 Vol.2, No.4, 244-255 (2012) Open Journal of Ecology http://dx.doi.org/10.4236/oje.2012.24028 The current state of the plant diversity in the Tlemcen region (Northwest Algeria) Smaïn El-Amine Henaoui*, Mohammed Bouazza Department of Ecology and Environment, Faculty of Natural Sciences and Life and Earth Sciences and the Universe, University of Tlemcen, Tlemcen, Algeria; *Corresponding Author: amine_33_ecoenv@yahoo.fr Received 28 July 2012; revised 11 September 2012; accepted 20 September 2012 ABSTRACT Tlemcen region is characterized by a Mediter- ranean climate, with a remarkable vegetal cover. The latter is influenced by the anthropozoo- logical action. For that, we realized a phytoeco- logical study based on the minimum area clas- sical method [1] which allow s us to have a good overview on plant biodiversity and better ana- lyze and interpret the vegetation at different levels (systematic composition; biological, mor- phological and biogeographical characteriza- tions; stratification; and plant species inventory which we hope comprehensive). The results show that our ecosystems are disrupted be- cause therophytisation, before last vegetation dynamics element [2], settles giving up a few feet of cork and holm oaks spe cies of ancient fo - rest. Keywords: Phytoecology; Plant Diversity; Minimum Area; Anthropic Action; Tlemcen (Western Algeria) 1. INTRODUCTION The nature and actual composition of Mediterranean plant communities cannot be understood without taking into account geological, paleoclimatic and anthropogenic factors that marked the evolution of various ecosystems specific to that biogeographical zone. The current Mediterranean region can be defined by evident floristic criteria since 50%, of some 25,000 spe- cies in the Mediterranean climatic area [3,4] and more so in the Mediterranean bioclimatic zone [5], are endemic [6]. Biodiversity includes three levels of biological vari- ability: ecosystem complexity, species richness and ge- netic variation [7]. Mediterranean plant biodiversity is the product, for many, of a traditional and harmonious environment use by man [8]. Despite the constant attacks they have suf- fered, for over a millennium, Mediterranean forests still offer, in some locations, a significant development. The problem posed by the dynamics of non-tree struc- tures (steppe, shrubland and lawns) can however lead to the understanding that their current expansion has been and is still under the human action impact, mainly through fire and grazing, but also the clearing [9]. In most North Mediterranean countries (France, Spain, etc.) these attacks are somehow controlled and give a biological rise, while in the Southern Mediterranean coun- tries (Algeria, Morocco, Tunisia, etc.) they continue in an uncontrolled manner. [10] emphasizes that “the analysis of the floristic rich- ness of different groups and their biological and chrono- logical characteristics allows to demonstrate their floral originality, conservation condition and, consquently, their heritage”. The flora of Algeria is characterized by a remarkable level of endemism (12.6% or 653 species listed on the 3139 including 7 endemic tree species [11]. However, Tlemcen region is not immune to Circum- Mediterranean natural laws. It has a very diverse flora and closely linked to various stressors. This vegetation has been largely studied; we can quote [12-16]. 2. LOCATION The study is located in the north-western part of Alge- ria. The study area is divided into two parts: one located in the mountains of Tlemcen and the other near the coast with rugged mountainous terrain (integral part of Traras Mountains). The stations choice mainly depends on the presence of pre-forest formations and shrubland. The area studied is located between 34˚ and 35˚25' North and 0˚55' and 2˚30' west and covers a 9017.69 km² area. It is limited geographically (Figure 1): ˗ To the north by the Mediterranean Sea, ˗ In the north-east by the wilaya of Aïn-Témouchent, ˗ To the east by the wilaya of Sidi-Bel-Abbès, ˗ To the west by Morocco. The stations of our study area are: Zarifet, Aïn-Fezza, Nedroma, Ghazaouet, Beni-Saf and Ouled-Mimoun (Fig- ure 1). Copyright © 2012 SciRes. OPEN A CCESS 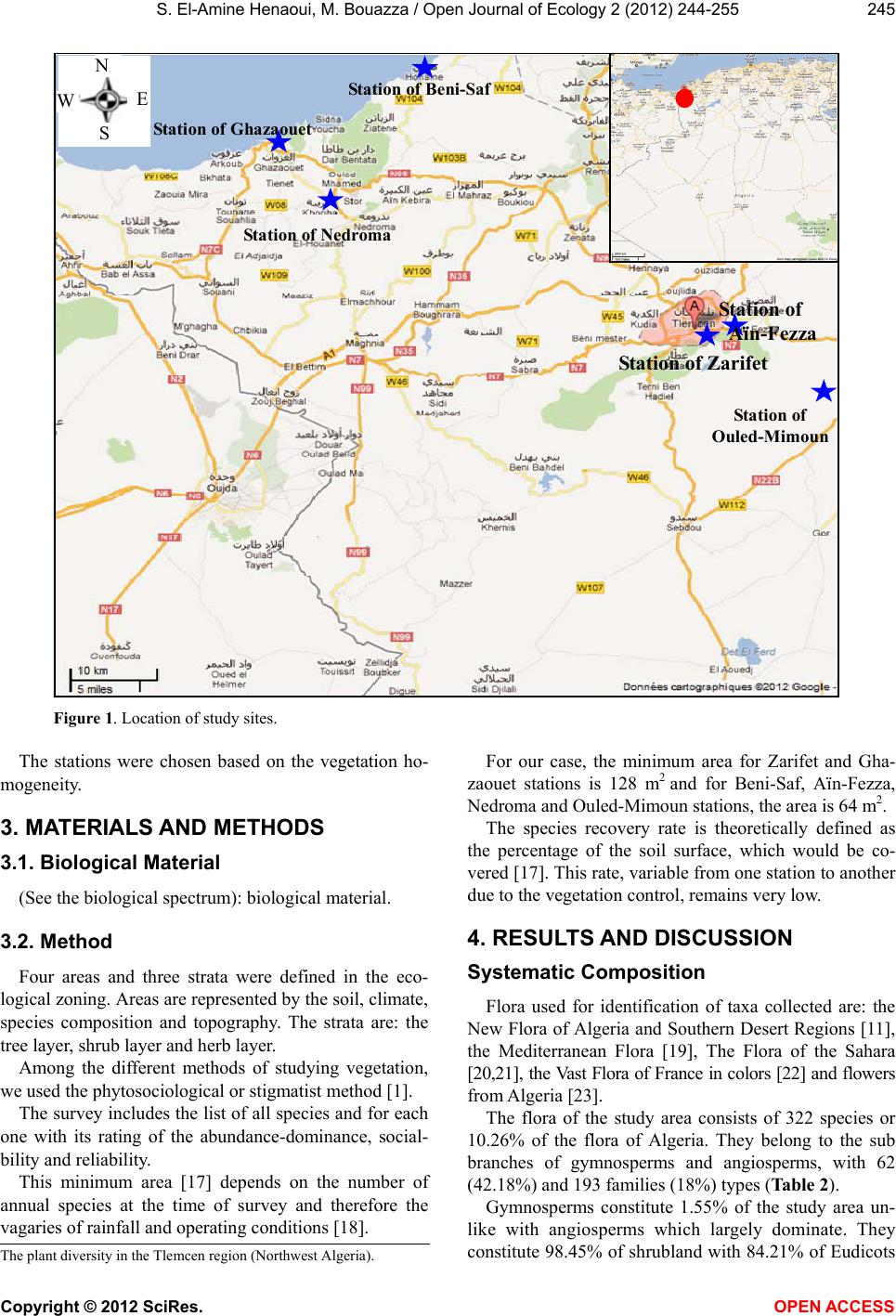 S. El-Amine Henaoui, M. Bouazza / Open Journal of Ecolo gy 2 (2012) 244-255 245 Station of Zarifet Station of Ghazaouet Sta t i o n of Be ni - S af Sta t i o n of Ne dr oma Station o f Aïn-Fezza St at io n o f Ouled-Mimoun W E S Figure 1. Location of study sites. The stations were chosen based on the vegetation ho- mogeneity. 3. MATERIALS AND METHODS 3.1. Biological Material (See the biological spectrum): biological material. 3.2. Method Four areas and three strata were defined in the eco- logical zoning. Areas are represented by the soil, climate, species composition and topography. The strata are: the tree layer, shrub layer and herb layer. Among the different methods of studying vegetation, we used the phytosociological or stigmatist method [1]. The survey includes the list of all species and for each one with its rating of the abundance-dominance, social- bility and reliability. This minimum area [17] depends on the number of annual species at the time of survey and therefore the vagaries of rainfall and operating conditions [18]. For our case, the minimum area for Zarifet and Gha- zaouet stations is 128 m2 and for Beni-Saf, Aïn-Fezza, Nedroma and Ouled-Mimoun stations, the area is 64 m2. The species recovery rate is theoretically defined as the percentage of the soil surface, which would be co- vered [17]. This rate, variable from one station to another due to the vegetation control, remains very low. 4. RESULTS AND DISCUSSION Systematic Composition Flora used for identification of taxa collected are: the New Flora of Algeria and Southern Desert Regions [11], the Mediterranean Flora [19], The Flora of the Sahara [20,21], the Vast Flora of France in colors [22] and flowers from Algeria [23]. The flora of the study area consists of 322 species or 10.26% of the flora of Algeria. They belong to the sub branches of gymnosperms and angiosperms, with 62 (42.18%) and 193 families (18%) types (Table 2). Gymnosperms constitute 1.55% of the study area un- like with angiosperms which largely dominate. They constitute 98.45% of shrubland with 84.21% of Eudicots The lant diversit in the Tlemcen re ion Northwest Al eria . Copyright © 2012 SciRes. OPEN A CCESS 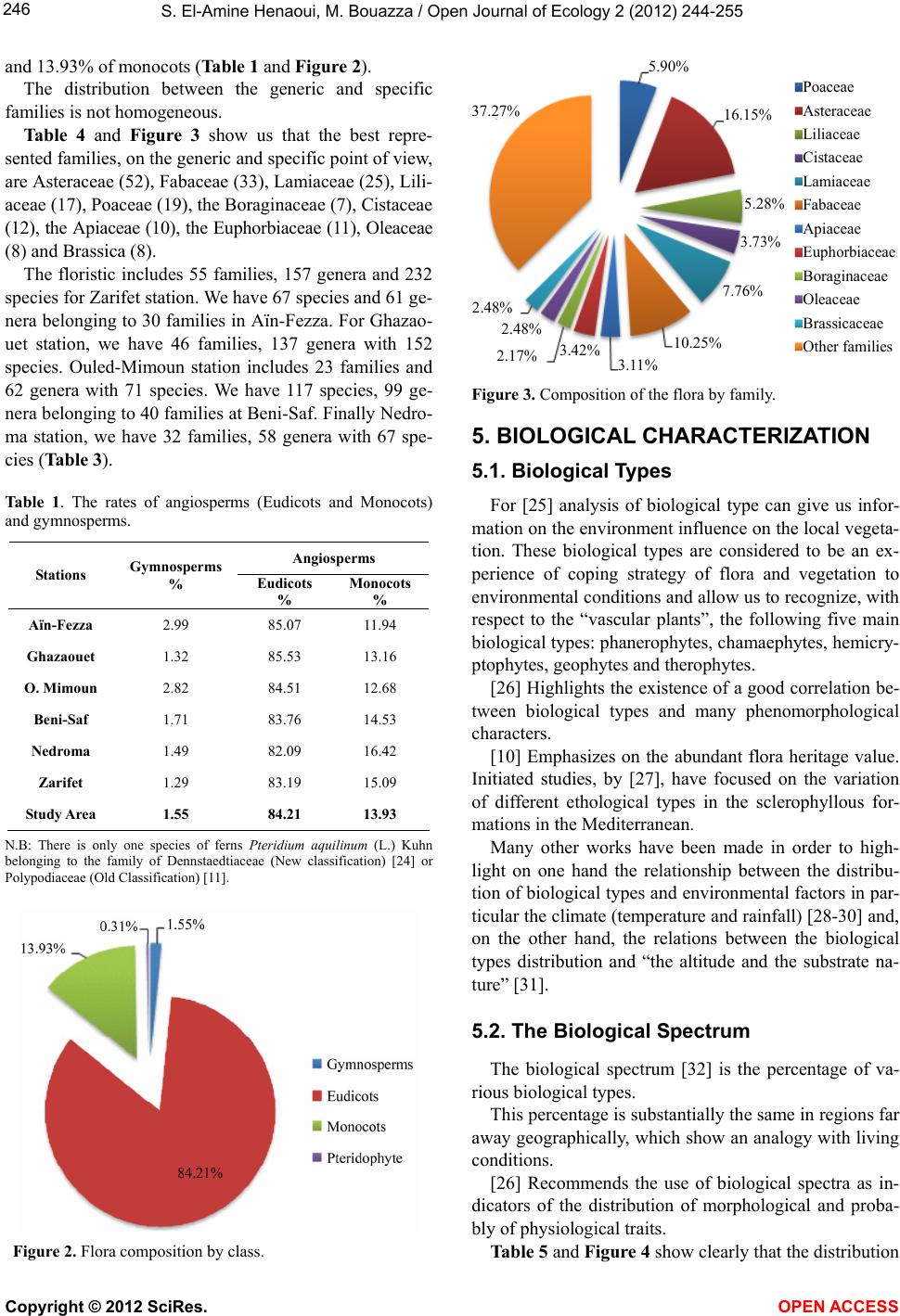 S. El-Amine Henaoui, M. Bouazza / Open Journal of Ecolo gy 2 (2012) 244-255 246 and 13.93% of monocots (Table 1 and Figure 2). The distribution between the generic and specific families is not homogeneous. Table 4 and Figure 3 show us that the best repre- sented families, on the generic and specific point of view, are Asteraceae (52), Fabaceae (33), Lamiaceae (25), Lili- aceae (17), Poaceae (19), the Boraginaceae (7), Cistaceae (12), the Apiaceae (10), the Euphorbiaceae (11), Oleaceae (8) and Brassica (8). The floristic includes 55 families, 157 genera and 232 species for Zarifet station. We have 67 species and 61 ge- nera belonging to 30 families in Aïn-Fezza. For Ghazao- uet station, we have 46 families, 137 genera with 152 species. Ouled-Mimoun station includes 23 families and 62 genera with 71 species. We have 117 species, 99 ge- nera belonging to 40 families at Beni-Saf. Finally Nedro- ma station, we have 32 families, 58 genera with 67 spe- cies (Table 3). Table 1. The rates of angiosperms (Eudicots and Monocots) and gymnosperms. Angiosperms Stations Gymnosperms % Eudicots % Monocots % Aïn-Fezza 2.99 85.07 11.94 Ghazaouet 1.32 85.53 13.16 O. Mimoun 2.82 84.51 12.68 Beni-Saf 1.71 83.76 14.53 Nedroma 1.49 82.09 16.42 Zarifet 1.29 83.19 15.09 S tudy Area 1.55 84.21 13.93 N.B: There is only one species of ferns Pteridium aquilinum (L.) Kuhn belonging to the family of Dennstaedtiaceae (New classification) [24] or Polypodiaceae (Old Classification) [11]. Figure 2. Flora composition by class. Poaceae Asteraceae Liliaceae Cistaceae Lamiaceae Fabaceae Apiaceae Euphorbiaceae Boraginaceae Oleaceae Brassicaceae Other families 37.27% 5.90% 16.15% 5.28% 3.73% 7.76% 10.25% 3.11% 3.42% 2.17% 2.48% 2.48% Figure 3. Composition of the flora by family. 5. BIOLOGICAL CHARACTERIZATION 5.1. Biological Types For [25] analysis of biological type can give us infor- mation on the environment influence on the local vegeta- tion. These biological types are considered to be an ex- perience of coping strategy of flora and vegetation to environmental conditions and allow us to recognize, with respect to the “vascular plants”, the following five main biological types: phanerophytes, chamaephytes, hemicry- ptophytes, geophytes and therophytes. [26] Highlights the existence of a good correlation be- tween biological types and many phenomorphological characters. [10] Emphasizes on the abundant flora heritage value. Initiated studies, by [27], have focused on the variation of different ethological types in the sclerophyllous for- mations in the Mediterranean. Many other works have been made in order to high- light on one hand the relationship between the distribu- tion of biological types and environmental factors in par- ticular the climate (temperature and rainfall) [28-30] and, on the other hand, the relations between the biological types distribution and “the altitude and the substrate na- ture” [31]. 5.2. The Biological Spectrum The biological spectrum [32] is the percentage of va- rious biological types. This percentage is substantially the same in regions far away geographically, which show an analogy with living conditions. [26] Recommends the use of biological spectra as in- dicators of the distribution of morphological and proba- bly of physiological traits. Table 5 and Figure 4 show clearly that the distribution Copyright © 2012 SciRes. OPEN A CCESS  S. El-Amine Henaoui, M. Bouazza / Open Journal of Ecolo gy 2 (2012) 244-255 Copyright © 2012 SciRes. OPEN A CCESS 247 Table 2. Composition in families, genera and species of flora. Families Genera Species Asteraceae Crassulaceae 33 1 51 3 Fabaceae Violaceae 20 1 30 2 Lamiaceae Resedaceae 15 1 25 2 Poaceae Solanaceae 13 2 18 2 Liliaceae Orchidaceae 10 2 17 2 Cistaceae Gentianaceae 5 2 12 2 Euphorbiaceae Aristolochiaceae 3 1 10 2 Apiaceae Thymeleaceae 7 1 9 2 Brassicaceae Pinaceae 6 1 7 2 Oleaceae Araceae 3 1 6 1 Renonculaceae Acanthaceae 4 1 7 1 Boraginaceae Apocynaceae 5 1 7 1 Caryophyllaceae Palmaceae 4 1 7 1 Rosaceae Buxaceae 4 1 5 1 Fagaceae Frankeniaceae 1 1 5 1 Scrofulariaceae Zygophyllaceae 4 1 4 1 Plantaginaceae Fumariaceae 1 1 5 1 Primulaceae Globulariaceae 1 1 4 1 Cupressaceae Juncaceae 2 1 3 1 Iridaceae Cesalpinieae 1 1 4 1 Linaceae Chenopodiaceae 1 1 4 1 Ericaceae Plumbaginaceae 2 1 3 1 Malvaceae Oxalidaceae 2 1 2 1 Dipsaceae Orobanchaceae 2 1 3 1 Convolvulaceae Polypodiaceae 1 1 3 1 Geraniaceae Asclepiadaceae 1 1 2 1 Rubiaceae Rutaceae 3 1 3 1 Caprifoliaceae Campanulaceae 2 1 3 1 Terebinthaceae Myrtaceae 1 1 2 1 Rhamnaceae Salicaceae 2 1 3 1 Valerianaceae Papaveraceae 1 1 1 1  S. El-Amine Henaoui, M. Bouazza / Open Journal of Ecolo gy 2 (2012) 244-255 248 Table 3. Inventory of families as a percentage (Study sites). Families Aïn-Fezza (%) Ghazaouet (%) O. Mimoun (%) Beni-Saf (%) Nedroma (%) Zarifet (%) Poaceae 4.48 6.58 8.45 9.40 4.48 5.17 Asteraceae 17.91 15.79 19.72 20.51 17.91 14.66 Liliaceae 4.48 4.61 2.82 3.42 8.96 6.47 Cistaceae 2.99 5.26 4.23 4.27 1.49 4.74 Lamiaceae 10.45 11.18 9.86 8.55 4.48 7.33 Fabaceae 5.97 7.24 18.31 7.69 14.93 9.48 Apiaceae 2.99 5.26 1.41 4.27 1.49 3.88 Euphorbiaceae - 1.97 2.82 3.42 1.49 2.59 Boraginaceae 2.99 2.63 - 0.85 2.99 1.72 Oleaceae 1.49 2.63 2.82 0.85 2.99 1.72 Brassicaceae 5.97 2.63 5.63 1.71 2.99 2.59 Other families 40.30 34.21 23.94 35.04 35.82 39.66 Table 4. Floristic composition per family (Study area). Family Number of species % Poaceae 19 5.90 Asteraceae 52 16.15 Liliaceae 17 5.28 Cistaceae 12 3.73 Lamiaceae 25 7.76 Fabaceae 33 10.25 Apiaceae 10 3.11 Euphorbiaceae 11 3.42 Boraginaceae 7 2.17 Oleaceae 8 2.48 Brassicaceae 8 2.48 Other families 120 37.27 of biological types in the vegetation in the same station and between stations is heterogeneous: ˗ Aïn-Fezza and Ghazaouet stations follow the pattern: Th > Ch > Ph > He > Ge ˗ Ouled-Mimoun station follows the pattern: Th > Ph > CH > He > Ge ˗ Beni-Saf station follows the pattern: Th > Ch > He > Ph > Ge ˗ Nedroma station follows the pattern: Th > Ch > He and Ph > Ge ˗ Zarifet station follows the pattern: Th > Ch > Ph > He and Ge ˗ The study area follows the pattern: Th > Ch > Ph > He > Ge For all the stations studied, therophytes have the high- est rate (39.41%), reflecting the high population pres- sure. In addition to human impacts, the therophytisation tra- ces its beginnings to the phenomenon of desiccation [12,15,33-36]. The latter present therophyty as a form of resistance to drought, and to drylands high tempera- tures. Despite the therophytes importance, the chamaephytes keep a particularly important place. For Zarifet and Ghazaouet stations, the rate of phan- erophytes is low, despite a semblance of sylvatic envi- ronment. This is mainly due to the floristic richness of these stations, and the recent invasion by therophytes. For Beni-Saf station, the rate of phanerophytes remains very low; the results clearly show its degraded state. It is dominated by therophytes, chamaephytes, hemicrypto- phytes, phanerophytes and geophytes. Some authors like [37] explain the abundance of hemi- cryptophytes, in North Africa, a greater richness in or- ganic matter in forest and elevation. This explains the low percentage, of hemicryptophytes in Ghazaouet station, Copyright © 2012 SciRes. OPEN A CCESS 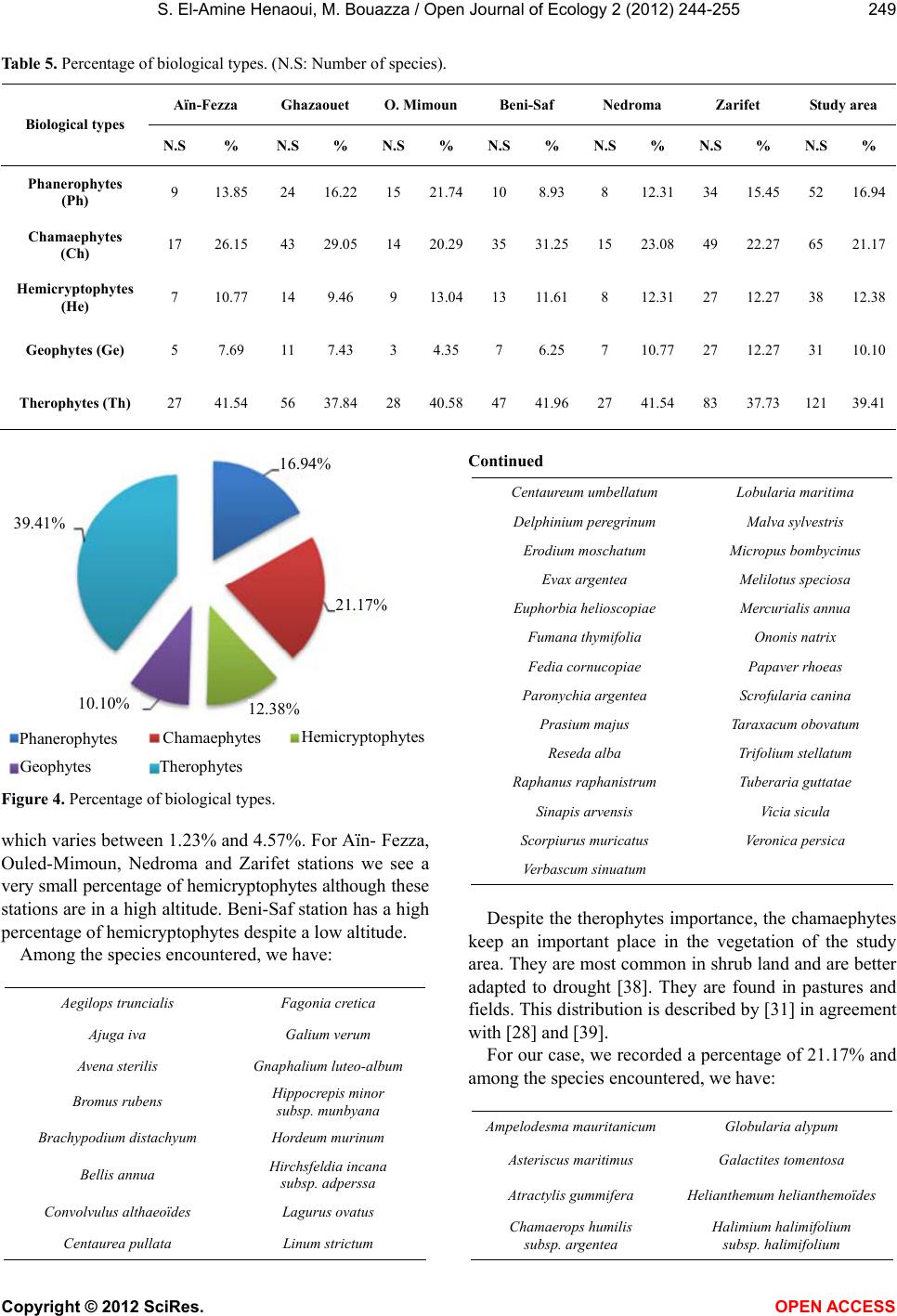 S. El-Amine Henaoui, M. Bouazza / Open Journal of Ecolo gy 2 (2012) 244-255 249 Table 5. Percentage of biological types. (N.S: Number of species). Aïn-Fezza Ghazaouet O. Mimoun Beni-Saf Nedroma Zarifet Study area Biological types N.S % N.S % N.S% N.S% N.S% N.S % N.S% Phanerophytes (Ph) 9 13.85 24 16.22 15 21.7410 8.938 12.31 34 15.45 52 16.94 Chamaephytes (Ch) 17 26.15 43 29.0514 20.29 35 31.2515 23.0849 22.27 65 21.17 Hemicryptophytes (He) 7 10.77 14 9.469 13.0413 11.61 8 12.3127 12.27 38 12.38 Geophytes (Ge) 5 7.69 11 7.433 4.35 7 6.25 7 10.7727 12.27 31 10.10 Therophytes (Th) 27 41.54 56 37.8428 40.58 47 41.9627 41.5483 37.73 12139.41 Hemicryptophytes Therophytes Chamaephytes Geophytes Phanerophytes 10.10% 21.17% 12.38% 16.94% 39.41% Figure 4. Percentage of biological types. which varies between 1.23% and 4.57%. For Aïn- Fezza, Ouled-Mimoun, Nedroma and Zarifet stations we see a very small percentage of hemicryptophytes although these stations are in a high altitude. Beni-Saf station has a high percentage of hemicryptophytes despite a low altitude. Among the species encountered, we have: Aegilops truncialis Fagonia cretica Ajuga iva Galium verum Avena sterilis Gnaphalium luteo-a lbum Bromus rubens Hippocrepis minor subsp. munbyana Brachypodium distachyum Hordeum murinum Bellis annua Hirchsfeldia incana subsp. adperssa Convolvulus althaeoïdes Lagurus ovatus Centaurea pullata Linum strictum Continued Centaureum umbellatum Lobularia maritima Delphinium peregrinum Malva sylvestris Erodium moschatum Micropus bombycinus Evax argentea Melilotus speciosa Euphorbia heli oscopiae Mercurialis annua Fumana thymifolia Ononis natrix Fedia cornucopiae Papaver rh oeas Paronychia argentea Scrofularia can i na Prasium majus Taraxacum obovatum Reseda alba Trifolium stellatum Raphanus raph a n istrum Tuberar ia guttatae Sinapis arvensis Vicia sicula Scorpiurus muricatus Veronica persica Verbascum sinuatum Despite the therophytes importance, the chamaephytes keep an important place in the vegetation of the study area. They are most common in shrub land and are better adapted to drought [38]. They are found in pastures and fields. This distribution is described by [31] in agreement with [28] and [39]. For our case, we recorded a percentage of 21.17% and among the species encountered, we have: Ampelodesma mauritanicumGlobularia alypum Asteriscus maritimus Galactites tomentosa Atractylis gummifera Helianthemum helianthemoïdes Chamaerops humilis subsp. argentea Halimium halimifolium subsp. halimifolium Copyright © 2012 SciRes. OPEN A CCESS  S. El-Amine Henaoui, M. Bouazza / Open Journal of Ecolo gy 2 (2012) 244-255 250 Continued Cistus monspeliensis Helichrysum stoechas Calycotome intermedia Inula viscosa Cistus salviifolius Jasminum fruticans Daphne gnidium Juncus maritimus Daucus carota Kundmannia sicu la Eryngium tricuspidatum Lavandula dentata Echinops spinosus Lavandula stoechas Erica arborea Lavandula multifida Erica multiflora Nepeta multibracteata Ferula communis Pallenis spinosa Frankenia laevis Psoralea bituminosa Phlomis bovei Thapsia garganica Reichardia picroïdes Ulex boivinii Ruta chalepensis Ulex parvif lo ru s Sideritis montana Viola arborescens Sedum acre Viola odorata Senecio vulgaris Globularia alypum Teucrium fruticans Galactites tomentosa Fumaria capreolata Helianthemum helianthemoïdes Halimium halimifolium subsp. halimifolium Thymus ciliatus subsp. coloratus Hemicryptophytes, with a percentage of 12.38%, are still poorly represented in the study area we find: Asperula hirsuta Dianthus serrulatus Atractylis humilis Echium vulgare Anchusa azurea Echium parviflorum Andropogon hirtus Echium italicum Bellis sylvestris Geranium sylvaticum Centaurea soltitialis Geranium robertianum subsp. purpureum Convolvulus tricolor Inula montana Carduus pycno-cephalus Leontondon hispidulus Dactylis glomerata Marrubium vulgare Plantago lagopus Sonchus arvens i s Plantago serraria Salvia verbenaca Ranunculus repens Solenanthus lanatus Ranunculus bulbatus Tr a g opogon porrifolius Rubia peregrina They seem to increase in forest and high altitude. The dominance of hemicryptophytes is then an obstacle to the installation of phanerophytes. Despite the low participation of phanerophytes species; they are dominant by their biomass, thus constitute the bushes and forests. They are abundant in the vegetation of Ghazaouet, Ouled Mimoun and Zarifet stations, which still bears witness to the existence of a forest and/or a pre-forest formation. These are usually Quercetea ilicis species: Arbutus unedo Myrtus communis Buxus sempervirens N icot ian a g lauca Cupressus sempervirens Olea europea Cistus ladaniferus subsp. africanus Pinus halepensis Clematis flammula Populus alba Crataegus oxyacantha Pistacia lentiscus Ceratonia siliqua Pistacia terebinthus Colutea arborescens Phillyrea angustifolia Fraxinus ornus Phillyrea angustifolia subsp. eu-angustifolia Genista tricuspidata Phillyrea angustifolia subsp. media Juniperus oxycedrus subsp. oxycedrus Phillyrea angustifolia subsp. latifolia Juniperus phoenicea Pinus maritima Lonicera implexa Periploca laevigata Lonicera biflora Rhamnus lycioïdes Ligustrum japonicum Rhamnus alaternus Rhus pentaphylla Viburnum tinus Rosmarinus officinalis Withania frutescens Rosa sempervirens Ziziphus lotus Rosa canina Quercus ilex Robinia pseudo-acacia Quercus coccifera Rubus ulmifolius Quercus suber Ricinus communis Quercus faginea subsp. tlemceniensis Tetraclinis articulata We note that the absence of nanophanerophytes is cer- tainly linked to severe erosion in the region but, espe- cially, the power of the winds [40]. Finally, geophytes are everywhere; the least repre- Copyright © 2012 SciRes. OPEN A CCESS  S. El-Amine Henaoui, M. Bouazza / Open Journal of Ecolo gy 2 (2012) 244-255 251 sented with only 10.10%. Representation of the latter remains higher in the shrub land (10.77% in Nedroma) than in the pre-forest formations and forest (7.43% in Ghazaouet). Among this group we have the Liliacaea, the Iridaceae, the Orchidaceae and Araceae represented by the follow- ing species: Allium roseum Agropyron repens Asphodelus microcarpus Ballota hirsuta Aristolochia longa Bellevallia dubia Aristolochia baetica Gladiolus segetum Asparagus stipularis Iris planifolia Asparagus acutifolius Iris unguicularis Asparagus albus Iris tingitana Asparagus officinalis Muscari neglectum Arisarum vulgare Muscari comosum Ornithogalum umbellatum Stipa tortilis Ophrys tenthredinifera subsp. lutescens Stipa tenacissima Orchis italica Tulipa sylvestris Smilax aspera subsp. altissima Urginea maritima Scilla lingulata [30] Also found higher geophytes proportions in Medi- terranean area than in steppe domain. The origin of the extension of therophytes is due: ˗ Either to the adaptation to the winter cold stress [20,28] or to summer drought [29,34]. ˗ Or even to the medium disturbances by grazing, crops, etc. [41]. Indeed, plant formations in dominant phanerophytic vegetal cover have the lowest recovery of therophytes while those, for which the phanerophytes rate is negligi- ble, therophytes recovery is much higher. Similarly, [42] explain the changes of floristic composition and species biological cycle (after cutting the undergrowth of ever- green oak), by the increasing of the lower layers bright- ness level. 5.3. Disruption Index The disturbance index (calculated according to [43]) quantifies the therophytisation of a medium. IP = Chamaephytes number + Therophytes number/Species total number For all stations this index remains high compared to [44] results in Tunisia, where he obtained 70% as high value. The chamaephytes and therophytes are the bio- logical types that dominate in the region. This shows the anthropozoologic high pressure experienced by vegeta- tion in the study area. The disturbance index being about 57% for the entire study area, this shows clearly the sharp decline (Table 6). 6. MORPHOLOGICAL CHARACTERIZATION The sharp deterioration affects species regeneration. The non-regenerating perennial causes changes that give non-resilient paths, and also causes a change in potential output and the botanical composition [45]. The vegetal cover is dominated by three vegetation types: woody per- ennials, herbaceous perennial and annual grasses. The morphological point of view, the vegetation of the study area is marked by heterogeneity between woody and herbaceous species and between perennials and an- nuals. The annual herbaceous species are dominant with a percentage of 50.80%. The herbaceous perennials come in second with 25.72%. In third place we find the woody perennial with a percentage of 23.47% (Table 7) and (Figure 5). The intense human impact, which the forests of the re- gion continue to suffer, results in the invasion of thero- phytes, which are typically annual herbs. Thus, the harsh conditions favor the development of short-cycle herba- ceous species at the expense of woody perennials gener- ally more demanding in water and nutritional needs. 7. STRATIFICATION At the study area we have the herbaceous layer, which dominates with a percentage of 75.45%, then the shrub layer with 14.67% and finally the tree layer with a small percentage that is of the order of 9.88% (Table 8). The herbaceous layer is generally formed by thero- phytes which are annual grasses in general (Figure 6). 8. BIOGEOGRAPHIC CHARACTERIZATION The study of the Mediterranean basin flora is of great Tabl e 6. Index of stations disruption studies/study area. (A.F: Aïn-Fezza, GZT: Ghazaouet, O.M: Ouled. Mimoun, B.S: Beni- Saf, NDR: Nedroma and ZRT: Zarifet). StationsA.FGZTO.M B.S NDR ZRTThe study area IP 67%66%60% 73% 64% 60%57% Copyright © 2012 SciRes. OPEN A CCESS 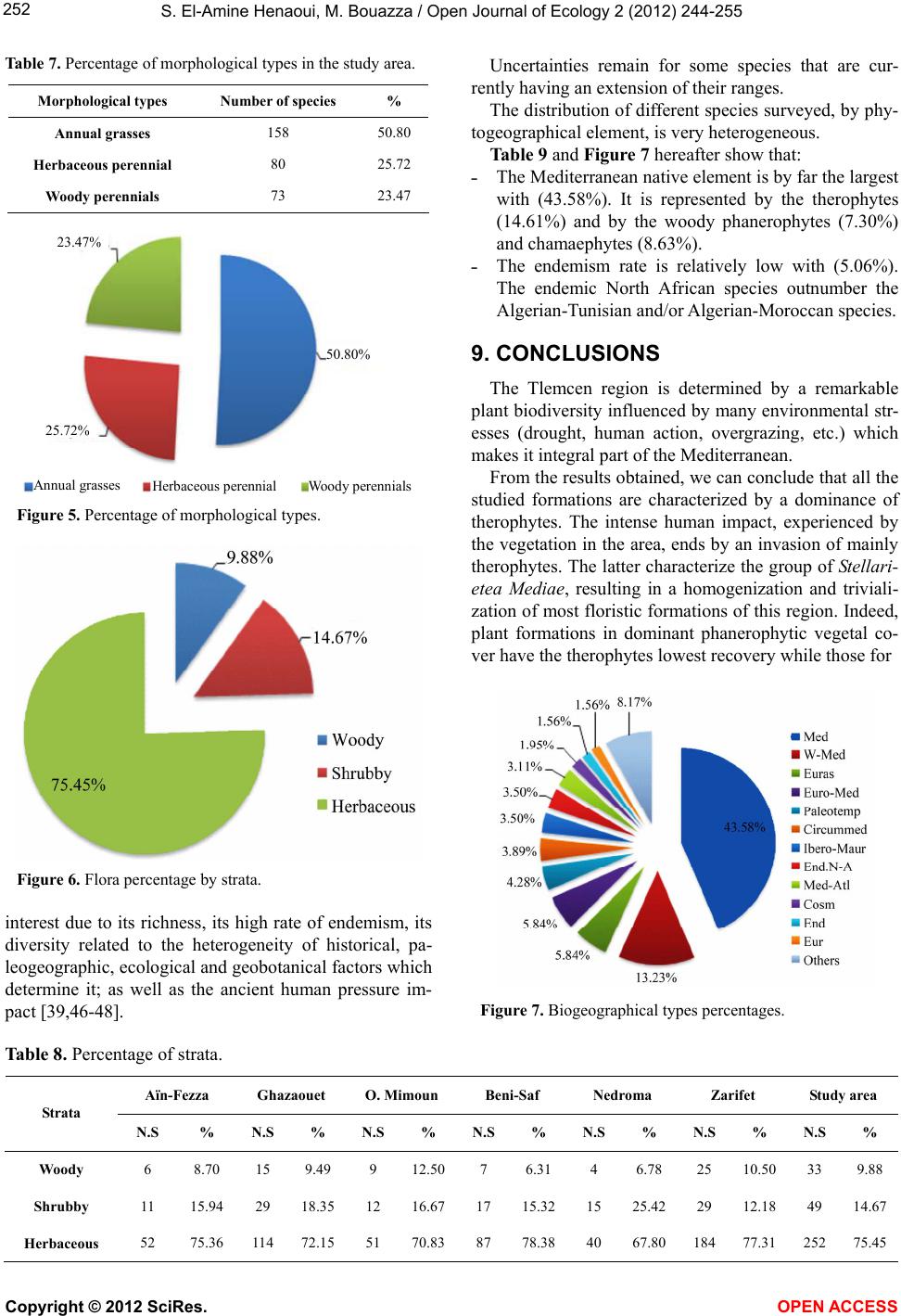 S. El-Amine Henaoui, M. Bouazza / Open Journal of Ecolo gy 2 (2012) 244-255 Copyright © 2012 SciRes. OPEN A CCESS 252 Table 7. Percentage of morphological types in the study area. Morphological types Number of species % Annual grasses 158 50.80 Herbaceous per ennial 80 25.72 Woody perennials 73 23.47 Uncertainties remain for some species that are cur- rently having an extension of their ranges. The distribution of different species surveyed, by phy- togeographical element, is very heterogeneous. Table 9 and Figure 7 hereafter show that: ˗ The Mediterranean native element is by far the largest with (43.58%). It is represented by the therophytes (14.61%) and by the woody phanerophytes (7.30%) and chamaephytes (8.63%). Woody perennials 50.80% 23.47% Herbaceous perennial Annual grasses 25.72% ˗ The endemism rate is relatively low with (5.06%). The endemic North African species outnumber the Algerian-Tunisian and/or Algerian-Moroccan species. 9. CONCLUSIONS The Tlemcen region is determined by a remarkable plant biodiversity influenced by many environmental str- esses (drought, human action, overgrazing, etc.) which makes it integral part of the Mediterranean. From the results obtained, we can conclude that all the studied formations are characterized by a dominance of therophytes. The intense human impact, experienced by the vegetation in the area, ends by an invasion of mainly therophytes. The latter characterize the group of Stellari- etea Mediae, resulting in a homogenization and triviali- zation of most floristic formations of this region. Indeed, plant formations in dominant phanerophytic vegetal co- ver have the therophytes lowest recovery while those for Figure 5. Percentage of morphological types. Figure 6. Flora percentage by strata. interest due to its richness, its high rate of endemism, its diversity related to the heterogeneity of historical, pa- leogeographic, ecological and geobotanical factors which determine it; as well as the ancient human pressure im- pact [39,46-48]. Figure 7. Biogeographical types percentages. Table 8. Percentage of strata. Aïn-Fezza Ghazaouet O. Mimoun Beni-Saf Nedroma Zarifet Study area St rata N.S % N.S % N.S % N.S% N.S % N.S % N.S % Woody 6 8.70 15 9.49 9 12.507 6.31 4 6.78 25 10.50 33 9.88 Shrubby 11 15.94 29 18.35 12 16.67 17 15.32 15 25.4229 12.18 49 14.67 Herbaceous 52 75.36 114 72.15 51 70.8387 78.38 40 67.80184 77.31 252 75.45  S. El-Amine Henaoui, M. Bouazza / Open Journal of Ecolo gy 2 (2012) 244-255 253 Table 9. Biogeographic types with species number and percentage. Biogeographic types Signification Number % Med Mediterranean 112 43.58 W-Med W-Mediterranean 34 13.23 Euras Eurasian 15 5.84 Euro-Med European-Mediterranean 15 5.84 Paleotemp Paleotempered 11 4.28 Circummed Circum-mediterranean 10 3.89 Ibero-Maur Ibero-Mauritanean 9 3.50 End. N-A Endemic. North-African 9 3.50 Med-Atl Mediterranean-Atlantic 8 3.11 Cosm Cosmopolitan 5 1.95 Subcosm Subcosmopolitan 5 1.95 Macar-Med Macaronesian-Mediterranean 4 1.56 End Endemic 4 1.56 Eur European 4 1.56 Paleosubtrop Paleosubtropical 2 0.78 Circumbor Circumboreal 2 0.78 Ibero-Mar Ibero-Morrocan 2 0.78 Euras-Med Eurasian-Mediterranean 2 0.78 N-A North-African 2 0.78 Sah Saharian 1 0.39 Sah-Sind Sahara-Sindian 1 0.39 which the rate of phanerophytes is negligible, the thero- phytes recovery is much higher. The disturbance index being about 57% for the entire study area, this shows clearly the sharp deterioration. From the morphological point of view, the study area vegetal formations are marked by heterogeneity between woody and herbaceous species and between perennials and annuals. The annual herbaceous species are domin- ant with a percentage of 50.80%. The herbaceous peren- nials come in second with 25.72%. In third place we find the woody perennials with 23.47%. From the phytogeographical point of view, the Medi- terranean element is the most dominant with 43.58%. Finally, conservation of natural heritage remains a ma- jor initiative to preserve the region plant biodiversity against a regression which may become irreversible in the near future. In this regard, Algeria, as all Mediterranean countries, has long been involved in the preservation and conservation of biodiversity policy through the creation of several national parks. Currently, Algeria has eight national parks that in- clude all original landscapes and where are the main hot spots of plant biodiversity in the country [49]. Meanwhile, several research works, mainly focused on the inventory and mapping of this phytobiodiversity, have been done in these hot spot. Among these works, we mention those of [50] in Chrea National Park, [51] and [52] in El-Kala national park, [53] and [54] in Tlem- cen National Park. All these works have stressed the im- portance of such an inventory in the sound management of natural ecosystems. Indeed, several authors suggested that the conservation and enhancement of a natural eco- system requires a good knowledge of its biodiversity [49,55]. Many of these works in these ecosystems have emphasized the rich flora of these environments and have shown a variety of endemic and/or rare species to be placed in conservation priorities. They also discussed the degradation advanced state of these natural ecosys- tems which is mainly attributed to the combined action of man and his animals (grazing). REFERENCES [1] Braun-Blanquet, J. (1951) The plant communities of Me- diterranean France. C.N.R.S., Paris. [2] Quezel, P. (2000) Reflections on the evolution of the flora Copyright © 2012 SciRes. OPEN A CCESS  S. El-Amine Henaoui, M. Bouazza / Open Journal of Ecolo gy 2 (2012) 244-255 254 and vegetation in Mediterranean Maghreb. Ibis Press, Paris. [3] Emberger, L. (1930) Climate on a formula applicable in botanical geography. Comptes Rendus de l’Académie des Sciences, 1991, 389-390. [4] Emberger, L. (1930b) The vegetation of the Mediterra- nean region. Test of a classification of plant communities. The Scientific Journal Revue Générale de Botanique, 42, 341-404, 641-662. [5] Daget, Ph. (1977) The Mediterranean bioclimate, general characteristics, classification schemes. Vegetatio, 34, 1- 20. doi:10.1007/BF00119883 [6] Quezel, P. (1985) Definition of the Mediterranean region and the origin of its flora. Gomez-Campo Edit—“Plant conservation in the Mediterranean area” Junk, Dordrecht, 9-24. [7] Robert-Pichette, P. and Gillespie, L. (2000) Monitoring pro- tocols for terrestrial plant biodiversity. Lexicon, Director- ate for Science Ecosystems, Environment Canada, Web Site. [8] Quezel, P. (1999) Plant biodiversity of Mediterranean forests its possible evolution by thirty years. Mediterra- nean Forest, 20, 3-8. [9] Quezel, P. (1999) Large structures in the Mediterranean vegetation: Determining factors in their implementation postglacial. Geobios, 32, 19-32. doi:10.1016/S0016-6995(99)80081-3 [10] Dahmani-Megrerouche, M. (1997) The oak in Algeria. Syn- taxonomy, phytosociology and stand dynamics. Ph.D. Thesis, University of Science and Technology, Houari Boumediene, Algiers. [11] Quezel, P. and Santa S. (1962-1963) New flora of Algeria and the desert regions Meridional. French National Cen- ter for Scientific Research, Paris. [12] Benabadji, N., Bouazza, M., Merzouk, A. and Ghezlaoui, S.M. (2004) Appearance of phytoecological atriplexaies north of Tlemcen (Orany-Algeria). Science and Techno- logy, 22, 62-79. [13] Benabadji, N., Benmansour, D. and Bouazza, M. (2007) The flora of the mountains of Ain Fezza in western Alge- ria, and biodiversity dynamics. Science and Technologies, 26, 47-59. [14] Bouazza, M., Benabadji, N. and Loisel R. (2001) Balance of flora in the region of Tlemcen (Orany-Algeria). Medi- terranean Forest, 22, 130-136. [15] Bouazza, M., Benabadji, N., Loisel, R. and Metge, G. (2004) Evolution of the steppe vegetation in the south- west of Orany (Algeria). Critical Reviews in Oral Biology and Medicine, 30, 221-231. [16] Mesli-Bestaoui, K. (2009) Contribution to an ecological survey and vegetation dynamics of the mounts of Tlem- cen by a mapping approach. Thesis, Abou-Bakr Belkaïd University of Tlemcen, Tlemcen. [17] Guinochet, M. (1973) Phytosociology. Masson Edit, Paris. [18] Djebaïli, S. (1984) Algerian steppe, Ecology and phyto- sociology. Odessa Polytechnic National State University, Algiers. [19] Paccalet, Y. (1981) Mediterranean flora. Hatier, Paris. [20] Ozenda, P. (1963) Organization and reproduction of an- giosperms. In: Abbayes, et al., Eds., Botany: Anato my, Life Cycles, Systematics, Masson and Cie Eds, Paris, 645-722. [21] Ozenda, P. (1977) Flora of the Sahara. 2nd Edition, French National Center for Scientific Research, Paris. [22] Bonnier, G. (1990) The vast flora colors Gaston Bonnier. France, Switzerland, Belgium and neighboring countries, Editions Belin, Paris. [23] Beniston, N.T. and Beniston, W.S. (1984) Flowers from Algeria. National Book Company, Algiers. [24] The International Plant Names Index (IPNI). http://www.uk.ipni.org/ [25] Raunkiaer, C. (1907) The life forms of plants and their bearing on geography. In: Raunkiaer, C., Ed., The Life Forms of Plants and Statistical Plant Geography, 1934. (Translated from Danish, with Introduction by A. G. Tan- sely), Oxford University Press, Oxford, 2-104. [26] Romane, F. (1987) Distribution efficiency of plant growth forms to anlyse vegetation at the regional scale. Case of some green oak coppice Languedoc. Ph.D. Thesis, State University, Aix-Marseille. [27] Barbero, M. and Quezel, P. (1989b) Contribution to the study of phytosociological matorrals Eastern Mediter- ranean. Lazoco, 11, 37-56. [28] Raunkiaer, C. (1934) The life forms of plants and statis- tical plant. Geography Claredon Press, Oxford. [29] Daget, Ph. (1980) On life forms as adaptive strategy. (Case therophytes). In: Barbault, R., Blandin, P. and Me- yer, J.A., Eds., Searches of Theoretical Ecology, Adaptive Strategies, Maloines, Paris, 89-114. [30] Danin, A. and Orshan, G. (1990) The distribution of Raun- kiaer life forms in Israël in relation to the environment. Journal of Vegetation Science, 1, 41-48. doi:10.2307/3236051 [31] Floret, C., Galan, M.J., Lefloch, E., Orshan, G. and Ro- mane F. (1990) Growth forms and phenomorphology traits along an environnemental gradient: Tools for studing vege- tation. Journal of Vegetation Science, 1, 71-80. doi:10.2307/3236055 [32] Gaussen, H., Leroy, J.F. and Ozenda P. (1982) Accurate botanical 2: Higher plants. Edit Masson, Paris. [33] Sauvage, Ch. (1961) Geobotanical research on cork oak in Morocco. Thesis, State, Montpellier, Sharifian, Bo- tanical Series, 21-462. [34] Negre, R. (1966) Therophytes. Memory, Society, Botany, F1, 92-108. [35] Daget, Ph. (1980) A current element of the characteriza- tion of the Mediterranean world: Climate. Naturalia Mon- speliensia, 1, 101-126. [36] Aïdoud, A. (1983) Contribution to the study of steppe ecosystems of South Orany: Phytomass, primary produc- tivity and pastoral applications. Thesis, Third Cycle, Uni- versity of Science & Technology, H. Boumediene, Algiers. [37] Barbero, M. and Quezel, P. (1989) Structures, architec- tures and sclerophyllous forest fire prevention. Bulletin Ecology, 20, 7-14. Copyright © 2012 SciRes. OPEN A CCESS  S. El-Amine Henaoui, M. Bouazza / Open Journal of Ecolo gy 2 (2012) 244-255 Copyright © 2012 SciRes. OPEN A CCESS 255 [38] Ellenberg, H., Mueller, A. and Dombois, D. (1968) Akey to Raunkiaer plant fee forms with revised. Subdivision, Berlin, Geobotanic, Institute, ETH, Stiftg. Rübel, Zürich, 37, 56-73. [39] Orshan, G. (1984) Monocharacter growth form types as a tool in an analytic. Synthetic study of growth forms in Mediterranean type ecosystems, Aproposal for an inter-re- gional program. Ecologia Mediterranea, 8, 159-171. [40] Gamisans, J. and Guber, H. (1980) The plant communi- ties of Niolo (Corsica). Ecologia Mediterranea, 11, 101- 113. [41] Grime, J.P. (1997) Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. The American Naturalist, 111, 1169-1194. doi:10.1086/283244 [42] Floret, C., Galan, M.J., Lefloch, E. and Romane F. (1992) Dynamics of holm oak (Quercus ilex L.) coppies after clear cutting in sounthen France. Vegetation 99-100. In: Romane, F. and Terradas, J., Eds., Quercus ilex L. Eco- systems, F unction, Dynamics and Management. Kluwer Academy Publischers, Belguin, 97-105. [43] Loisel, R. and Gamila, H. (1993) Translation effects of clearing on forest ecosystems by forest pre-disturbance index. Annales de la Societe des Sciences Naturelles et d’Archeologie de Toulon et du Var, 45, 123-132. [44] El-Hamrouni, A. (1992) Forest vegetation and pre-forest of Tunisia: Typology and elements for managing. Thesis, Es-Science, Faculty of Sciences and Techniques of Saint Jérôme, University of Aix-Marseille III, France, 220. [45] Wilson, A.D. (1986) Principles of grazing management systems. In rangelands: A resource under siege. Proceed- ings of the 2nd International Rangeland Congress, Can- berra, 1986, 221-225. [46] Quezel, P., Gamisans, J. and Guber, M. (1980) Biogeog- raphy and introduction of Mediterranean floras. Naturalia Monspeliensia, 41-51. [47] Quezel, P. (1983) Flora and vegetation of North Africa, their significance through the origin, evolution and mi- gration of flora and vegetation structures past. Bothalia, 14, 411-416. [48] Heywood, V. (1995) The Mediterranean flora in the con- text of World biodiversity. Ecologia Mediterranea, 21, 11- 18. [49] Vela, E. and Benhouhou, S. (2007) Evaluation of a new hotspot of plant biodiversity in the Mediterranean basin (North Africa). Comptes Rendus Biologies, 330, 589-605. doi:10.1016/j.crvi.2007.04.006 [50] Zeraia, L. (1981) Test of comparative interpretation of ecolo- gical data, phenological and production Suberowoody in cork oak forests of crystalline province (southern France) and Algeria. Thesis, Aix Marseille Université, Marseille. [51] Stevenson, A.C., Skinner, J., Hollis, G.E. and Smart, M. (1988) The El Kala National Park and Environs, Algeria: An ecological evaluation. Environmental Conservation, 15, 335-348. doi:10.1017/S0376892900029830 [52] Belouahem, D., Belouahem, F. and Belair, G. (2009) Flo- ristic biodiversity and vulnerability of Numidia Aulnaies glutinous Algeria (N-E Algeria). European Journal of Sci- entific Research, 32, 329-361. [53] Yahi, N., Djellouli Y. and De Foucault, B. (2008) Floristic diversity and biogeography of the cedar forests of Algeria. Acta Botanica Gallica, 155, 403-414. [54] Letreuche-Belarouci, A., Medjahdi, B., Letreuche-Bela- rouci, N. and Benabdeli, K. (2009) Floristic diversity of cork oak forests of the park national de Tlemcen (Al- geria). Acta Botanica Malacitana, 34, 1-13. [55] Medail, F. and Quezel, P. (1997) Hot-spot analysis for con- servation of plants biodiversity in the Mediterranean Ba- sin. Annals of the Missouri Botanical Garden, 84, 121- 127. doi:10.2307/2399957
|