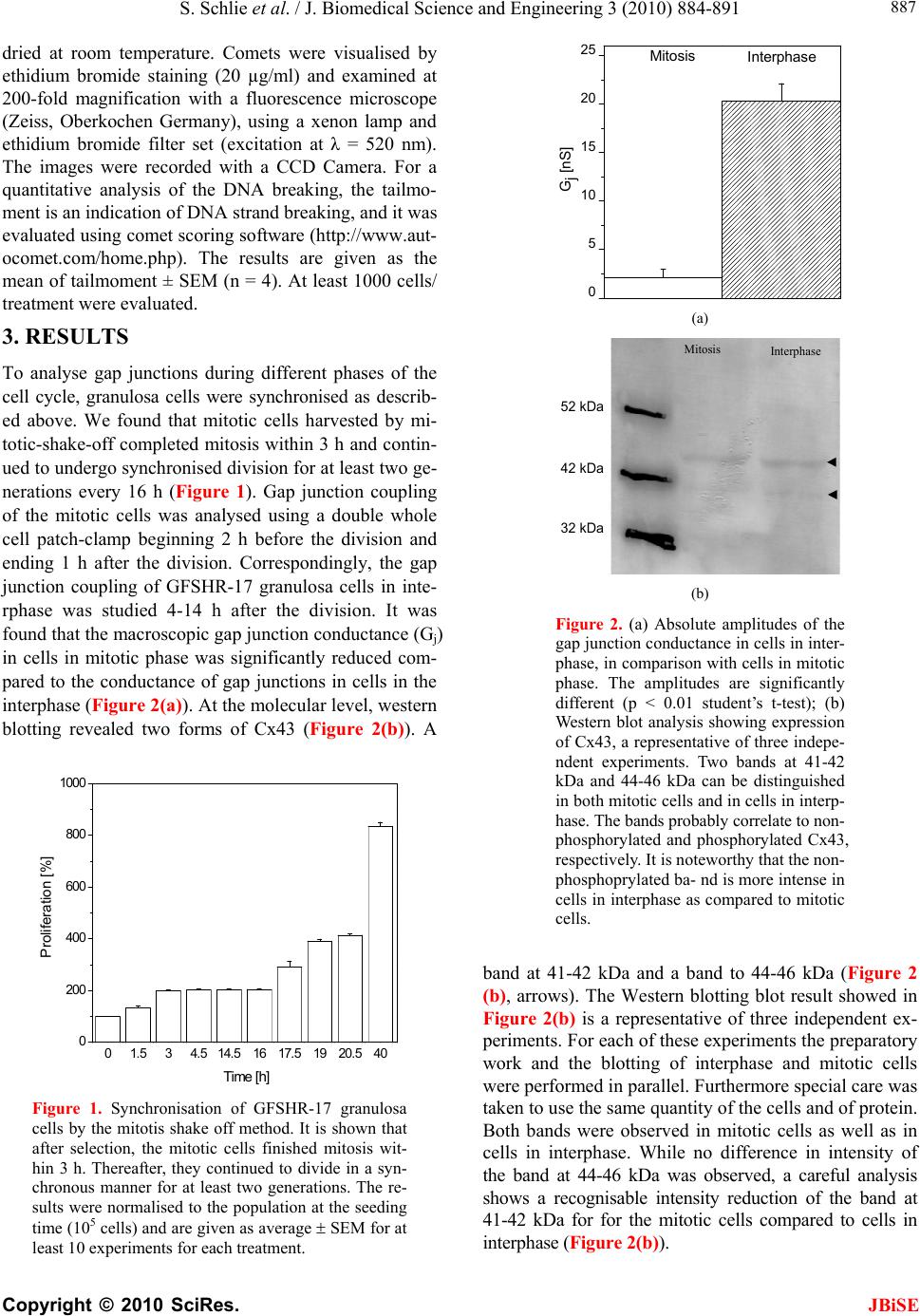
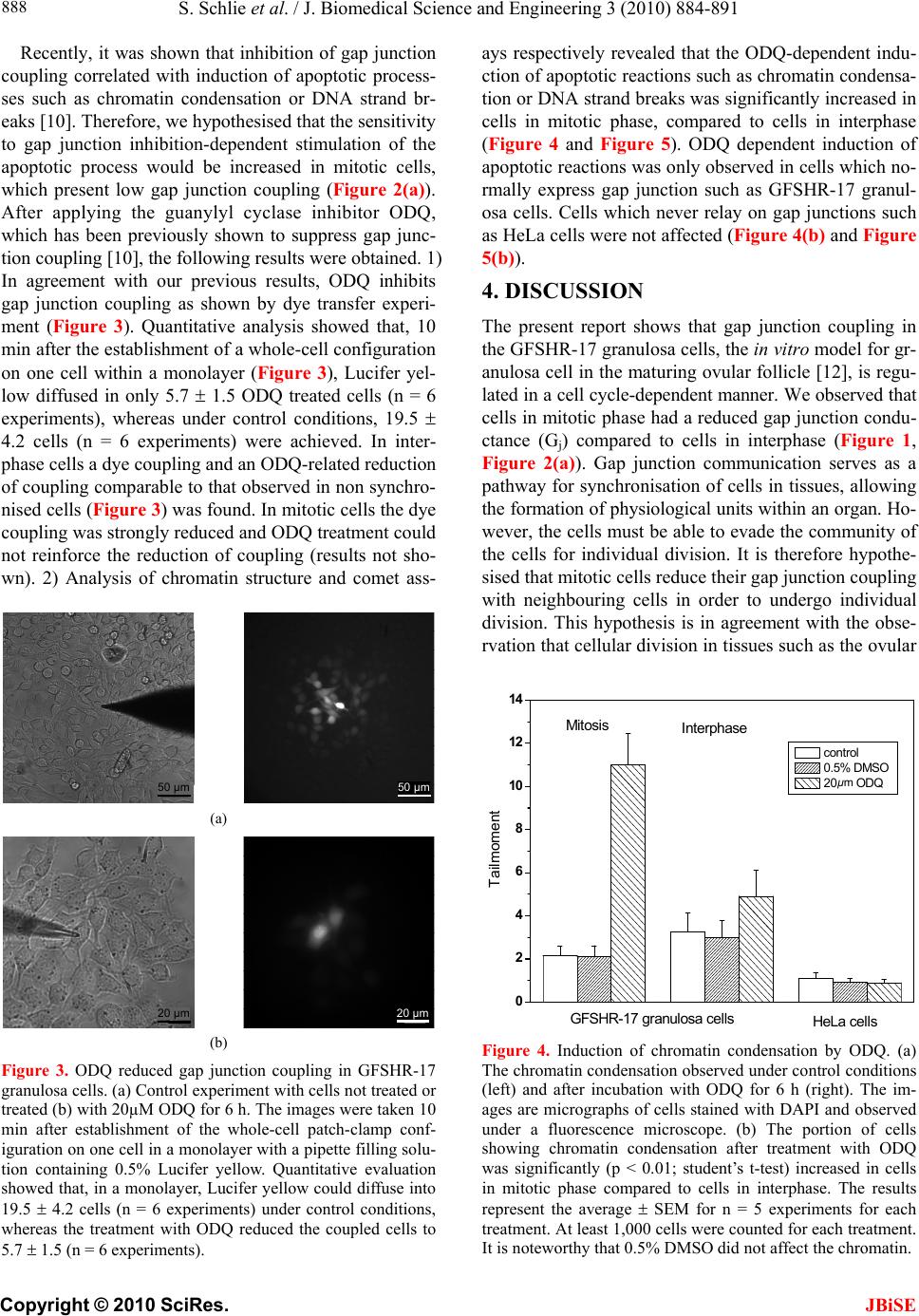
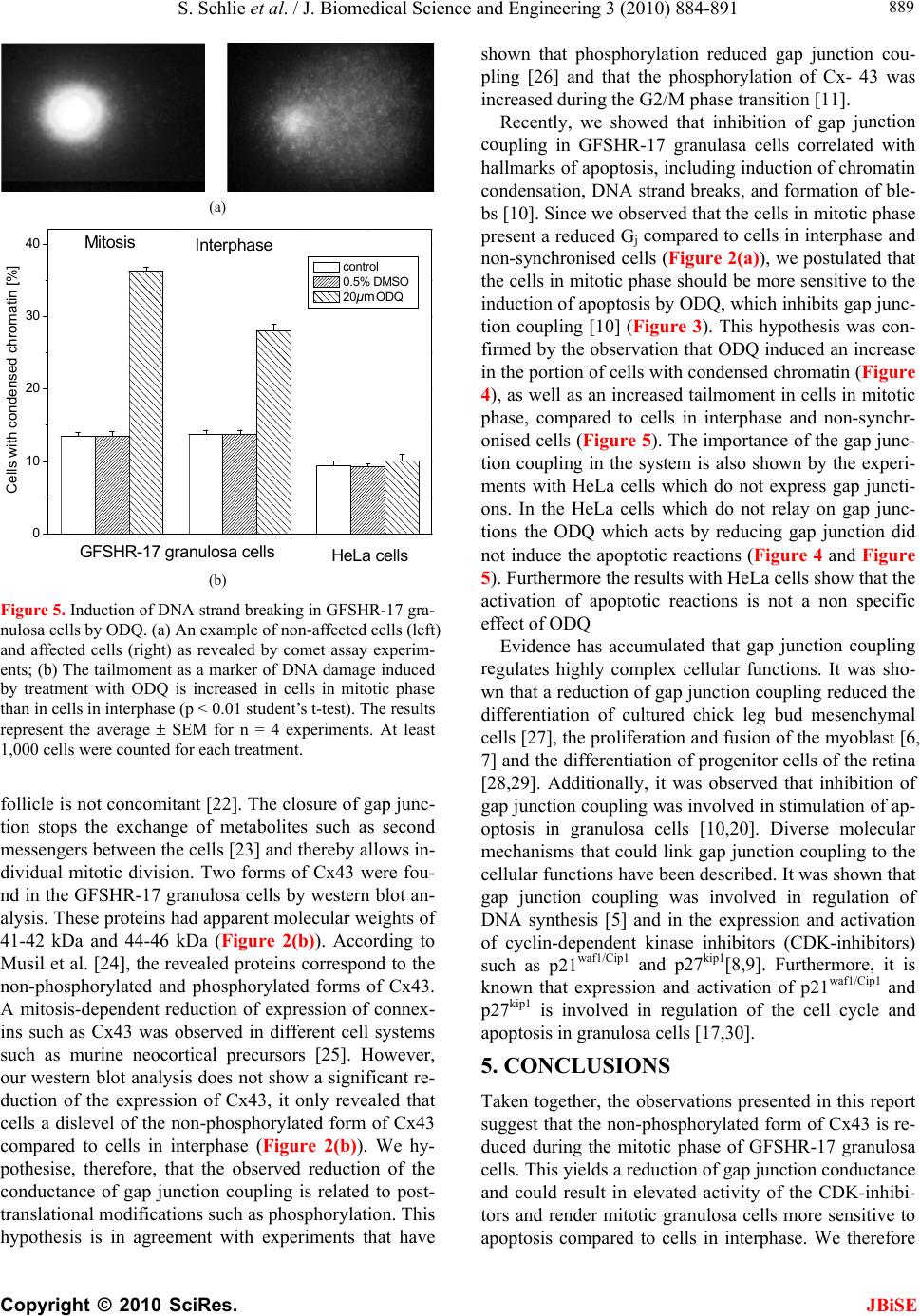
S. Schlie et al. / J. Biomedical Science and Engineering 3 (2010) 884-891
Copyright © 2010 SciRes. JBiSE
890
LEDGEMENTS
excel-
y supported by the project NANOTOME and
th
uropean Graduate College; I
fer
1) Emerging issues of connexin chan-
2003) Astrocytic
, Dang, X., Ping, P., Fandrich, R.R., Nickel,
, Becker, D.L., Dux, L., Stelkovics, E., Kren-
, J E. and Becker, D.L
a, I., Ikeda. M., Ma, K.W. and Mu-
eng,
.
204, 137-144.
in43 phosphorylation at S368 is acute
Establishment of steroidegenic granulosa cell li-
43
(2001) Intercellular communication via
3 gap junction messenger ribonucleic acid and
and differential express-
one receptor and the cell cycle modulate apoptosis
ular
tion.
onal coupling, ion fluxes and cell
ytosolic phosph-
l Oncology, 21(6),
004) Complex changes in cellular inositol
tion
cortical
propose that connexins phosphorylation-dependent mod-
ulation of gap junction coupling is a relevant mechanism
to regulate apoptosis of granulosa cells during the folic-
ular maturation.
6. ACKNOW
The authors thank Hans-Georg Hannibal and Frank Koepke for
lent technical support.
The project was partl
e DFG project Transregio 37/Q1.
Sabrina Schlie was supported by Enter-
and connexin 45 but absence of connexin 40 in granulosa
cell gap junctions of rat ovary. Journal Reprod Fertil,
107(2), 255-264.
[14] Ackert, C.L., Gittens, J.E.I., O’Brien, M.J., Eppig, J.J.
and Kidder, G.M.
ence and Quantum Applications.
REFERENCES
[1] Harris, A.L. (200
nels: Biophysics fills the gap. Quarterly Review of Bio-
physics, 34(3), 325-472.
[2] White, T.W. and Paul D.L. (1999) Genetic diseases and
gene knockouts reveal diverse connexin functions. Ann-
ual Review of Physiology, 61(1), 283-310.
[3] Willecke, K., Eiberger, J., Degen, J., Eckardt, J.D., Ro-
mualdi, A., Guldenagel, M., Deutsch, U. and Sohl, G.
(2002) Structural and functional diversity of connexin
genes in the mouse and human genome. The Journal of
Biological Chemistry, 383(5), 725-737.
[4] Nakase, T., Fushiki, S. and Naus, C.C. (
gap junctions composed of connexin 43 reduce apoptotic
neuronal damage in cerebral ischemia. Stroke, 34(8),
1987-1993.
[5] Doble, B.W.
B.E., Jin, Y., Cattini, P.A. and Kardami, E. (2003) Phos-
phorylation of serine 262 in the gap junction protein
connexin-43 regulates DNA synthesis in cell-cell contact
forming cardiomyocytes. Journal of Cell Science, 117(3),
507-514.
[6] Gorbe, A.
acs, L., Bagdi, E. and Krenacs, T. (2005) Transient upr-
egulation of connexin43 gap junctions and synchronized
cell cycle control precede myoblast fusion in regenerat-
ing skeletal muscle in vivo. Histochemistry and Cell Bi-
ology, 123(6), 573-583.
[7] Gorbe, A., Krenacs, T., Cook. oli
(2007) Myoblast proliferation and syncytial fusion both
depend on connexin43 function in transfected skeletal
muscle primary cultures. Experimental Cell Research,
313(6), 1135-1148.
[8] Zhang, Y.W., Morit
rota, S. (2001) Connexin43 suppresses proliferation of
osteosarcoma U2OS cells through post-transcriptional
regulation of p27. Oncogene, 20(31), 4138-4149.
[9] Zhang, Y.W., Chen, X., Wu, D., Liu, W., Wang, J., F
Z., Cai, G., Fu, B., Hong, Q. and Du, J. (2006) Downreg-
ulation of connexin 43 expression by high glucose indu-
ces senescence in glomerular mesangial cells. Journal of
the American Society of Nephrology, 17(6), 1532-1542.
[10] Ngezahayo, A., Altmann, B., Steffens, M. and Kolb, H
A. (2005) Gap junction coupling and apoptosis in GF-
SHR-17 granulosa cells. Journal of Membrane Biology
[11] Solan, J.L., Fry, M.D., TenBroek, E.M. and Lampe, P.D.
(2003) Connex
during S and G2/M and in response to protein kinase C
activation. Journal of Cell Science, 116(pt11), 2203-
2211.
[12] Keren, T.I., Dantes, A., Sprengel, R. and Amsterdam, A.
(1993)
nes expressing follicle stimulating hormone receptors.
Molecular and Cellular Endocrinology, 95, R1-R10.
[13] Okuma, A., Kuraoka, A., Iida, H., Inai, T., Wasano, K.
and Shibata, Y. (1996) Colocalization of connexin
connexin43 gap junctions is required for ovarian folicu-
logenesis in the mouse. Developmental Biology, 233(2),
248-270.
[15] Wiesen, J.F. and Midgley, R.A. (1994) Expression of
connexin4
protein during follicular atresia. Biology of Reproduction,
50(2), 336-348.
[16] Wright, C.S., Becker, D.L., Lin, J.S., Warner, A.E. and
Hardy, K. (2001) Stage-specific
ion of gap junctions in the mouse ovary: Connexin-spe-
cific roles in follicular regulation. Reproduction, 121, 77-
88.
[17] Quirk, M., Cowan, R.G. and Harman, R.M. (2004) Prog-
ester
in granulosa cells. Endocrinol, 145(11), 5033-5043.
[18] Sasson, R. and Amsterdam, A. (2003) Pleiotropic anti-
apoptotic activity of glucocorticoids in ovarian follic
cells. Biochemical Pharmacology, 66(8), 1393-401.
[19] Lampe, P.D. and Lau, A.F. (2004) The effects of con-
nexin phosphorylation on gap junctional communica
The International Journal of Biochemistry & Cell Biol-
ogy, 36(7), 1171-1186.
[20] Ngezahayo, A., Altmann, B. and Kolb, H.A. (2003)
Regulation of gap juncti
volume by cGMP in GFSHR-17 granulosa cells. Journal
of Membrane Biology, 194(3), 165-176.
[21] Van Rossum, G.S.A.T., Vlug, A.S., van den Bosch, H.,
Verkleij, A.J. and Boonstra, J. (2001) C
pase A2 activity during the ongoing cell cycle. Journal
of Cellular Physiology, 188, 321-328.
[22] Schumer, S.T. and Cannistra, S.A. (2003) Granulosa cell
tumor of the ovary. Journal of Clinica
1180-1189.
[23] Barker, C.J., Wright, J., Hughes, P.J., Kirk, C.J. and Mi-
chel, R.H. (2
phosphate complement accompany transit through the
cell cycle. Biochemical Journal, 380(pt 2), 465-473.
[24] Musil, L.S., Cunningham, B.A., Edelmann, G.M. and
Goodenough, D.A. (1990) Differential phosphoryla
of the gap junction protein connexin43 in junctional co-
mmunication-competent and deficient cell lines. The
Journal of Cell Biology, 111(5 pt 1), 2077-2088.
[25] Bittman, K.S. and LoTurco, J.J. (1999) Differential
regulation of connexin 26 and 43 in murine neo
precursors. Cerebral cortex, 9(2), 188-195.
[26] Solan, J.L. and Lampe, P.D. (2007) Key connexin43