Shoot Biomass Assessments of the Marine Phanerogam Zostera marina for Two Methods of Data Gathering
1544
and processing sufficient data to test a robust statistical
method, could be an extremely time consuming endeavor.
For that reason some authors use aggregated data or data
at a shoot level instead of data at a leaf level (non-ag-
gregated) to test differences of some measurements in
time and space [13]. Moreover, data gathering for eel-
grass assessments methods could be time consuming and
destructive. For example at a leaf level, each leaf in a
shoot should be counted, measured (length and width)
and weighted implying a enormous time of sample proc-
essing and for biomass (for example) a bigger error in
weighting each leaf separately.
On the other hand, aggregated data, could be easier to
obtain and present less error, but could not be used in
some models that require individual measurements, as
Equation (1).
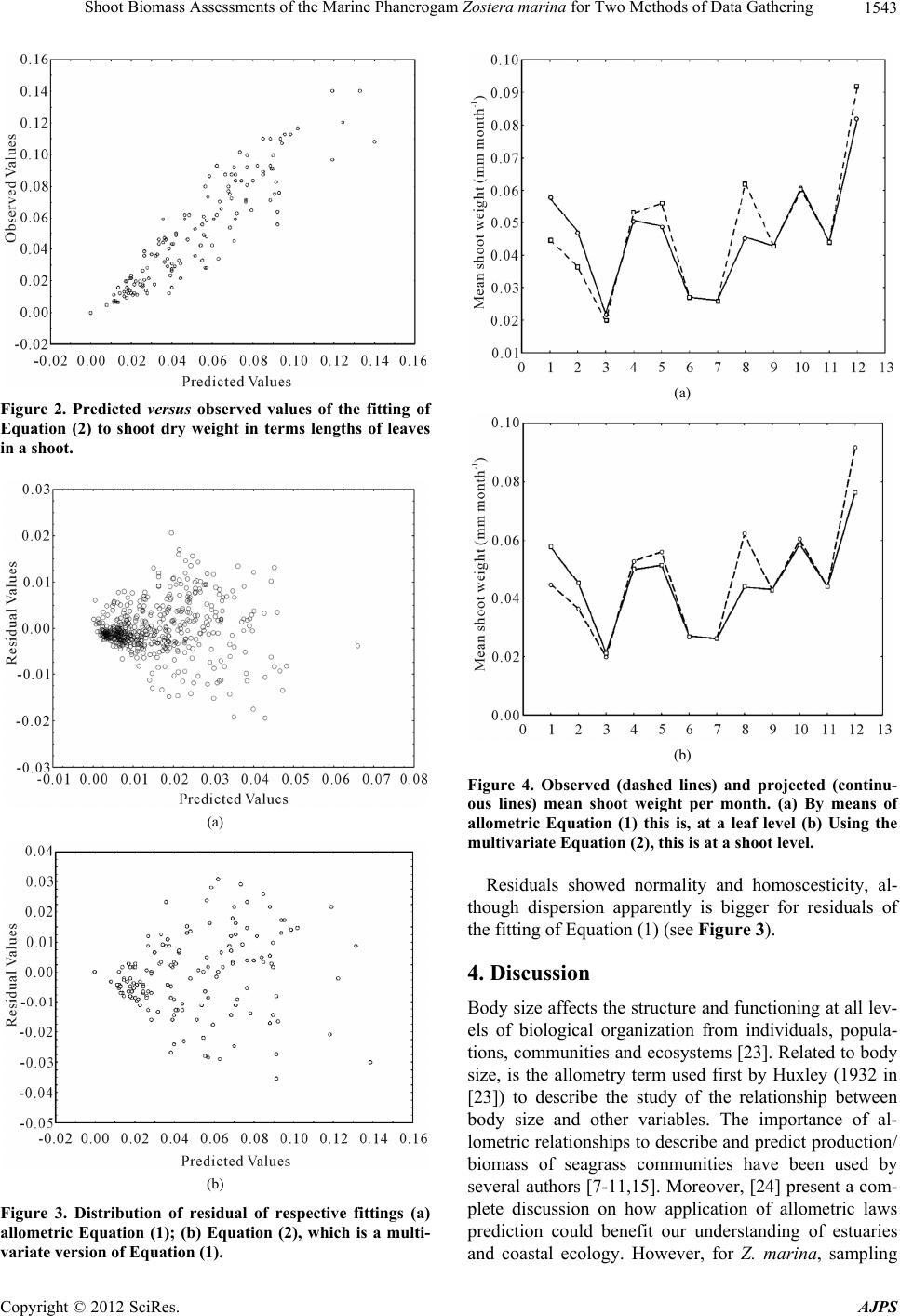
We have demonstrated in this work, that for biomass
assessments a several-variables version of the allometric
Equation (1), Equation (2) where leaf dry weights are
aggregated at a shoot level gives the same results as the
assessments found for the fitting of Equation (1). We
found that parameters fitted for both models were statis-
tically the same with p = 0.11 for
and p = 0.50 for
.
The determination coefficients for both fittings were
also statistically the same (p > 0.05). Moreover Lin [19]
Concordance Correlation Coefficient of reproducibility
(ˆ
) attained exactly the same value for both fits (0.92),
and The AIC shows that both models deserve considera-
tion for statistical inference. Figure 4 shows that pro-
jected values of mean shoot biomass per month are al-
most identical and in a good correspondence with ob-
served values, therefore we consider that Equations (1)
and (2) can be used indistinctly for shoot biomass as-
sessments. Nevertheless, the fitting of Equation (1) gives
us the advantages of a smaller estimation error, but with
the disadvantage of bigger time consuming in data proc-
essing. Whereas in the fitting of Equation (2) processing
data is much less time consuming but have a slight
higher estimation error and a better disposition of residu-
als. Moreover, since the time of processing material in
aggregated form is much less time consuming, bigger
samples can be taken if necessary. In any case, regardless
of the type of data (aggregated or non-aggregated), the
allometric relationship between leaf or shoot dry weight
and leaf length is consistent for Zostera marina. In con-
clusion, for shoot weight assessments, we proposed
model of Equation (2) for large samples and faster data
processing and model of Equation (1) if data at leaf level
is required for other endeavors.
5. Acknowledgements
The authors thank Jose Maria Dominguez and Francisco
Ponce for the art work.
REFERENCES
[1] F. T. Short and C. A. Short, “The Seagrass Filter: Purifi-
cation of Coastal Water,” In: V. S. Kennedy, Ed., The
Estuary as a Filter, Academic Press, Orlando, 1984, pp.
395-413.
[2] G. W. Thayer, W. J. Kenworthy and M. S. Fonseca, “The
Ecology of Eelgrass Meadows of the Atlantic Coast: A
Community Profile,” US Fish and Wildlife Service Divi-
sion of Biological Services, FWS/OBS-84/02, No. 147,
1984.
[3] L. G. Ward, W. M. Kemp and W. R. Boynton, “The In-
fluence of Waves and Seagrass Communities on Sus-
pended Particulates in an Estuarine Embayment,” Journal
of Marine Geology, Vol. 59, No. 1-4, 1984, pp. 85-103.
doi:10.1016/0025-3227(84)90089-6
[4] M. S. Fonseca and J. S. Fisher, “A Comparison of Can-
opy Friction and Sediment Movement between Four Spe-
cies of Seagrass with Reference to Their Ecology and
Restoration,” Marine Ecology Progress Series, Vol. 29,
1986, pp. 15-22. doi:10.3354/meps029015
[5] T. P. Williams, J. M. Bubb and J. N. Lester, “Metal Ac-
cumulation within Salt Marsh Environment: A Review,”
Marine Pollution Bulletin, Vol. 28, No. 5, 1994, pp. 277-
290. doi:10.1016/0025-326X(94)90152-X
[6] K. L. Heck, K. W. Able, C. T. Roman and M. P. Fahay,
“Composition, Abundance, Biomass and Production of
Macrofauna in a New England Estuary: Comparison
among Eelgrass Meadows and Other Nursery Habitats,”
Estuaries, Vol. 18, No. 2, 1995, pp. 379-389.
doi:10.2307/1352320
[7] S. P. Hamburg and P. S. Homann, “Utilization of Growth
Parameters of Eelgrass, Zostera marina, for Productivity
Estimates under Laboratory and in Situ Conditions,” Ma-
rine Biology, Vol. 93, No. 2, 1986, pp. 299-303.
doi:10.1007/BF00508267
[8] E. Solana-Arellano, D. J. Brobon-Gonzalez and H. A.
Echavarria-Heras, “A General Allometric Model for Blade
Production in Zostera marina L.,” Bulletin of Southern
California Academy of Sciences, Vol. 97, 1998, pp. 39-
48.
[9] C. M. Duarte, “Temporal Biomass Variability and Pro-
duction/Biomass Relationships of Seagrass Communi-
ties,” Marine Ecology Progress Series, Vol. 51, 1989, pp.
269-276. doi:10.3354/meps051269
[10] H. Echavarria-Heras, K. S. Lee, E. Solana-Arellano and E.
Franco-Vizcaíno, “Formal Analysis and Evaluation of
Allometric Methods for Estimating Above-Ground Bio-
mass of Eelgrass,” Annals of Applied Biology, Vol. 159,
No. 3, 2011, pp. 503-515.
doi:10.1111/j.1744-7348.2011.00511.x
[11] E. Solana-Arellano, S. E. Ibarra-Obando and H. A. Echa-
varria-Heras, “Calibración de un Modelo Alométrico para
Evaluar la Producción Foliar de Zostera marina L.,”
Hidrobiológica, Vol. 1, 1991, pp. 41-44.
[12] K. Aioi, “Seasonal Change in the Standing Crop of Eel-
grass Zostera marina L. in Odawa Bay, Central Japan,”
Aquatic Botany, Vol. 8, 1980, pp. 343-354.
doi:10.1016/0304-3770(80)90064-9
Copyright © 2012 SciRes. AJPS