0.02 C1C2C3C3 (70) 645.23±5.48 3.41±0.05a 2.97±0.02 4.45±0.04 C1C2C3T3 (120) 666.21±3.89b 3.59±0.04b 2.93±0.01 4.47±0.03 Note: The data are expressed as least square means ± standard errors. Values with different superscripts within the same column in particular ge ner a t io n dif fer s ignific a ntl y at P < 0.05. Numbers in brackets indicate the number of s ample s . M il k s a mple s from thi rd la c ta t io n have be en a na l yz e d fo r mil k con s t i t ue nt s . (inset homozygote) and T3T3 (mutation homozygote) genotypes were not obs erved, respectively at the two loci in SN and GZ goat br eeds. Flisikowski et al. (2003) reported C→T at position 6853 within the exon 7 of STAT5A gene and they found the TT genotype only in Polish native breeds (Polish Red and Polish White-Back cattle) [1 8]. We consider that the results can be explained by the following two reasons. (1) There is a lower frequency for missing genotypes, and the samples are small. (2 ) The missing genotypes of the two loci have negative ef fects on individual performance, so the individuals with missing geno- types have been eliminated in breeding process. We firstly revealed the significant association of DGAT1 in- del (g.407_408insC) and STAT5A SNP (g.6852C>T) with milk yi e ld and fat percentage in Chinese dairy goats (P < 0.05). Al- though the mutations of g.407_408insC and g.6852C>T loci do not concern the coding region and the change of amino acid, they possibly influence the stability of the mRNA, and can affect th e mechani sm of mRNA deadenylation and degradation [19-21]. Linkage disequilibrium with the causal mutation pos- sibly affects the variation of milk production traits in goat [22]. Previous studies have demonstrated the importance of DGAT1 and STAT5 A genes in milk production traits in cattle [7,8, 23]. DGAT1 candidate gene was found to have a significant effect not only on milk yield and component traits but also on the metabolism of intramuscular fat [7, 8 , 24] . Amills et al. (20 07) indicated T to C substitution at the intron 16 of goat DGAT1 gene could be used as a marker in association studies with milk traits [25]. Dario et al. (2009) studied the effect of STAT5A/ AvaI polymorphism on growth performance traits in Podolica bulls and suggested the superiority of C allele for growth per- formances because both CC and CT bulls tended to show a higher live weight and a faster growth in comparison with TT animals [11]. Sadeghi et al. (2009) studied the association be- twe e n this polymorphism of STAT5A gene and the breeding values of milk production traits in 134 Iranian Holstein bulls [26]. Dario et al. (2009) reported a substitution C→T at posi- tion 6853 of STAT5A gene led to three genotypes (CC, CT and CT), and the cows with CC genotype had higher milk yield and protein content than those with CT genotype [27]. The bio- chemical and physiological functions, together with the results obtained in our study, indicate that the DGAT1 and STAT5A genes might play important roles affecting milk production traits in goat. Genotypic value includes additive effect and do- minant effect. Additive effect could be truly transmitted to offspring, so it is the focus of marker-assisted selection [28]. In this study, we took into account additive effect between SNP loci and milk production traits. The result showed the additive effect of g.407_408insC and g.6852C>T on milk yield and fat percentage was extre mely signi fican t (P < 0.001), resp ectively. Compared with single SNP analysis, combination genotypes analysis provides more information on gene interactions. Mul- tipl e lo cus anal ysis u s ed in th e s tudy revealed th at the combined effect of DGAT1 g.407_408insC and S TAT5A g.6852C>T sig- nificantly affected milk yield and fat percentage. Kong et al. (2007) indicated no significant effects on economic traits in Hanwoo cattle were found in the separate analysis of K232A and T11993C polymorphisms of DGAT1 gene, but the interac- tion between K232A and T11993C showed a significant effect (P < 0.005) on marbling score [24]. Based on the above con- siderations, we thought milk production traits were subjected to the impacts of g.407_408insC and g.6852C>T loci, and there was an i nteracti on between both loci. 5. Acknowledgements This study was supported by the National Support Program of China (2011BAD28B05-3) and Science and Technology Inno- vation Project of Shaanxi Province (2011KTCL02-09) REFERENCES [1] X.P. An, S.G. Son g, J. X. Hou, C.M. Zhu, J.X. Peng, X.Q. Liu, et al., “Polymorphism identification in goat DGAT2 gene a nd a sso- ciation analysis with milk yield and fat percentage,” Small Ru- minant Research, vol. 100, pp. 107-112, 2011. [2] J.C. Dekkers, “Commercial application of marker- and gene-assisted selection in livestock: strategies and lessons,” Journal of Animal Science, vol. 82, pp. E313-328, 2004. [3] I. Parmentier, D. Portetellea, N. Gengler, A. Prandic, C. Bertoz- zia, L. Vleuricka, et al., “Can di da t e gene ma r k ers as so cia t ed wi t h somatotropic axis and milk selection,” Domestic Animal Endo- crinology, vol. 17, pp . 139-148, 1999. [4] P. Hat zopoulos, G. B anila s, M. Karampelias, I. Makariti, and A. Kou r ti , “The olive DGAT2 gene is developmentally regulated and shares overlapping but distinct expression patterns with DGAT1,” Journal of Experiment al Botany, val. 62, pp. 521-532, 2011. [5] K. Giannoulia, K. Haralampidis, Z. Poghosyan, D.J. Murphy, and P. Hatzopoulos, “Differential expression of diacylglycerol acyltransferase (DGAT) gen es in olive t iss ues,” B io chemi cal So- ciety Transactions, vol. 28, pp. 695-697, 2000. [6] S. Cases, S.J. Sto ne, P. Zhou, E. Yen, B. Tow, K.D. Lardizabal, et al., “Cloning of DGAT2, a second mammalian diacylglycerol acyltransferase and related family members,” Journal of Biolog-  X. P. AN ET AL. Copyright © 2012 SciRes. ENG 34 ical Che m istry, vol. 276, pp. 38870-38876, 2001. [7] B. Grisart , W. Co ppi eters , F. Farnir, L. Karim, C. For d, P . Berzi, et al., “Positional candidate cloning of a QTL in dairy cattle: Identification of a missense mutation in the bovine DGAT1 gene with major effect on milk yield and composition,” Genome Re- search, vol. 12, pp. 222-231, 2002. [8] A. Winter, W. Kramer, F.A.O. Werner, S. Kollers, S. Kata, G. Durstewitz, et al., “Association of a lysine-232/alanine poly- morphism in a bovine gene encoding acyl-CoA : diacylglycerol acyltransferase (DGAT1) with variation at a quantitative trait locus for milk fat content,” Proceedings of the National Acade- my of Scien ces, vol. 99, p p. 9300-9305, 2002. [9] J.E. Darnell, I.M. Kerr, and G.R.Stark, “Jak-Stat pathways and transcriptional activation in response to IFNs and other extracel- lular signaling proteins,” Science, vol. 264, pp. 1415-1421, 1994. [10] H. Wakao, F. Gouilleux, and B.Grone r , “Mammary-gland factor (Mgf) is a novel member of the cytokine regulated transcription factor gene family and confers the prolactin response,” Embo Journal, vol. 13, pp. 2182-2191, 1994. [11] C. Dario, M. Selvaggi, D. Carnicella, and G. Bufano, “STAT5A/AvaI polym orphism in Podoli ca bulls and its effect on growth performance traits,” Livestock Science, vol. 123, pp. 83-87, 2009 [12] H.M. S eyfert, C. Pitra, L. Meyer, R.M. Brun ner, T.T. Wheel er, A. Molenaar, et al., “Molecular characterization of STAT5A- and STAT5B-encoding genes reveals extended intragenic sequence homogeneity in cattle and mouse and different degrees of diver- gent evolu ti on of vari ous domains,” Journ al of Molecu lar Evolu- tion, vol. 50, pp. 550-561, 2000. [13] A. Molenaar, T.T. Wheeler, J.Y. McCracken, and H.M. Seyfert, “The STAT3-encoding gene resides within the 40 kbp gap be- tween the STAT5A- and STAT5B-encoding genes in cattle,” An- imal Genetics, vol. 31, pp. 339-340, 2000 [14] E. Antoniou, B.J. Hirts, M. Gros z , and J. Skidmorec, “A single stra nd conformat ion polymorp his m in the bovine gene STAT5A,” Animal Genetics, vol. 30, pp. 225-244, 1999 [15] P. B rym, S. Kamiński, and A. Ruść, “New SSCP polymorphism within bovine STAT5A gene and its associations with milk per- forma nc e trai ts in Black -and-Whit e and Jer sey ca tt le,” Journa l of Applied Genetics, vol. 45, pp. 445-452, 2004. [16] R. Mullenbach, P.J. Lagoda, and C. Welter, “An efficient salt chloro-form extraction of DNA from blood and tissue,” Trends in Genetics, vol. 5, pp. 391, 1989. [17] X.P. An, J.G. Wang, J.X. Hou, H.B. Zhao, L. Bai, G. Li, et al., “Polymorphism identification in the goat MSTN gene and a ssoci- ation analysis with growth traits,” Czech Journal of Animal Science, vol. 56, pp. 529-535, 2011. [18] K. Flisikowski, J. Oprzdek, E. Dymnicki, and L.Zwi e rzcho ws k i, “New polymorphism in bovine STAT5A gene and its a ssociation with meat production traits in beef cattle,” Animal Science Pa- pers and Reports, vol. 21, pp. 147-157, 2003. [19] D.R. Gallie, and T.E. Young, “The re gu lat ion of gen e-expression in transformed maize aleurone and endosperm protop- lasts-analysis of promoter activity, intron enhancement, and messenger-RNA untranslated regions on expression ,” Plan t Phy- siology, vol. 106, pp. 929-939, 1994. [20] J.Q. Clement, S. Maiti, and M.F. Wilkinson, “Localization and stability of introns spliced from the Pem homeobox gene,” Jour- nal of Biological Chemistry, vol. 276, pp. 16919-16930, 2001. [21] Z.E. Sauna, and C. Kimch i-Sarfaty, “Understanding the contri- bution of synonymous mutations to human disease,” Nature Re- views Genetics, vol. 12, pp. 683-691, 2011. [22] J.H.J. Van der Werf, K. Marshall, and S. Lee, “Methods and experimental designs for detection of QTL in sheep and goats,” Small Ruminant Research, vol. 70, pp. 21-31, 2007. [23] H. Khatib, R.L. Mons on, V. Schutzkus, D . M. Ko h l , G.J. M. Rosa, J.J. Rutledge, “Mutations in the STAT5 A gene ar e ass o ciated with embryonic survival and milk composition in cattle,” Journal of Dairy Science, vol. 91, pp. 784-793, 2008. [24] H.S. Kong, J. D. Oh , J.H. Lee, D.H. Yoon, Y.H. Choi , B.W. Ch, et al., “Association of sequence variations in DGAT1 gene with economic traits in Hanwoo (Korea cattle),” Asian-Australasian Journal of Animal Sciences, vol. 20, pp. 817-820, 2007. [25] M. Amills, A. Angi olil lo, B . Urrutia, A. Dom enec h , Y. Sastre, B. Badaoui, et al., “Identification of a single nucleotide polymor- phism at intron 16 of the caprine acyl-coenzyme A: Diacylgly- cerol acyltransferase 1 (DGAT1) gene,” Journal of Dairy Re- search, vol. 74, pp. 47-51, 2007. [26] M. Sadeghi, M.M. Shahrbabak, G.R. Mianj, and A.N. Javaremi, “Polymorphism at locus of STAT5A and its association with breeding values of milk production traits in Iranian Holstein bu lls,” Lives toc k Science, vol. 123, pp. 97-100, 2009. [27] C. Dario, M. Selvaggi, G. Normanno, G.V. Celano, and M. Dar io , “Genetic polymorphism of STAT5A protein: relationships with production traits and milk composition in Italian Brown cattle,” Journal of Dairy Research, vol. 76, pp. 441-445, 2009. [28] X.P. An, D. Ha n, J.X. Hou , G. Li , Y.N. Wa ng, L. Li, et al. , “Po- lymorph ism of exon 2 of FSHβ gene and its relationship with re- production performance in two goat breeds,” Agricultural Science s in C hi na, vol. 9, pp. 880-886, 2010. |