Rolling Circle Amplification Is More Sensitive than PCR and Serology-Based
Methods in Detection of Banana streak virus in Musa Germplasm
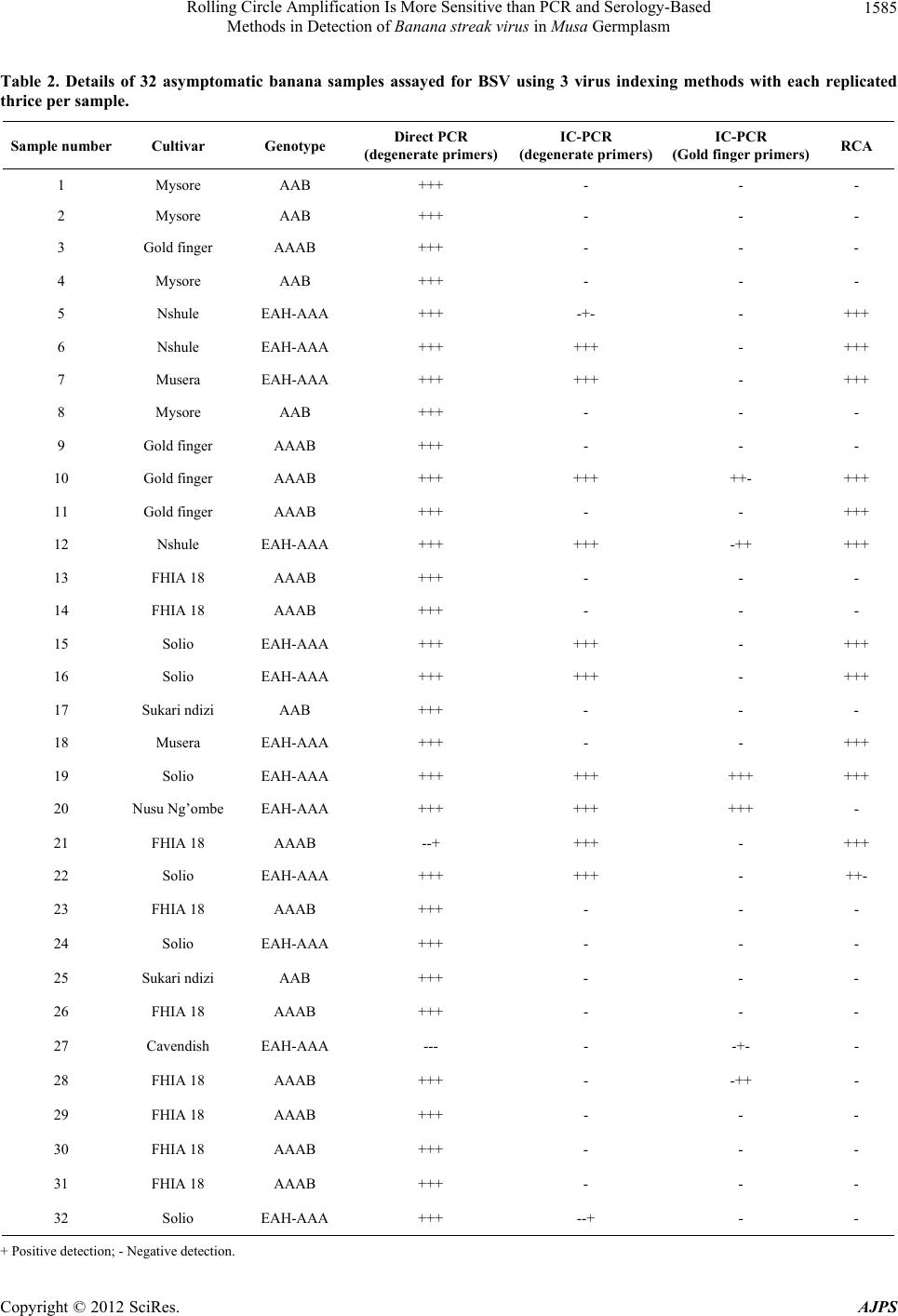
1587
5. Acknowledgements
We would like to thank the Queensland University of
Technology (QUT), Australia, through the Banana 21
project for funding this work. Special thanks also go to
the Kenya Agricultural Research Institute, Njoro for
availing laboratory facilities.
REFERENCES
[1] J. W. Daniells, A. D. W. Geering, N. J. Brynde and J. E.
Thomas, “The Effect of Banana streak virus on the
Growth and Yield of Dessert Bananas in Tropical Austra-
lia,” Annals of Applied Biology, Vol. 139, No. 1, 2001, pp.
51-60. doi:10.1111/j.1744-7348.2001.tb00130.x
[2] B. E. L. Lockhart and D. R. Jones, “Banana Streak,” In: D.
R. Jones, Ed., Diseases of Banana, Abacá and Ensete,
CABI Publishing, Oxon, 1999, pp. 262-274.
[3] B. E. L. Lockhart, T. C. Ndowora, N. E. Olszewski and G.
Dahal, “Studies on Integration of Banana streak bad-
navirus Sequences in Musa: Identification of Episomally-
Expressible Badnaviral Integrants in Musa Genotypes,”
In: E. A. Frison and S. E. Sharrock, Eds., Banana streak
virus: A Unique Virus—Musa Interaction: Proceedings of
a Workshop of the PROMUSA Virology Working Group,
Montpellier, 19-21 January 1998, pp. 35-42.
[4] A. D. W. Geering, J. N. Parry, L. Zhang, N. Olszewski, B.
E. L. Lockhart and J. E. Thomas, “Is Banana streak virus
Strain OL the Only Activatable Virus Integrant in the
Musa Genome?” Proceedings of the 2nd International
Conference on Molecular Cellular Biology of Banana,
InfoMusa, Vol. 10, No. 2, 2001, pp. 13-20.
[5] G. Harper, R. Hull, B. E. L. Lockhart and N. Olszewski,
“Viral Sequences Integrated into Plant Genomes,” Annual
Review of Phytopathology, Vol. 40, 2002, pp. 119-136.
doi:10.1146/annurev.phyto.40.120301.105642
[6] International Network for the Improvement of Banana
and Plantain (INIBAP), “Report of the Meeting,” In: E. A.
Frison and S. E. Sharrock, Eds., Banana streak virus: A
Unique Virus—Musa Interaction: Proceedings of a
Workshop of the PROMUSA Virology Working Group,
Montpellier, 19-21 January 1997, pp. 7-14.
[7] G. Thottappilly, G. Dahal and B. E. L. Lockhart, “Studies
on a Nigerian Isolate of Banana streak badnavirus. I. Pu-
rification and Enzyme-Linked Immunoassay,” Annals of
Applied Biology, Vol. 132, No. 2, 1998, pp. 253-261.
doi:10.1111/j.1744-7348.1998.tb05201.x
[8] B. E. L. Lockhart and N. E. Olszewski, “Serological and
Genomic Heterogeneity of Banana streak badnavirus:
Implications for Virus Detection in Musa Germplasm,” In:
J. Ganry, Ed., Breeding Banana and Plantain for Resis-
tance to Diseases and Pests, CIRAD, Montpellier, 1993,
pp. 105-130.
[9] D. Haible, S. Kober and H. Jeske, “Rolling Circle Ampli-
fication Revolutionizes Diagnosis and Genomics of Ge-
miniviruses,” Journal of Virological Methods, Vol. 135,
No. 1, 2006, pp. 9-16.
doi:10.1016/j.jviromet.2006.01.017
[10] N. J. Gawel and R. L. Jarret, “A Modified CTAB Extrac-
tion Procedure for Musa and Ipomoea,” Plant Molecular
Biology Reporter, Vol. 9, No. 3, 1991, pp. 262-266.
doi:10.1007/BF02672076
[11] A, P. James, R. J. Geijskes, J. L. Dale and R. M. Harding,
“Development of a Novel Rolling—Circle Amplification
Technique to Detect Banana streak virus Which Also
Discriminates between Integrated and Episomal Virus
Sequences,” Plant Disease, Vol. 95, No. 1, 2011, pp. 57-
62.
[12] G. Harper, D. Hart, S. Moult and R. Hull, “Detection of
Banana streak virus in Field Samples of Bananas from
Uganda,” Annals of Applied Biology, Vol. 141, No. 3,
2002b, pp. 247-257.
doi:10.1111/j.1744-7348.2002.tb00216.x
[13] M. Bousalem, E. J. P. Douzery and S. E. Seal, “Taxon-
omy, Molecular Phylogeny and Evolution of Plant Re-
verse Transcribing Viruses (Family Caulimoviridae) In-
ferred from Full-Length Genome and Reverse Transcrip-
tase Sequences,” Archives of Virology, Vol. 153, No. 6,
2008, pp. 1085-1102. doi:10.1007/s00705-008-0095-9
[14] L. Blanco and M. Salas, “Relating Structure to Function
in Phi29 DNA Polymerase,” Journal of Biological Chem-
istry, Vol. 271, No. 15, 1996, pp. 8509-8512.
doi:10.1074/jbc.271.15.8509
[15] L. Blanco, A. Bernad, J. M. Lazaro, G. Martin, C. Gar-
mendia and M. Salas, “Highly Efficient DNA Synthesis
by the Phage Phi29 DNA Polymerase,” Journal of Bio-
logical Chemistry, Vol. 264, No. 15, 1989, pp. 8935-
8940.
[16] B. O. Agindotan, G. Thottappilly, A. Uwaifo and S. Win-
ter, “Production of Monoclonal and Polyclonal Antibod-
ies against a Nigerian Isolate of Banana streak virus,” Af-
rican Journal of Biotechnology, Vol. 2, No. 7, 2003, pp.
171-178.
[17] L. Karanja, A. Wangai, G. Harper and R. S. Pathak, “Mo-
lecular Identification of Banana streak virus Isolates in
Kenya,” Journal of Phytopathology, Vol. 156, No. 11-12,
2008, pp. 678-686.
doi:10.1111/j.1439-0434.2008.01415.x
[18] G. Dahal, F. Dahal, C. Pasberg-Gauhl, J. D. A. Hughes, G.
Thottapilly and B. E. L. Lockhart, “Evaluation of Micro-
propagated Plantain and Banana (Musa spp.) for Banana
streak badnavirus Incidence Under Field and Screen-
house Conditions in Nigeria,” Annals Applied Biology,
Vol. 134, No. 2, 1999, pp. 181-191.
doi:10.1111/j.1744-7348.1999.tb05254.x
Copyright © 2012 SciRes. AJPS