 Vol.2, No.4, 233-243 (2012) Open Journal of Ecology http://dx.doi.org/10.4236/oje.2012.24027 Effects of micro-environmental conditions and forest disturbance on the establishment of two Andean palms in Ecuador Diana Rodríguez-Paredes1*, Rommel Montúfar-Galárraga2, Henrik Meilby1 1Forest & Landscape, University of Copenhagen, Cop e n h a g e n, Denmark; *Corresponding Author: rodriguezp.diana@gmail.com 2Ecology and Genetics, Pontificia Universidad Católica del Ecuador, Quito, Ecuador Received 17 September 2012; revised 20 October 2012; accepted 28 October 2012 ABSTRACT The wax p alm (Ceroxylon echinulatu m) and palm heart (Prestoea acuminata) are key elements in Andean natural forests. This ecosystem is th- reatened by deforestation and habitat degra- dation, thus, it is vital to explore the impact of anthropogenic activities on the ecological struc- tu re an d p r ef erences of t hese species in order to develop proper conservation strategies. Fifty- seven 400 m2 plots were established in two for- est areas, a di sturbed forest (n = 30 plots) and an undisturbed forest (n = 27 plots) in the Ecua- dorean Andes. Nine micro-environmental vari- ables and the number of individuals of C. echi- nulatum and P. acuminata in five size classes were recorded in each plot. Logistic regression models helped identify environmental variables that influence the establishment of the species. Results showed that different micro-environ- mental variables determined the presence of different size classes. Both species were bene- fitted by the environmental conditions of the dis- turbed forest. Light availability was import ant for the establish ment of both specie s , especially for the canopy species (C. echinulatum). This palm was negatively affected by the variable “fallen wood”, while P. acuminata was negatively af- fected mainly by the steepness of the terrain. The environmental variables for the successful establishment of the studied species that were identified by this study provide clues for the su- stainable management of C. echinulatum and P. acuminata in Andean forests. Keywords: Andean Forest; Ceroxylon; Forest Disturbance; Micro-Environmental Variables; Palm Establishment; Prestoea; Sustainable Manag ement 1. INTRODUCTION The palm family is important to humans as a diverse source of non-timber forest products [1,2]; however un- sustainable harvesting and deforestation are threatening the permanence of harvested species in natural forests [3-5]. Human activities modify environmental conditions in forest remnants thereby also altering the conditions under which palm species regenerate and establish [6,7]. South America has a high diversity of palms with 457 species and 50 genera [8]. Part of this diversity is found in the Andes above 1000 m.a.s.l where palms represent 37% of the palm genera and 15% of the palm species reported in the Americas [9]. Four genera have their cen- ter of distribution in the Andes: Aiphanes, Ceroxylon, Parajubaea and Wettinia; among which Cero xylon is endemic to the Andes [9]. In the Ecuadorean Andes, the most economically important palm genera are Ceroxylon and Prestoea. Leaves of the former genus are used as raw material for handicraft in Easter and unfolded leaves of the latter genus are source of palm heart or palmito [10- 13]. Palms respond to environmental heterogeneity at both large and small scales. At the large scale, the distribution of palms is mainly determined by climate [14-16], geo- logical regions [17], historical legacies [14,18,19], and to a smaller extent by edaphic characteristics [20-22], and dispersal dynamics [23]. Small scale environmental het- erogeneity (0.1 to 10 m2; [15]) is important for the distri- bution and diversity of palms [15,24,25]. Sources of mi- cro-environmental variation include canopy heterogene- ity, inter- and intra-specific relations, edaphic conditions (pH, litter) and topography. Among these variables, ca- nopy heterogeneity is suggested as an important source of variation since it can modify variables su ch as ligh t in- tensity and quality, soil, microtopography, risk of damage, pollination and seed d ispersal [15]. The dependence of many palms on humidity makes the Andean cloud forests important for palm species Copyright © 2012 SciRes. OPEN A CCESS  D. Rodríguez-Paredes et al. / Open Journal of Ecology 2 (2012) 233- 243 234 growing at high elevations. Unfortunately, this kind of forests (characterized by a constant mist and high biodi- versity and endemism) is highly threatened by deforesta- tion [26-28]. Environmental conditions of forests may change as a result of forest fragmentation [4], logging [26], harvesting of selected products [29], and/or other extractive activities affecting plant and animal population dynamics and distribution [6,30,31]. The effects of de- forestation on palm populations may be better under- stood when analyzed separately for each life stage of the individuals since some palm species change their envi- ronmental preferences during ontogeny [32]. Due to the threatened status of Andean cloud forest palms and their importance in local ecosystems, it is necessary to under- stand the effects of different environmental conditions on the regeneration, establishment and growth of harvested palm species in order to predict consequences of envi- ronmental changes on populations and as a basis for suggesting sustainable management strategies that pro- tect the target species and natural forest dynamics. Accordingly, the objective of this study is to identify micro-environmental variables that determine the distri- bution of the different life stages of Ceroxylon echinula- tum and Prestoea acuminata growing in the cloud forests of the Ecuadorean Andes. Additionally, this work aims to compare the establishment requirements of palm species with different habits (canopy and understory) and in for- ests with different degrees of disturbance (disturbed and undisturbed forest). The selection of the species was based on the following criteria: a) both species are im- portant components of the Andean forest due to their abundance and the resources they provide to the local fauna [10,33], b) their natural populations are threatened by deforestation, habitat degradation, and harvesting [34,35], c) both species are important for the local economy [1], and d) there is a lack of information about their ecology that impedes the development of proper management plans. 2. STUDY SITE The study took place in the north-western part of the Pichincha province (Figure 1). This region contains a cloud forest ecosystem at elevations ranging from 1500 to 2000 m.a.s.l. with monthly mean temperatures ranging from 18˚C to 24˚C. The rainy season can last for 10 months with maximum precipitation values in April and November. The dry season starts between July and Au- gust and lasts for two to three months [36]. In this region two localities characterized by different levels of human disturbance were selected: a) Inti Llacta reserve (dis- turbed forest), and b) Rio Bravo Reserve (undisturbed forest). The two localities are located ca. 20 km from each other. Inti Llacta (Miraflores community) is a private reserve (00˚02'N, 78˚43'W; 1875 ± 16.2 m.a.s.l) with ca. 100 ha of secondary cloud forest. This forest reserve is a mosaic of forests patches with different levels of regeneration and pasture. In this forest, selective logging and harvest- ing of palms were undertaken for several years until the creation of the reserve in 1982 . Particu larly, in the 1970’s and 1980’s the natural populations of P. acuminata were heavily harvested due to the high demand for palm heart [37]. Leaves of C. echinulatum were harvested until 2009 when the government of Ecuador banned the ex- traction from natural forests. Illegal extraction of both products from the forest still happens along with defor- estation, mining, floriculture and poultry farming, caus- ing environmental damages [34]. Rio Bravo is a private reserve (00˚04'S, 78˚44'W; 1690 ± 72.1 m.a.s.l) with ca. 4000 ha of undisturbed cloud forest (Cantón San Miguel de los Bancos) and is part of the Mindo-Nambillo protected area (19.200 ha; [34]). Due to its difficult access, the area has been relatively isolated from human activities, and therefore the forest is in a well conserved cond ition with no repo rted extraction activities. 3. STUDY SPECIES Ceroxylon echinulatum Galeano is a canopy (10 - 30 m tall; diameter 15 - 30 cm), solitary and dioecious palm [38]. Stems are covered with a layer of wax that usually have oblique ring-shaped leaf scars. This palm species prefers humid and rainy mountain regions of Ecuador and northern Perú between 1600 and 2200 m.a.s.l. Un- folded young leaves (spear leaf or cogollo) are harvested once or twice a year to weave handicrafts for Easter and/or Christmas. This harvesting activity affects mainly stem-less individuals with leaf size larger than 2 m be- cause of the easy access to the spear leaf. In Ecuador, the trade of handicrafts made with C. echinulatum’s leaves is an economic activity that represents an important source of income to local communities and artisans (Montúfar, unpublished data). Prestoea acuminata (Willd.) H. E. Moore is a mid- story species, monoecious, with solitary or clustered stems that can reach heights of 6 - 15 m and diameters of 4 - 20 cm [39]. The species has a wide distribution, grow- ing in mountain regions of the An tilles and Central Ame- rica; and premontane and montane areas (1000 - 2000 m.a.s.l) of the Andean slopes from Venezuela to Bolivia [39]. Unfolded leaves (apical meristem) of this species are harvested and sold as palm hearts in local communi- ties [10]. The extraction is done any time through the year with no discrimination betw een reproductive and non- reproductive individuals. Copyright © 2012 SciRes. OPEN A CCESS  D. Rodríguez-Paredes et al. / Open Journal of Ecology 2 (2012) 233-243 Copyright © 2012 SciRes. 235 anegalito 0 750 1500 Meters Rio Bravo Mindo Mira flo res Inti Liact a N Figure 1. Study area. Black triangles are the study sites; gray circles are clos- est towns to the study sites. 4. METHODS from each other and each transect was about 600 m long and included about 15 plots of 400 m2. This design for data collection allowed covering the topographic and environmental variation of the studied places. The num- ber of adults of both species and subadults of C. echinu- latum were recorded in the 400 m2 plots, juveniles of both species and subadults of P. acuminata were re- corded in 100 m2 (10 m × 10 m) plots nested within the 400 m2 plots, and seedlings were recorded in five sub- plots of 4 m2 (2 m × 2 m) located in the center and in the corners of each 400 m2 plot. 4.1. Census Data were collected in March and April 2010. A plot- based inventory of C. echinulatum and P. acuminata was carried out in disturbed (Inti Llacta reserve) and undis- turbed (Río Bravo reserve) forests. Individuals of C. echinulatum were classified as: 1) seedlings (individuals with undivided leaves), 2) juveniles 1 (individuals with- out stem and with pinnate leaves shorter than 2 m), 3) juveniles 2 (individuals without stem and with pinnate leaves longer than 2 m), 4) subadults (individuals with stem but without reproductive structures) and 5) adults (stemmed reproductive individuals). P. acuminata indi- viduals were classified as: 1) seedlings, 2) juveniles 1 (individuals with at least one pinnate leaf and/or with a single stem), 3) juveniles 2 (individuals with clones), 4) subadults (individuals with clones, stem and crownshaft), 5) reproductive adults with stem and/or clones. 4.2. Variables Measured Two types of variables were measured within each 400 m2 plot: topo-edaphic variables and forest structure vari- ables. The topo-edaphic variables were: 1) aspect (mea- sured using compass, degrees), 2) slope inclination (mea- sured in the steepest area of the 400 m2 plot using a cli- nometer, Sunnto Tandem; degrees), 3) leaf cover (per- centage of soil covered by leaf litter in each 4 m2 plot) and 4) leaf depth (number of leaves on the forest floor pierced with a pointed stick [40] in the center of each 4 m2 plot). The forests structure variables were: 1) can- opy height (height of the tree with the largest diameter For the data collection, a total of fifty-seven 400 m2 (20 m × 20 m) plots were established (30 plots in dis- turbed forest and 27 plots in undisturbed forest) along two linear transects in each forest (total of 4) with a dis- tance between plots centers of 40 m. In both forests transects were located at a minimum distance of 120 m OPEN A CCESS  D. Rodríguez-Paredes et al. / Open Journal of Ecology 2 (2012) 233- 243 236 measured using a clinometer Sunnto Tandem; meters), 2) basal area (of all trees with diameter at breast height (DBH) >10 cm measured using a measuring tape; m2, 3) canopy openness (measured in the center of each of the 4 m2 plots using the canopy-scope scoring system; percent; [41]), 4) gaps (absence or presence), and 5) fallen wood (number of trees or branches with diameter >10 cm lying on the floor). 4.3. Statistical Analyses Aspect values (α) were conver ted into north exposu res (cosine α) and east exposures (sine α), i.e. the lower the value of north exposure, the farther from north the aspect; similarly, for east exposure in relation to east. The five measurements of each of the variables “leaf cover”, “leaf depth” and “canopy openness” obtained for each 400 m2 plot were averaged with the purpose of obtaining a single value per 400 m2 plot to be used in the statistical analy- ses. T-tests were done in order to identify significant dif- ferences between densities of different life stages and en- vironment al va ri abl es bet ween forests. Variables that best explained the distribution (presence) of each of the life stages of the two palm species were selected using logistic regression models. In addition to the micro-environmental variables, the presence of adults was also included as a potential explanatory variable for seedlings. A Principal Component Analysis (PCA) was carried out in order to identify uncorrelated factors and thereby reduce the number of explanatory variables. PCA results were used as a reference for selecting the vari- ables to be included in the final logistic analyses. Selec- tion of independent variables in the logistic models was based on the significance level of the parameter esti- mates (p-value < 0.05), the Akaike Information Criterion (AIC), log-likelihood values and the percentage concor- dance of the model. Models with independent variables that were almost uncorrelated with each other were pre- ferred. Statistical analyses were done using the software package SAS version 9.2 (SAS Institute, INC., Cary, NC). 5. RESULTS 5.1. Description of the Forests Statistically significant differences between forests were observed for the majority of the environmental variables (Table 1), the disturbed forest show ing greater values for most of them. In the undisturbed forest the terrain was steeper than in the disturbed forest. The light related vari- ables, “canopy openness” and “gaps”, were found to hav e larger values in the disturbed forest than in the undis- turbed forest, but the difference was significant only for the former variable. The variable “canopy height” had lar- ger values in the undisturbed forest. 5.2. Population Structure within the Forest Population structures of both C. echinulatum and P. acuminata differed significantly between disturbed and undisturbed forests (Figures 2 and 3). All life stages of C. echinulatum had more individuals in the disturbed forest than in the undisturbed forest. Early life stages of P. acu- minata were more abundant in the undisturbed forest than in the disturbed forest but the opposite pattern was ob- served in later life stages. In general, and irrespective of forest type, the density of individuals of C. echinulatum decreased towards older life stages. This was also the case for P. acuminata with the exception of subadults in the disturbed forest and adults in the undisturbed forest, which were more abundant than previous life stages. For both species in both forests, about 90% of the individuals were seedlings. By comparing densities of adult individuals Table 1. Comparison of the environmental variables between disturbed and undisturbed forest using a t-test. Disturbed forest Undisturbed forest N Mean Stdev N Mean Stdev Pr > |t| Slope (˚) 30 29.067 10.998 27 35.741 8.202 0.013 Leaf cover (%) 30 87.133 11.584 27 74.389 20.701 0.007 Leaf depth (#) 30 1.793 1.191 27 1.102 0.610 0.008 Canopy height (m) 30 17.238 5.411 27 20.049 5.475 0.057 Canopy openness (%) 30 4.720 6.212 27 2.089 2.736 0.042 Gaps (#) 30 0.600 0.498 27 0.519 0.509 0.544 Fallen trees (#) 30 4.500 2.688 27 6.778 4.353 0.024 Basal area/trees (m2/ind) 30 2.120 0.866 27 1.285 0.536 0.000 N = number of plots; Stdev = standard deviation. Copyright © 2012 SciRes. OPEN A CCESS 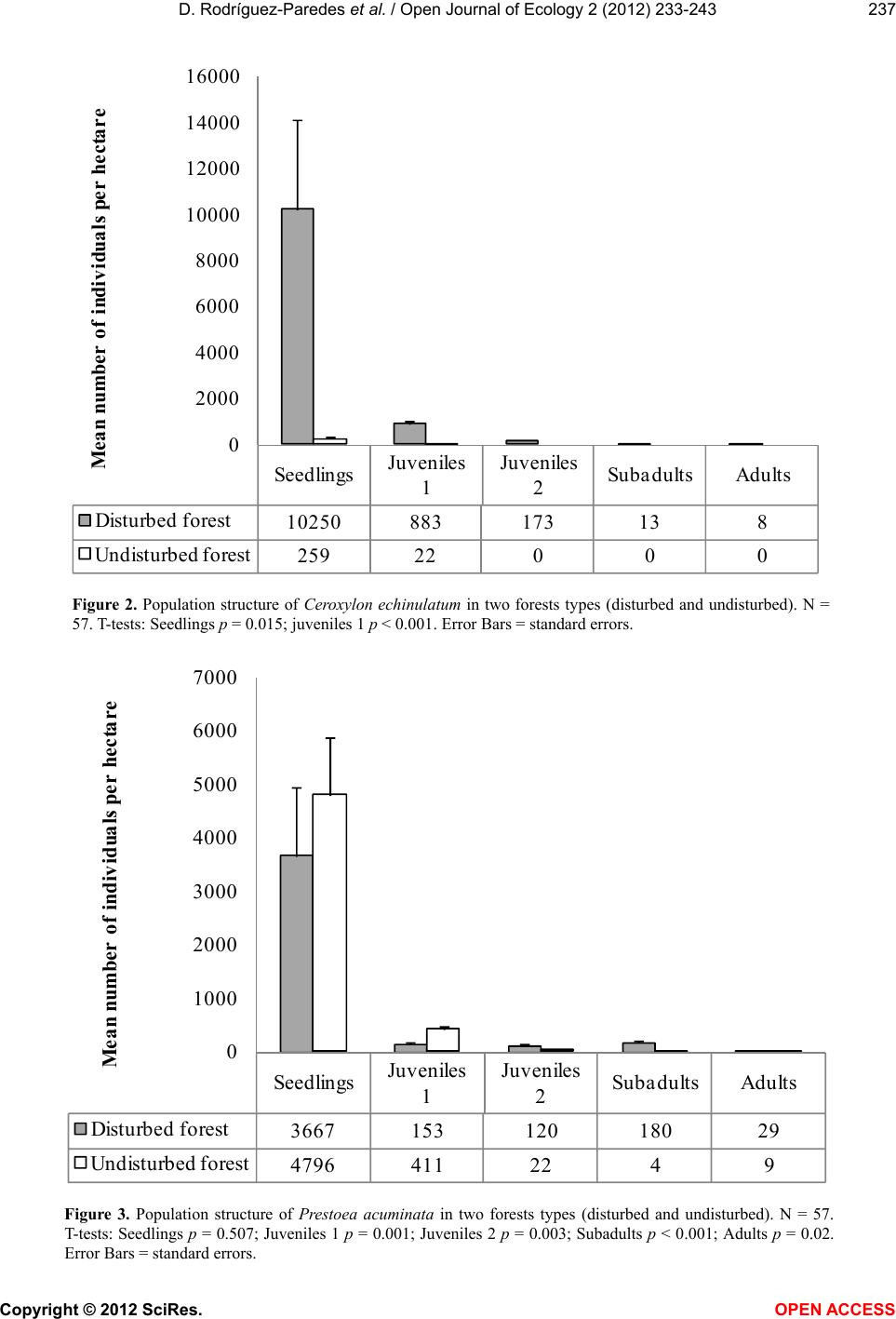 D. Rodríguez-Paredes et al. / Open Journal of Ecology 2 (2012) 233-243 237 Seedlings Juveniles 1Juveniles 2Subadults Adults Disturbed forest10250 883173138 Undi sturbed forest259 22000 0 2000 4000 6000 8000 10000 12000 14000 16000 Mean number of individuals per hectare Figure 2. Population structure of Ceroxylon echinulatum in two forests types (disturbed and undisturbed). N = 57. T-t ests: Seedlings p = 0.015; juveniles 1 p < 0.001. Error Bars = standard errors. Seedlings Juveniles 1Juveniles 2Subadults Adults Dis turbed fore st3667153 120 18029 U ndisturbed forest4796 4112249 0 1000 2000 3000 4000 5000 6000 7000 M ean number of in d ivid uals per hectare Figure 3. Population structure of Prestoea acuminata in two forests types (disturbed and undisturbed). N = 57. T-tests: Seedlings p = 0.507; Juveniles 1 p = 0.001; Juveniles 2 p = 0.003; Subadults p < 0.001; Adults p = 0.02. Error Bars = standard errors. Copyright © 2012 SciRes. OPEN A CCESS  D. Rodríguez-Paredes et al. / Open Journal of Ecology 2 (2012) 233- 243 Copyright © 2012 SciRes. OPEN A CCESS 238 and seedlings of C. echinulatum in the disturbed forest, it appeared that only about 0.081% of the seedlings are likely to reach the adult stage. A similar survival rate could not be estimated for the undisturb ed forest because no adults were found in the plots. For P. acuminata the apparent survival rates were also low; in disturbed forest only 0.79% of all seed lin gs were likely to become ad ults; in undisturbed forest the estimated survival rate was only 0.19%. 5.3. Logistic Models: Variables Determining Distribution Different variables were found to determine the distribu- tion of different life stages of the studied species (Tables 2 and 3). For C. echinulatum four out of the five forest struc- ture variables (basal area, canopy height, canopy op e n nes s and fallen wood) appeared to affect the majority of the life stages. Among the variables related to the availability of light, “canopy openness” and “canopy height” influ- enced the presence of most of the life stages. Rich-lit environ- ments appeared to benefit the establishment of the spe- cies, but the presence of gaps did not influence any of the life stages for C. echinulatum. Moreover, the absence of adults did not influence the presence of seed- lings of this species significantly. The parameters of the variables included in the best logistic models for P. acuminata were not all signifi- cantly different from zero at the 5% level. This was the case for the life stages “juveniles 1” and “adults”. Never- theless it was decided to include these models since their p-values were not far from the applied significance level (5%). For P. acuminata, the variable “slope” appeared to be the only variable that generally had a negative effect on the presence of the species. Among variables related to light conditions, “canopy openness” was important on l y for subadults and the presence of gaps was negatively related to the presence of adults. Contrary to the findings for C. echinulatum, the absence of adults of P. acuminata did appear to influence the presence of seedlings of this species. Other variables that influenced the distribution of at least one life stage were: basal area, east exposure and leaf cover. 6. DISCUSSION 6.1. Variables Determining the Presence of Seedlings As expected for this life stage, light related variables did not influence the presence of seedlings of any of the studied species since both are shade tolerant species that can germinate and establish under closed canopy condi- tions. Some studies found that seedlings of both P. acumi- nata and C. echinulatum can grow in places characte- rized by a wide range of light conditions [10,33,42], but full light exposure may have a negative effect on the es- tablishment of C. echinulatum [33]. Other factors such Table 2. Logistic regression models predicting the probability of presence of individuals of Ceroxylon echinulatum in each life stage. Model: probability = exp(a + b(x))/(1 + exp(a + b(x)). a Basal area Canopy height Canopy openness East exposure Fallen wood Gaps* Leaf Cover Slope Absence adults Concordant (m2/ha) (m) (%) (#) (%) (˚) % Seedling 1.268 0.059 –0.129 74.2 sterror 1.322 0.027 0.061 p 0.337 0.028 0.036 Juvenile1 –1.543 0.098 0.312 1.951 –0.318 88.9 sterror 1.216 0.037 0.153 0.671 0.127 p 0.204 0.009 0.041 0.004 0.013 Juvenile2 –2.138 0.062 0.344 –0.323 84.3 sterror 1.18 0.029 0.138 0.133 p 0.07 0.034 0.013 0.015 Subadult 5.072 –0.241 0.135 –0.095 81.8 sterror 2.409 0.101 0.071 0.042 p 0.035 0.017 0.055 0.025 Adults 9.208 –0.551 –1.012 95 sterror 4.247 0.246 0.489 p 0.03 0.025 0.038 a = intercept.*presence gaps. Sterror = standard error.  D. Rodríguez-Paredes et al. / Open Journal of Ecology 2 (2012) 233-243 239 Tab le 3 . Logistic regression models predicting the probability of presence of individuals of Prestoea acuminata in each life stage. Model: probability = exp(a + b(x))/(1 + exp(a + b(x)). a Basal area Canopy height Canopy openness East exposure Fallen wood Gaps*Leaf Cover Slope Absence adults concordant (m2/ha) (m) (%) (#) (%) (˚) % Seedling 5.863 –0.055 –0.674 72.9 sterror 2.367 0.027 0.360 p 0.013 0.038 0.061 Juvenile 1 3.268 –0.066 65.1 sterror 1.281 0.035 p 0.011 0.060 Juvenile 2 –0.478 0.893 64.4 sterror 0.295 0.446 p 0.105 0.045 Subadult –1.576 0.089 0.258 –0.079 85.2 sterror 1.451 0.032 0.117 0.036 p 0.277 0.005 0.028 0.030 Adult 1.455 –0.551 –0.054 70.1 sterror 0.978 0.289 0.030 p 0.137 0.057 0.066 a = intercept; *presence gaps; Sterror = s tandard error. as temperature or humidity are presumably more impor- tant for seedling establishment in both C. echinulatum and P. acuminata. Madriñán and Schultes [43] suggested that the germination of Ceroxylon quinduense required special conditions of high humidity. Leaf litter may be an important element d etermining the estab lishment o f seed- lings since it modifies micro-habitat conditions [44,45], pro- tects seeds from predation [46], or enhances fungi attack [47]. In accordance with the study by Didham and Law- ton [48], the present study found more leaf litter in the disturbed forest. The low abundance of P. acuminata in this forest may be to a negative effect of leaf litter on germination. A high canopy was found to negatively affect the estab- lishment of seedlings of C. echinulatum. This result may be explained by the negative relation between the num- ber of adults and the canopy height as also reported by Paredes [49] on western slopes for the same species. The lower height of the canopy in the disturbed forest could explain the higher number of seedlings of C. echinulatum in this forest. Fo r P. acuminata the presence of seedlings did depend on the absence of adults. This result sugg ests that spatial dispersion of P. acuminata is very low. As a consequence, the risk of death for seeds and seedling of this species may be high according to Janzen and Con- nell’s model [50]. 6.2. Variables Determining the Presence of Juveniles Shade tolerance in palms generally declines with on- togeny [32]. This characteristic may be particularly evi- dent for canopy palms whose adult form is found in rich- lit canopy environments [51,52]. This is in agreement with this study, where light availability was found to posi- tively affect the presence of juveniles of the canopy spe- cies (C. echinulatum) only. Mortality as a result of fallen pieces of wood may be a good indicator of conditions affecting the establishment of juveniles of C. echinulatum in the undisturbed forest. According to Laurance i.e. [31] more tree damage is ex- pected in disturbed forests as a result of lower water availability and stronger wind impact. Accordingly, the disturbed forest was expected to have more “fallen wood ” than the undisturbed forest, but the opposite result was found in this study. The reason may be related to specific characteristics of the former forest such as the steepness of the terrain and/or the predominant plant species. Fallen wood may also affect juveniles of P. acuminata, however; variables such as “slope” of the terrain and “as- Copyright © 2012 SciRes. OPEN A CCESS  D. Rodríguez-Paredes et al. / Open Journal of Ecology 2 (2012) 233- 243 240 pect” seemed to be more important. The steepness of the terrain in the undisturbed forest did not favor the estab- lishment of individuals of P. acuminata, probably as a result of mortality due to soil instability and small land- slides. As expected, juveniles 1 of P. acuminata were af- fected by the steepness of the terrain s ince the y are small in size and/or have only one slim stem. Any damage to a single-stemmed individual would kill it [53]. The variable “east exposure” appeared to influence the abundance of juveniles of both species positively. Other studies also reported an effect of aspect on the distribu- tion of C. echinulatum [54] and P. acuminata [55]. The vari- able “basal area” positive ly affected the early stages of C. echinulatum. This r esult is in agreemen t with Anth elme i.e. [33], who found that the basal area of regenerating indi- viduals (20 - 30 cm DBH) positively affected the abun- dance of C. echinulatum in disturbed forests. 6.3. Variables Determining the Presence of Subadults In C. echinulatum, the end of the establishment phase and the beginning of the subadult phase is determined by the appearance of the stem [32]. By contrast, in P. acu- minata stem diameter and height increase simultaneously [56]. Regardless of the differences in the stem develop- ments, both species showed faster growth early in the subadult phase [57,58 ]. In this phase of fast growth, light availability is believed to be important for the establish- ment of subadults especially for the canopy species that needs the rich-lit condition above the canopy to develop reproductive structures. As expected, the distribution of subad ults of both species was positivel y related with lig ht availability. The negative relationship between the abun- dance of subadults of C. echinulatum and the light relate d variable “canopy height” provides additional evidence of the dependence of this canopy palm on rich-lit environ- ments. “Slope” appeared to be one of the determinants of the presence of subadults for both species. For C. echinula- tum this may be explained by low recruitment into this stage, as previous stem-less stages may be more vulner- able to die from landslides. In the case of P. acuminata, a partial explanation may be that the small diameter of its stem makes it easy to bend. In the case of single-stem individuals, the bending may cause death 6.4. Variables Determining the Presence of Adults Adults of the canopy species (C. echinulatum) are be- lieved to rely more on well-lit environments than adults of the midstory species (P. acuminata) [52]. Light related variables such as gaps and canopy openness were not part of the final model for the canopy palm; but the impor- tance of light for adult establish ment for this species was indicated by the variable canopy height which was part of the final model. Under tall canopies, individuals have to grow more before reaching the full light above the canopy, which will give them the energy to produce re- productive organs. Similar results were found by Paredes [49] for the same species in the eastern Andes. The pre- sent study found lower canopy heights in the disturbed forest which may be one of the reasons for the success of C. echinulatum in this particular forest. The variable “f al- len wood” appeared to affect the presence of adults of C. echinulatum negatively. This result was not expected since adult palms are tall individuals with large diameters and strong stems; however Paredes [49] also reported adult’s death as a result of fallen trees. As expected for P. acuminata, light was not as relevant as for C. echinulatum given that the former species th- rives well in relative ly shaded conditions. This resu lt con- trasts with the observations by Svenning [55] for the same species. However results have to be interpreted with caution since, as explained before, the best model obtained for adults of this species included variab les with p-values slightly above the chosen significance level of 0.05. In this case, a larger sample size would be needed in order to achieve a better understanding of the pre- ferred establishment conditions for P. acuminata’s adults. 6.5. Conservation and Management Strategies Despite the harvesting pressure that C. echinulatum and P. acuminata experienced in the past, their estab- lishment and population development is benefited by the environmental conditions found in the disturbed forest. The present study identified importan t elements that help understanding the preferred environmental conditions of the studied species; information that may be of vital im- portance for their conservation since it can be applied in the development of sustainable management plans for forest and when analyzing possible impacts of changing environments on populations of the two species. Based on the findings of this study the following management actions are suggested: 1) Steep slopes negatively affect the establishment of the studied species; thus it is recom- mended to focus management activities on flat areas where palm populations are dense. Populations on steep slopes should be protected as a source of genetic vari- ability for these species; 2) Light benefits the establish- ment of the majority of the life stages in C. echinulatum and P. acuminata. Hence, reducing the density of the canopy may be a way to enhance the growth of the spe- cies. This method is particularly promising for the first life stages since they are more susceptible to mortality; 3) Copyright © 2012 SciRes. OPEN A CCESS  D. Rodríguez-Paredes et al. / Open Journal of Ecology 2 (2012) 233-243 241 Leaf litter negatively affects the establishment of seed- lings in P. acuminata. It is therefore recommended to re- move part of the leaf litter that covers the seedlings; 4) Connectivity of forest remnants is important for the abi- lity of pollinators and seed dispersers to move between the forests, especially for the dioic species (C. echinula- tum). Connectivity also enhances genetic diversity which is important for the species to be resilient to a changing environment; and 5) Forests with different disturbance regimes have different environmental conditions. The present study provides an important first attempt to un- derstand the response of the two palm species to distur- bances. Future research should study the population dy- namic of C. echinulatum and P. acuminata in forest rem- nants of different sizes, as well as the relation of disper- sers and pollinators with these palm species. 7. ACKNOWLEDGEMENTS This study was funded by the University of Copenhagen, the Eras- mus Mundus Programme of the European Union, and 7th Framework Program “PALMS” (FP-7, grant 212631). We thank Nina Duarte and Inti Llacta family and Sonia Saltos from Rio Bravo for their help dur- ing the fieldwork. We also thank Luis Saltos, Juan Lincango, and Manuel for their collaboration during the data collection. D. Rodríguez is grateful to Henrik Balslev for his continuous support. We thank Luis Cárdenas for his technical support with the map. REFERENCES [1] Brokamp, G., Valderrama, N., Mittelbach, M., Grández, C., Barfod, A. and Weigend, M. (2011) Trade in palm products in north-western South America. The Botanical Review, 77, 571-606. doi:10.1007/s12229-011-9087-7 [2] Dransfield, J., Uhl, N., Asmussen, C., Baker, W., Harley M. and Lewis, C. (2008) Genera palmarum: The evolu- tion and classification of palms. Kew Publishing, Royal Botanic Gardens, Kew. [3] Galetti, M. and Fernández, J. (1998) Palm heart harves- ting in the Brazilian Atlantic Forest: Changes in industry structure and the illegal trade. Journal of Applied Ecology, 35, 294-301. [4] Steffan-Dewenter, I. and Tscharntke, T. (1999) Effects of habitat isolation on pollinator communities and seed set. Oecologia, 121, 432-44 0. doi:10.1007/s004420050949 [5] Zuidema, P. and Boot, R. (2000) Demographic constraints to sustainable palm heart extraction from a sub-canopy palm in Bolivia. In: Zuidema, P., Ed., Demography of Ex- ploited Tree Species in the Bolivian Amazon, Riberalta: Promab, Chapter 3, Riberalta. [6] Laurance, W., Ferreira, L., Rankin-De Merona, J., Lau- rance, S., Hutchings, R. and Lovejoy, T. (1998) Effects of forest fragmentation on recruitment patterns in Amazo- nian tree communities. Conservation Biology, 12, 460- 464. doi:10.1046/j.1523-1739.1998.97175.x [7] Williams-Linera, G. (1990) Vegetation structure and en- vironmental conditions of the forest edges in Panamá. Journal of Ecology, 78, 356-373. doi:10.2307/2261117 [8] Pintaud, J., Galeano, G., Balslev, H., Bernal, R., Bor- chsenius, F., Ferreira, E., De Granville, J., Mejía, K., Millán, B., Moraes, M., Noblick, L., Stauffer, F. and Kahn, F. (2008) Las palmeras de América del Sur: Diversidad, distribución e historia evolutiva. Revista Pe- ruana de Biología, 15, 7-29. [9] Borchsenius, F. and Moraes, M. (2006) Diversidad y usos de las palmeras Andinas (Arecaceae). In: Moraes, M., et al. Eds., Botánica económica de los Andes centrales. Uni- versidad Mayor San Andrés, La Paz, Bolivia. [10] Bonilla, D. and Feil, J. (1995) Production of ramets and germination of Prestoea trichoclada (Arecaceae): A source of palm heart in Ecuador. Principes, 39, 210-214. [11] Borchsenius, F., Borgtoft-Pedersen, H. and Balslev, H. (1998) Manual to the palms of Ecuador. AAU Reports 37, University of Aarhus, Aarhus. [12] De La Torre, L., Navarrete, H., Muriel, P., Macia, M. and Balslev, H. (2008) Enciclopedia de las plantas útiles del Ecuador. Herbari o QCA de la Escuela de ciencia biológicas de la Pontificia Universidad Católica del Ecuador y Herbario AAU del Departamento de Ciencias Biológicas de la Universidad de Aarhus, Quito and Aarhus. [13] Pedersen, H. and Balslev, H. (1992) The economic botany of Ecuadorean palms. In: Plotkin, M. and Famolare, L., Eds., Sustainable Harvest and Marketing of Rain Forests Products, Islands Press, Washington DC, 173-191. [14] Kissling, D.W., Baker, W.J., Balslev, H., Barfod, A.S., Borchsenius, F., Dransfield, J., Govaerts, R. and Sven- ning, J. (2011) Quaternary and pre-quaternary historical legacies in the global distribution of a major tropical plant lineage. Global Ecology and Biogeography, 21, 909-921. [15] Svenning, J. (2001) On the role of micro-environmental heterogeneity in the ecology and diversification of neo- tropical Rain-forest palms (Arecaceae). Botanical Review, 67, 1-53. doi:10.1007/BF02857848 [16] Skov, F. and Borchsenius, F. (1997) Predicting plant spe- cies distribution patterns using simple climatic parame- ters: A case study of Ecuadorean palms. Ecography, 20, 347-355. doi :10 .1111/j .1600-0587.1997.tb00379.x [17] Henderson, A. (1995) The palms of the Amazon. Oxford University Press, Oxford. doi:10.1126/science.165.3889.131 [18] Haffer, J. (1969) Speciation in Amazonian forest birds. Science, 165, 131-137. doi:10.1126/science.165.3889.131 [19] Montúfar, R. and Pintaud, J. (2006) Variation in species composition, abundance and microhabitat preferences among western Amazonian terra firme palm communities. Botanical Journal of the Linnean Society, 151, 127-140. d oi :10. 1111/j .1 0 95 -8339.2006.00528.x [20] Clark, D., Clark, D., Sandoval, M. and Castro M. (1995) Edaphic and human effects on landscape-scale distribu- tions of tropical rain forest palms. Ecology, 76, 2581- 2594. doi:10.2307/2265829 [21] Svenning, J. (1999) Microhabitat specialization in a spe- cies-rich palm community in Amazonian Ecuador. Jour- nal of Ecology, 87, 55-65. Copyright © 2012 SciRes. OPEN A CCESS  D. Rodríguez-Paredes et al. / Open Journal of Ecology 2 (2012) 233- 243 242 doi:10.1046/j.1365-2745.1999.00329.x [22] Vormisto, J., Hanna, T. and Jari, O. (2004) Palm distribu- tion patterns in Amazonian rainforests: What is the role of topographic variation. Journal of Vegetation Science, 15, 485-494. doi :10 .1111/j .1654-1103.2004.tb02287.x [23] Karubian, J., Sork, V. L., Roorda, T., Duraes, R. and Smith, T. (2010) Destination-based seed dispersal homogenizes genetic structure of a tropical palm. Molecular Ecology, 19, 1745-1753. doi: 10. 1111/j .1365 -294X.2010.04600.x [24] Souza, A. and Martins, F. (2004) Microsite specialization and spatial distribution of Geonoma brevispatha a clonal palm in south-eastern Brazil. Ecological Research, 19, 521- 532. d oi: 10.1111 /j. 1440-1703.2004.00670.x [25] Svenning, J. (2000). Small canopy gaps influence plant distributions in the rain forest understory. Biotropica, 32, 252-261. [26] Bubb, P., May, I., Miles, L. and Sayer, J. (2004) Cloud forest. http://www.unep-wcmc.org/resources/publications/UNEP _WCMC_bio_series/20.htm [27] Cuesta, F., Peralvo, M. and Valarezo, N. (2009) Los bos- ques montanos de los Andes Tropicales. Una evaluación re gio n a l de s u estad o de c onservació n y de su vulner abilid ad a efectos del cambio climático. Programa Regional ECO- BONA-Intercooperation, Imprenta Mariscal, Quito. [28] Josse, C., Cuesta, F., Navarro, G., Barrena, V., Cabrera, E., Chacón-Moreno, E., Ferreira, W., Peralvo, M., Saito, J. and Tovar, A. (2009) Ecosistemas de los Andes del Norte y Centro. Bolivia, Colombia, Ecuador, Perú y Venezuela. Secretaría General de la Comunidad Andina, programa Regional ecobona-intercooperation, codesan-proyecto pá- ramo andino, programa bio-andes, ecociencia, nature serve, iavh, lta-unalm, icae-ula, cdc-unalm, rumbol srl. Lima. [29] Montúfar, R., Anthelme, F., Pintaud, J. and Balslev, H. (2011) Disturbance and resilience in tropical American palm populations and communities. Botanical Review, 77, 426-461. doi:10.1007/s12229-011-9085-9 [30] Kapos, V. (1989) Effects of isolation on the water status of forest patches in the Brazilian Amazon. Journal of Tropical Ecology, 5, 173-185. doi:10.1017/S0266467400003448 [31] Laurance, W., Ferreira, L., Rankin-De Merona, J. and Laurence, S. (1998) Rain forest fragmentation and the dy- namics of Amazonian tree communities. Ecology, 79, 2032-2040. doi:10.1890/0012-9658(1998)079[2032:RFFATD]2.0.CO ;2 [32] Tomlinson, P. (1990) The structural biology of palms. Oxford University Press, New York. [33] Anthelme, F., Lincango, J., Gully, C., Duarte, N. and Mon- túfar, R. (2011) How anthropogenic disturbances affect the resilience of a keystone palm tree in the threatened Andean cloud forest. Biological Conservation, 14 4, 1059- 1067. doi:10.1016/j.biocon.2010.12.025 [34] Guevara, M., Fuentes-Pozo, P., Josse, C. and Peñafiel, M. (2001) Tres décadas de cambios en el uso de la tierra en el área de Nanegal. In: Rhoades, R.E., Ed., Tendiendo puentes entre los paisajes humanos y naturales. La in- vestigación participativa y el desarrollo ecológico en una frontera agrícola andina, Abya Yala, Quito, 416. [35] Jensen, A. and Meilby, H. (2008) Does commercialization of non-timber forest product reduce ecological impact. A case study of critically endangered aquilaria crassna in Lao PDR. Oryx, 42, 214-221. doi:10.1017/S0030605308007825 [36] Cañadas-Cruz, L. (1983) El mapa bioclimático y eco- lógico del Ecuador. Banco Central del Ecuador. Quito, Ecuador. [37] Knudsen, H. (1995) Demography, palm-heart extracti- vism, and reproductive biology of P. acuminata (Areca- ceae) M.Sc. Thesis, Department of Systematic Botany, University of Aarhus, Aarhus. [38] Sanín, M. J. and Galeano, G. (2011). A revision of the an- dean wax palm, Ceroxylon (Arecaceae). Phytotaxa, 34, 1- 64. [39] Henderson, A., Galeano, G. and Bernal, R. (1995) Field guide to the palms of the Americas. Princenton University Press, New Jersey. [40] Daws, M., Pearson, T., Burslem, D., Mullins, C. and Dal- ling, J. (2005) Effects of topographic position, leaf litter and seed size on seedling demography in a semi-decidu- ous tropical forest in Panamá. Plant Ecology, 17 9, 93-105. doi:10.1007/s11258-004-5801-4 [41] Brown, N., Jennings, S., Wheeler, P. and Nabe-Nielsen, J. (2000) An improved method for the rapid assessment of the forest understorey light environments. Journal of Ap- plied Ecology, 37, 1044-1053. doi:10.1046/j.1365-2664.2000.00573.x [42] Svenning, J. (1998) The effect of land-use on the local distribution of palm species in an Andean rain forest fragment in northwestern Ecuador. Biodiversity and Con- servation, 7, 1529-1537. doi:10.1023/A:1008831600795 [43] Madriñán, S. and Schultes, R. (1995) Colombia’s national tree: The wax palm Ceroxylon quindiuense and its rela- tives. Elaeis, 7, 35-56. [44] Molofsky, J. and Augspurger, C. (1992) The effects of leaf litter on early seedling establishment in a tropical forest. Ecology, 73, 68-77. doi:10.2307/1938721 [45] Vázquez-Yanes, C., Orozco-Segovia, A., Rincón, E., Sán- chez-Coronado, M., Huante, P., Toledo, J. and Barradas, V. (1990) Light beneath the litter in a tropical forest: Ef- fect on seed germination. Ecology, 71, 1952-1958. doi:10.2307/1937603 [46] Cintra, R. (1997) Leaf litter effects on seed and seedling predation of the palm Astrocaryum murumuru and the legume tree Dypteryx micrantha in Amazonian forest. Journal of Tropical Ecology, 13, 709-725. doi:10.1017/S0266467400010889 [47] Rudas, C. (1998) Evaluación del estado actual de una población de la palma de cera, Ceroxylon sasaimae Gale- ano: Aportes a su historia de vida y estudio demográfico. B.Sc. Thesis, Universidad de Los Andes, Bogotá. [48] Didham, R. and Lawton, J. (1999) Edge structure deter- mines the magnitude of changes in microclimate and vegetation structure in tropical forests fragments. Bio- Copyright © 2012 SciRes. OPEN A CCESS  D. Rodríguez-Paredes et al. / Open Journal of Ecology 2 (2012) 233-243 Copyright © 2012 SciRes. OPEN A CCESS 243 tropica, 31, 17-30. [49] Paredes, T. (1995) Primeros estudios biológicos de la palma de ramos (Ceroxylon echinulatum) presente en Co- sanga (provincia del Napo) entre Agosto de 1991 y Octubre de 1992. Tesis de Licenciatura, Departamento de Biología, Pontificia Universidad Católica del Ecuador, Quito. [50] Schupp, E. (1992) The janzen-connell model for tropical tree diversity: Population implications and the importance of spatial scale. The American Naturalist, 140, 526-530. doi:10.1086/285426 [51] Kahn, F. (1986) Life forms of Amazonian palms in rela- tion to forest structure and dynamics. Biotropica, 18, 214- 218. doi:10.2307/2388487 [52] Svenning, J. (1999) Recruitment of tall arborescent palms in the Yasuní National Park, Amazonian Ecuador: Are large treefall gaps important. Journal of Tropical Ecology, 15, 355-366. doi:10.1017/S0266467499000875 [53] Svenning, J. (2000) Growth strategies of clonal palms (Arecaceae) in a neotropical rainforest, Yasuní, Ecuador. Australian Journal of Botany, 48, 167-178. doi:10.1071/BT98048 [54] Galeano, G. (1995) Novedades del género Ceroxylon (Pal- mae). Caldasia, 17, 395-408. [55] Svenning, J. (2001) Environmental heterogeneity, recr- uitment limitation and the mesoscale distribution of palms in a tropical montane rain forest (Maquipucuna, Ecuador). Journal of Tropical Ecology, 17, 97-113. doi:10.1017/S0266467401001067 [56] Henderson, A. (2002) Evolution and ecology of palms. The New York Botanical Garden press, New York. [57] Lugo, A. and Rivera, C. (1987) Leaf production, growth rate, and age of the palm Prestoea Montana in the Lu- quillo experimental forest, Puerto Rico. Journal of Tropi- cal Ecology, 3, 151-161. doi:10.1017/S0266467400001905 [58] Vergara, L. (2002) Demografía de ceroxylon alpinum en bosques relictuales del Valle de Cocora, Salento (Quin- dio). B.Sc. Thesis, Departamento de Biología, Universi- dad Nacional de Colombia, Bogotá.
|