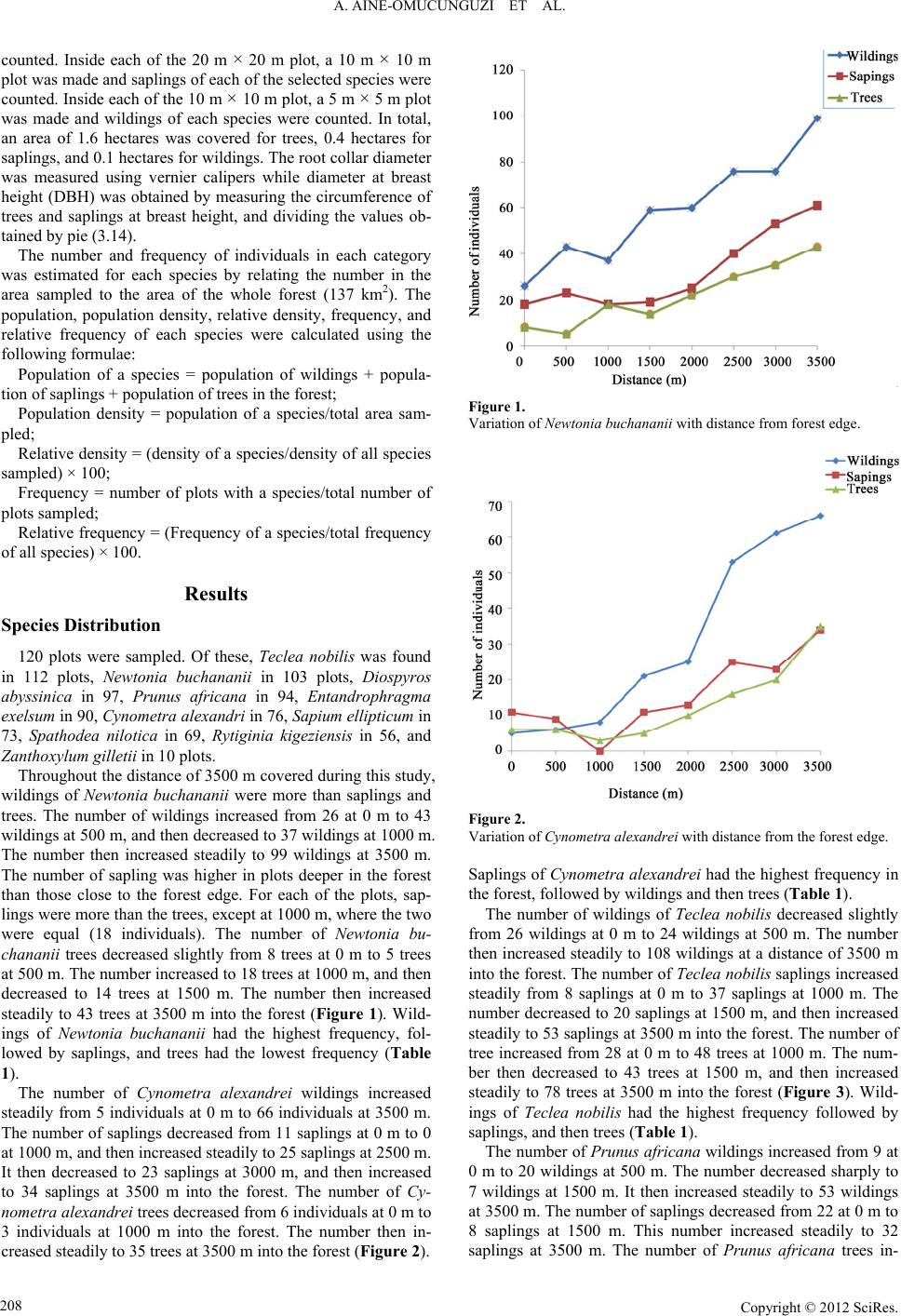
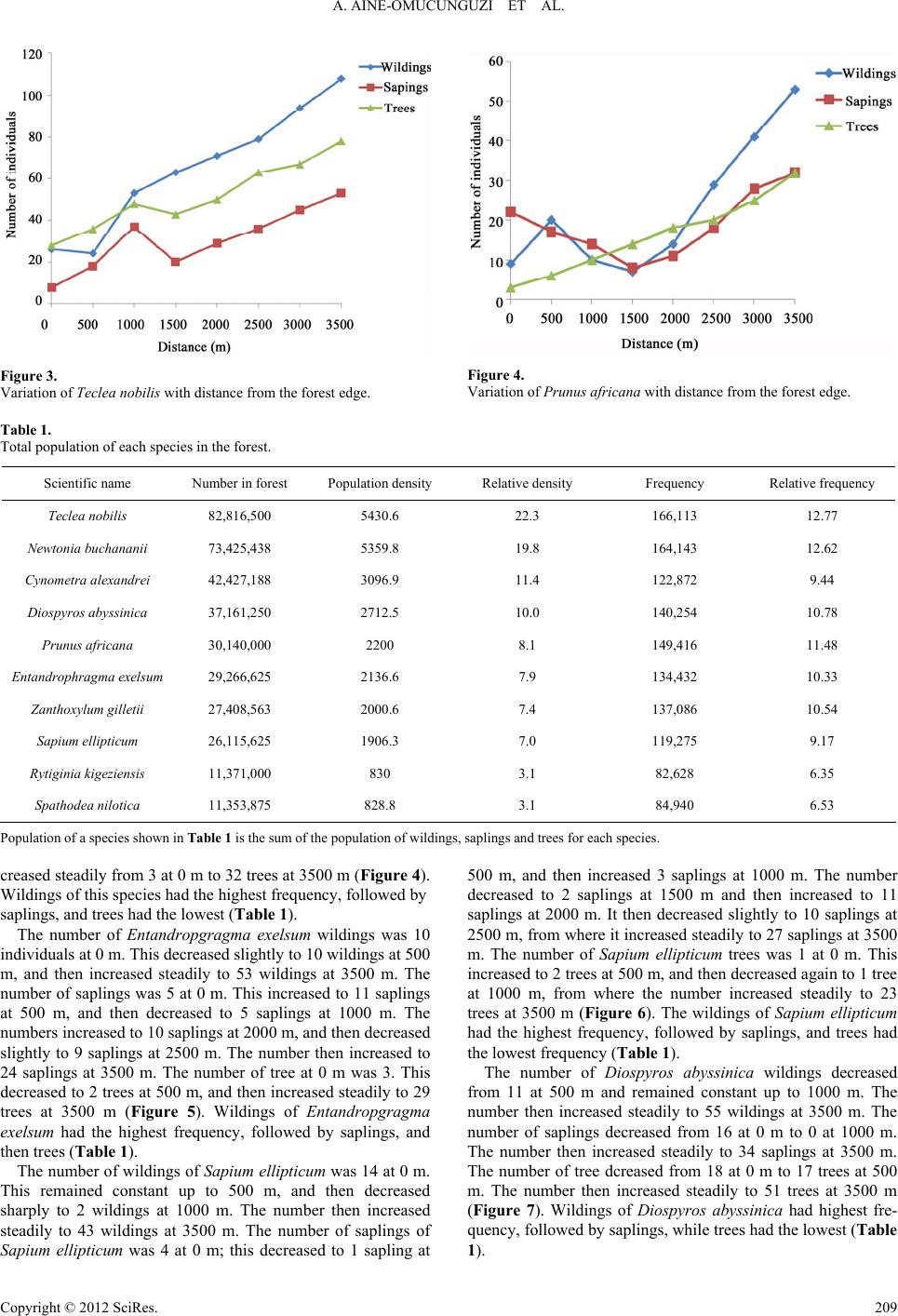
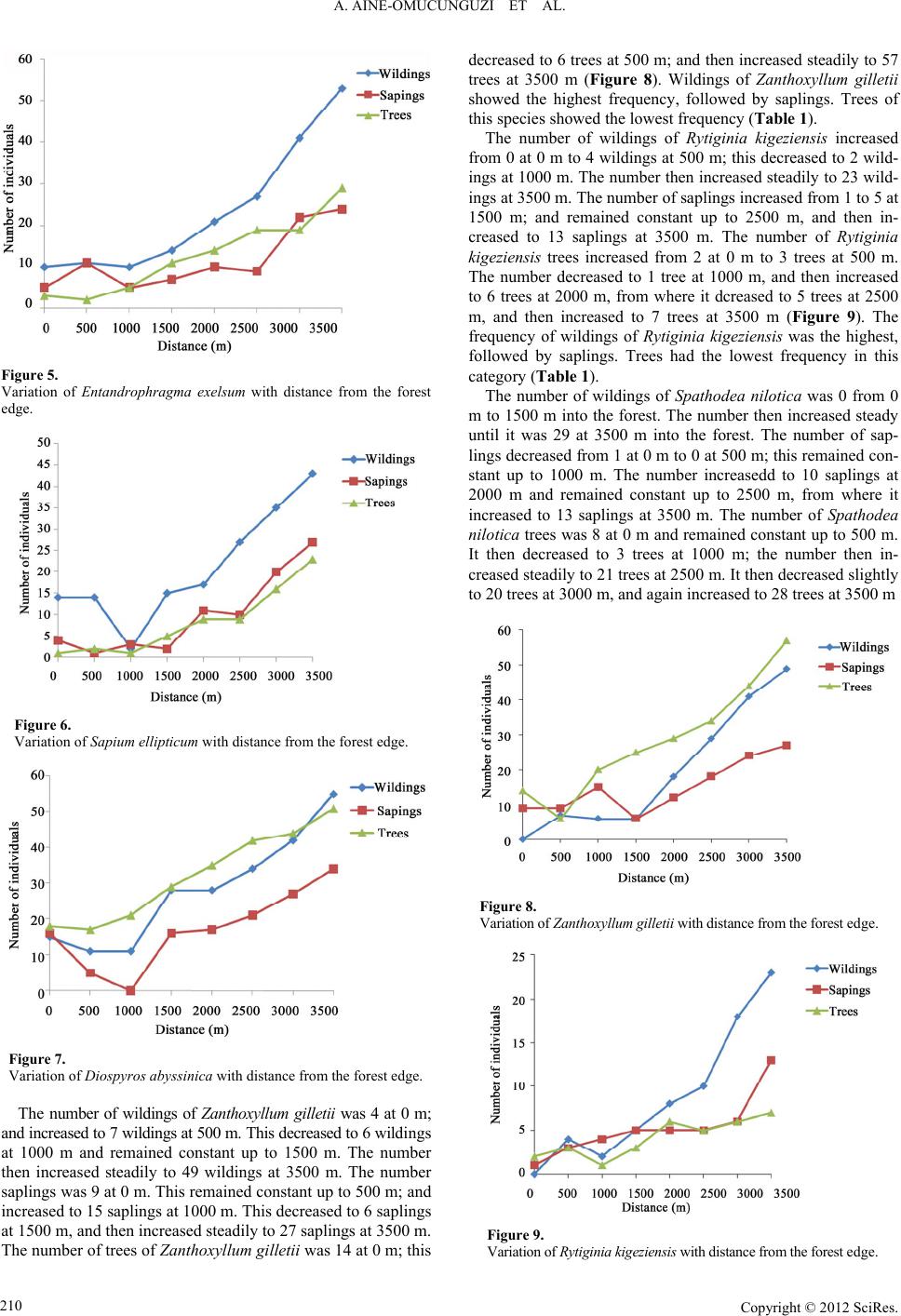
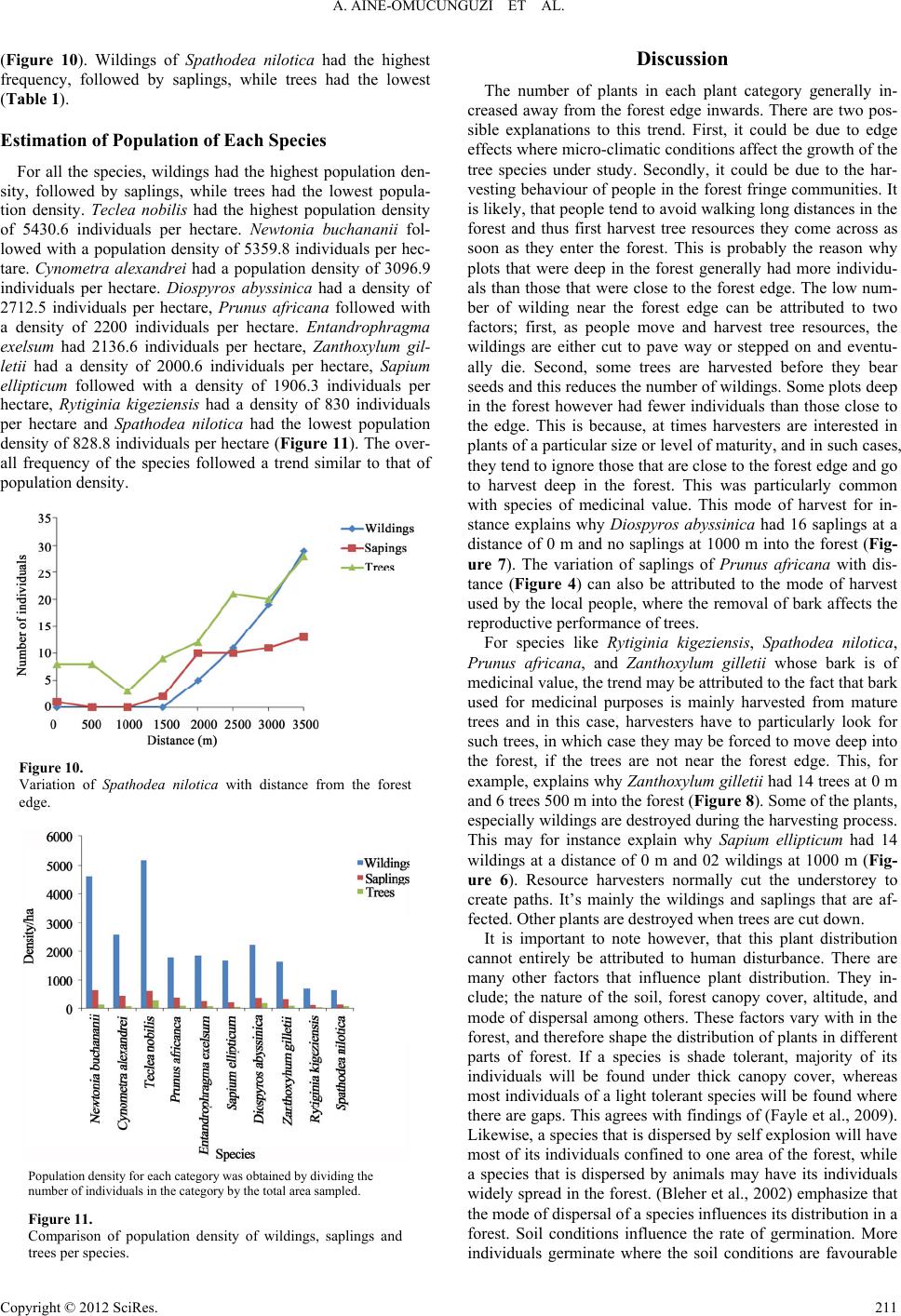
A. AINE-OMUCUNGUZI ET AL.
Copyright © 2012 SciRes.
212
(Nazre et al., 2009). It is important to note that tree populations
with low densities and clumped distributions are at risk in the
face of increased human dependence on forest products.
Frequency is a good measure of species distribution. Species
with high frequencies are frequently encountered in the forest,
and are widely distributed, while those with very low frequen-
cies are either confined to particular parts of the forest, or are
scarce. This kind of distribution is usually due to high rate of
harvest, nature of soil, mode of dispersal, or altitude. The high
frequency of Teclea nobilis (Table 1) for instance, suggests that
it is widely distributed in the forest, while the low frequency of
Spathodea nilotica is an indication that it is not common in the
forest. Comparing the frequency of wildings, saplings and trees
of a given species gives an insight into the stage of the plant
that is affected most by harvesting. The low frequency of trees
in this study suggests that they are the most affected by har-
vesting, and the high frequency of wildings suggests that they
are the least affected by harvesting, while the saplings are in-
termediate. The high frequency of wildings for most of the
species suggests that they are widely distributed in the forest
and that their numbers are still high due to low levels of harvest.
The low frequency of Cynometra alexandrei wildings com-
pared to other species is an indication that the population of this
species is on the decline. This could probably be attributed to
the high demand of this species for fuel wood. Most of the trees
of this species are harvested before fruiting and this has greatly
reduced its seed bank.
Conclusion and Recommendations
For all the species studied, the low number of trees implies
that there are fewer individuals contributing to the forest seed
bank. If the rate of tree harvest is not regulated, the seed bank
may in the long run be depleted. However, the high number of
wildings and saplings suggests that the population of most of
these species is generally stable. It’s worth noting however, that
human disturbance may in some cases increase resilience of
some species. Effective conservation of these tree species re-
quires strategies that specify the age and size of plants from
which to harvest, and the frequency of harvest. Since wildings
are normally destroyed during the harvesting process, it’s im-
portant to also specify parts of the forest in which harvesting is
allowed. These parts will then need effective monitoring. All
this will only be possible, if the local people are involved in the
decision making process and management of the forest.
Changes in population can be used to show the intensity of
harvesting of a species; however it was quite hard in this par-
ticular study, because no previous data was available to com-
pare populations of particular species in order to determine the
population decline or increase over time. It is therefore hoped
that the results of this study will provide a basis for studying
changes in the population of these ten species. Never the less,
the low population of Zanthoxylum gilletii, Rytiginia kigezien-
sis and Spathodea nilotica, can be attributed to the destructive
methods used to harvest their bark, which in most cases result
into death of the affected plants.
Species with low numbers are mainly those with medicinal
value and they have been massively and destructively harvested
for a long time. This has tremendously affected their distribu-
tion in the forest. They are now left very deep in the forest.
There is a need to sensitize the resource harvesters about re-
sponsible harvesting of plant resources. This will minimize the
destruction of the understorey during the harvesting process.
There is need to incorporate the resource users in the manage-
ment process of Kalinzu Forest Reserve if their negative im-
pacts on the forest are to be minimized. Their incorporation
requires a clear understanding of the socio-economic attributes
that shape their forest resource demands.
Acknowledgements
This work was funded by the Toyota Foundation of Japan
through the Kalinzu Forest Project. We are grateful to Professor
Takeshi Furuichi and Associate Professor Chie Hashimoto of
the Kalinzu Forest Project for the financial and technical help
they gave to us. We are also indebted to the local people around
Kalinzu Forest Reserve for all the assistance extended to us
during the study.
REFERENCES
Aine-Omucunguzi, A., Rugunda, G. K., & Byarugaba, D. (2010). Utili-
zation of forest plant resources by the local people around Kalinzu
forest reserve, south-western Uganda. African Journal of Ecology, 48,
819-821.
Alder, D., & Synnot, T. J. (1992). Permanent sample plot techniques
for mixed tropical forest. Teopical Forestry Paper No. 25, Oxford:
Oxford Forestry Institute.
Bleher, B., Oberrath, R., & Bohning-Gaese, K. (2002). Seed dispersal,
breeding systems, tree density and the spatial pattern of trees—A
simulation approach. Basic and Applied E cology, 3, 115-123.
doi:10.1078/1439-1791-00088
Eilu, G. (1995). The natural regeneration of canopy tree species fol-
lowing human disturbance in Bwindi impenetrable forest, Uganda.
Master’s thesis, Kampala: Makerere University.
Fayle, T. M., Chung, A. Y. C., Dumbrell, A. J., Eggleton, P., & Foster
W. A. (2009). The effects of rain forest canopy architecture on the
distribution of epiphytic ferns (Asplenium spp.) in Sabah Malaysia.
Biotropica, 41, 676-681. doi:10.1111/j.1744-7429.2009.00523.x
Howard, P. (1991). Nature conservation in Uganda’s tropical forest
reserves. Gland and Cambridge: International Union for Conserva-
tion of Nature.
Nakibuka, M. (1994). Some aspects of tropical rain forest regeneration
following encroachment in the Mabira forest reserve. Dissertation,
Kampala: Makerere University.
Nazre, M., Latiff, A., & Mohamad-Roslan, M. K. (2009). Effects of
topography and soil on the distribution of under canopy trees of Gar-
cinia (Guttiferae) in lowland forest of Peninsular Malaysia. Interna-
tional Journal of Botany, 5, 287-294.