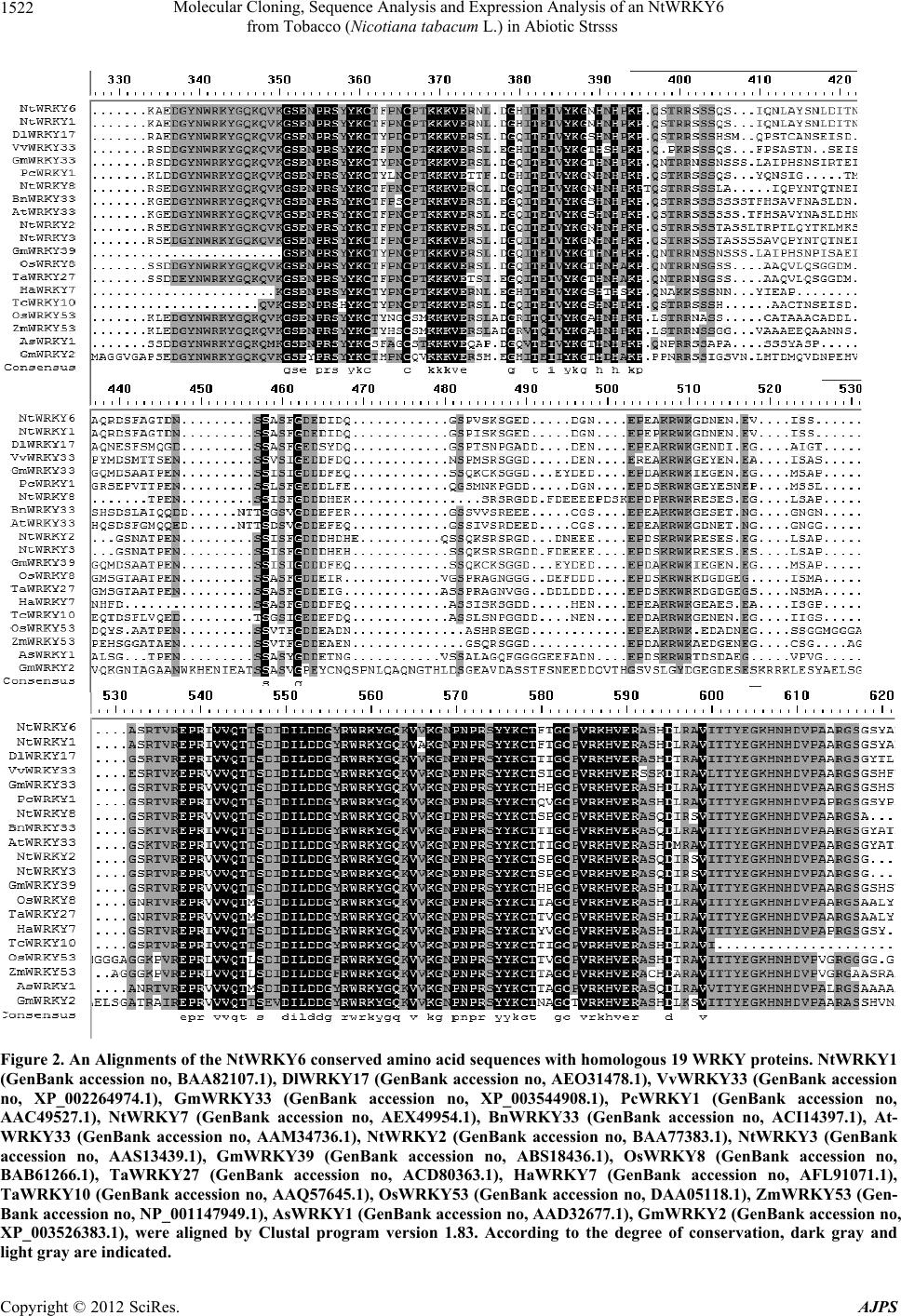
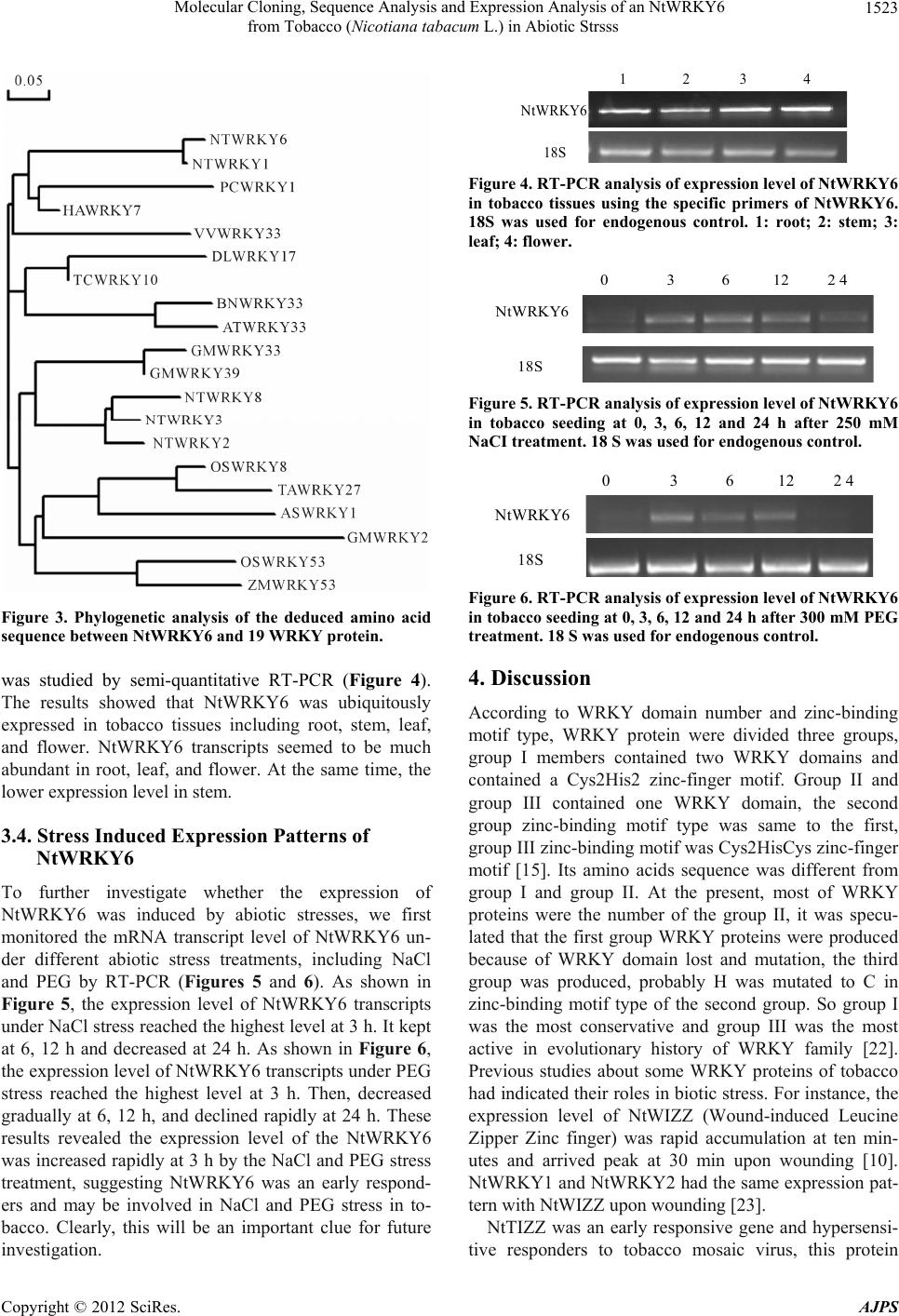
Molecular Cloning, Sequence Analysis and Expression Analysis of an NtWRKY6
from Tobacco (Nicotiana tabacum L.) in Abiotic Strsss
1525
[8] P. Z. Yang, C. H. Chen, Z. P. Wang, B. F. Fan and Z. X.
Chen, “A Pathogen and Salicylic Acid-Induced WRKY
DNA-Binding Activity Recognizes the Elicitor Response
Element of the Tobacco Class I Chitinase Gene Pro-
moter,” The Plant Journal, Vol. 18, No. 2, 1999, pp. 141-
149.
[9] L. Du and Z. Chen, “Identification of Genes Encoding
Receptorlikeprotein Kinases as Possible Targets of Patho-
gen-Andsalicylic Acid-Induced WRKY DNA-Binding
Proteins in Arabidopsis,” The Plant Journal, Vol. 24, No.
6, 2000, pp. 837-847.
[10] K. Hara, M. Yagi, T. Kusano and H. Sano, “Rapid Sys-
temic Accumulation of Transcripts Encoding a Tobacco
WRKY Transcription Factor upon Wounding,” Molecu-
lar Genetics and Genomics, Vol. 263, No. 1, 2000, pp.
30-37. doi:10.1007/PL00008673
[11] S. Robatzek and I. E. Somssich, “Targets of AtWRKY6
Regulation during Plant Senescence and Pathogen De-
fense,” Genes & Development, Vol. 16, 2002, pp. 1139-
1149. doi:10.1101/gad.222702
[12] M. Luo, E. S. Dennis, F. Berger, W. J. Peacock and A.
Chaudhury, “A MINI SEED3 (MINI3), a WRKY Family
Gene, and HAIKU2 (IKU2), a Leucine-Rich Repeat
(LRR) Kinase Gene, Are Regulators of Seed Size in
Arabidopsis,” Proceedings of the National Academy of
Sciences of the USA, Vol. 102, No. 48, 2005, pp. 17531-
17536. doi:10.1073/pnas.0508418102
[13] B. N. Devaiah, A. S. Karthikeyan and K. G. Raghothama,
“WRKY75 Transcription Factor Is a Modulator of Phos-
phate Acquisition and Root Development in Arabidop-
sis,” Plant Physiology, Vol. 143, No. 4, 2007, pp. 1789-
1801.
[14] Y. F. Chen, L. Q. Li, Q. Xu, Y. H. Kong, H. Wang and W.
H. Wu, “The WRKY6 Transcription Factor Modulates
PHOSPHATE1 Expression in Response to Low Pi Stress
in Arabidopsis,” The Plant Cell, Vol. 21, No. 11, 2009,
pp. 3554-3566. doi:10.1105/tpc.108.064980
[15] T. Eulgem, P. J. Rushton, S. Robatzek and I. E. Somssich,
“The WRKY Superfamily of Plant Transcription Fac-
tors,” Trends in Plant Science, Vol. 5, No. 5, 2000, pp.
199-206. doi:10.1016/S1360-1385(00)01600-9
[16] P. J. Rushton, H. Macdonald, A. K. Huttly, C. M. Lazarus
and R. Hooley, “Members of a New Family of DNA-
Binding Proteins Bind to a Conserved cis-Element in the
Promoters of Alpha-Amy2 Genes,” Plant Molecular Bi-
ology, Vol. 29, No. 4, 1995, pp. 691-702.
[17] M. Skibbe, N. Qu, I. Galis and I. T. Baldwin, “Induced
Plant Defenses in the Natural Environment: Nicotiana
Attenuata WRKY3 and WRKY6 Coordinate Responses
to Herbivory,” The Plant Cell, Vol. 20, No. 7, 2008, pp.
1984-2000. doi:10.1105/tpc.108.058594
[18] Y. L. Zhuang, G. J. Ren, C. M. He, X. Y. Li, Q. M. Meng,
C. F. Zhu, R. C. Wang and J. Zhang, “Cloning and Char-
acterization of a Maize cDNA Encoding Glutamate De-
carboxylase,” Plant Molecular Biology Reporter, Vol. 28,
No. 4, 2010, pp. 620-626.
[19] X. M. Liu, J. M. Anderson and P. M. Pijut, “Cloning and
Characterization of Prunus serotina AGAMOUS, a Puta-
tive Flower Homeotic Gene,” Plant Molecular Biology
Reporter, Vol. 28, No. 2, 2010, pp. 193-203.
doi:10.1007/s11105-009-0140-1
[20] K. Tamura, J. Dudley, M. Nei and S. Kumar, “MEGA4:
Molecular Evolutionary Genetics Analysis (MEGA)
Software Version 4.0,” Molecular Biology and Evolution,
Vol. 24, No. 8, 2007, pp. 1596-1599.
[21] L. J. Wu, X. T. Wang, L. C. Wu, P. G. Wang and Y. H.
Chen, “Molecular Cloning and Expression Analysis of an
HINT1Homologue from Maize (Zea mays L.),” Plant
Molecular Biology Reporter, Vol. 29, 2011, pp. 1006-
1012.
[22] B. Ulker and I. E. Somssich, “WRKY Transcription Fac-
tors: From DNA Binding towards Biological Function,”
Current Opinion in Plant Biology, Vol. 263, No. 5, 2004,
pp. 491-498. doi:10.1016/j.pbi.2004.07.012
[23] T. Nishiuchi, H. Shinshi and K. Suzuki, “Rapid and Tran-
sient Activation of Transcription of the ERF3 Gene by
Wounding in Tobacco Leaves Possible Involvement of
NtWRKYs and Auto Repression,” Journal of Biological
Chemistry, Vol. 279, No. 53, 2011, pp. 55355-55361.
doi:10.1074/jbc.M409674200
[24] H. Yoda, M. Ogawa, Y. Yamaguchi, N. Koizumi, T. Ku-
sano and H. Sano, “Identification of Early-Responsive
Genes Associated with the Hypersensitive Response to
Tobacco Mosaic Virus and Characterization of a WRKY-
Type Transcription Factor in Tobacco Plants,” Molecular
Genetics and Genomics, Vol. 267, No. 2, 2002, pp. 154-
161. doi:10.1007/s00438-002-0651-z
[25] N. Ishihama, R. Yamada, M. Yoshioka, S. Katou and H.
Yoshioka, “Phosphorylation of the Nicotiana Bentha-
miana WRKY8 Transcription Factor by MAPK Functions
in the Defense Response,” The Plant Cell, Vol. 23, No. 3,
2011, pp. 1153-1170.
[26] L. Rizhsky, H. Liang and R. Mittler, “The Combined
Effect of Drought Stress and Heat Shock on Gene Ex-
pression in Tobacco,” Plant Physiology, Vol. 130, No. 3,
2012, pp.1143-1151. doi:10.1104/pp.006858
[27] F. Wang, X. L. Hou, J. Tang, Z. Wang, S. M. Wang, F.
Jiang and Y. Li, “A Novel Cold-Inducible Gene from
Pak-Choi (Brassica campestrisssp. chinensis), BcWRKY46,
Enhances the Cold, Salt and Dehydration Stress Toler-
ance in Transgenic Tobacco,” Molecular Biology Reports,
Vol. 39, No. 4, 2012, pp. 4553-4564.
doi:10.1007/s11033-011-1245-9
[28] Q. Q. Zhang, J. H. Zhu, Y. M. Ni, Y. B. Cai and Z. L.
Zhang, “Expression Profiling of HbWRKY1, an Ethe-
phon-Induced WRKY Gene in Latex from Hevea brasil-
iensis in Responding to Wounding and Drought,” Trees,
Vol. 26, No. 2, 2012, pp. 587-595.
doi:10.1007/s00468-011-0623-1
[29] X. L. Zou, Q. Shen and D. Neuman, “An ABA Inducible
WRKYgene Integrates Responses of Creosote Bush (Lar-
rea tridentata) to Elevated CO2 and Abiotic Stresses,”
Plant Science, Vol. 172, No. 5, 2004, pp. 997-1004.
doi:10.1016/j.plantsci.2007.02.003
[30] R. Y. Guo, F. F. Yu, Z. Gao, H. L. An, X. C. Cao and X.
Q. Guo, “GhWRKY3, a Novel Cotton (Gossypium hirsu-
Copyright © 2012 SciRes. AJPS