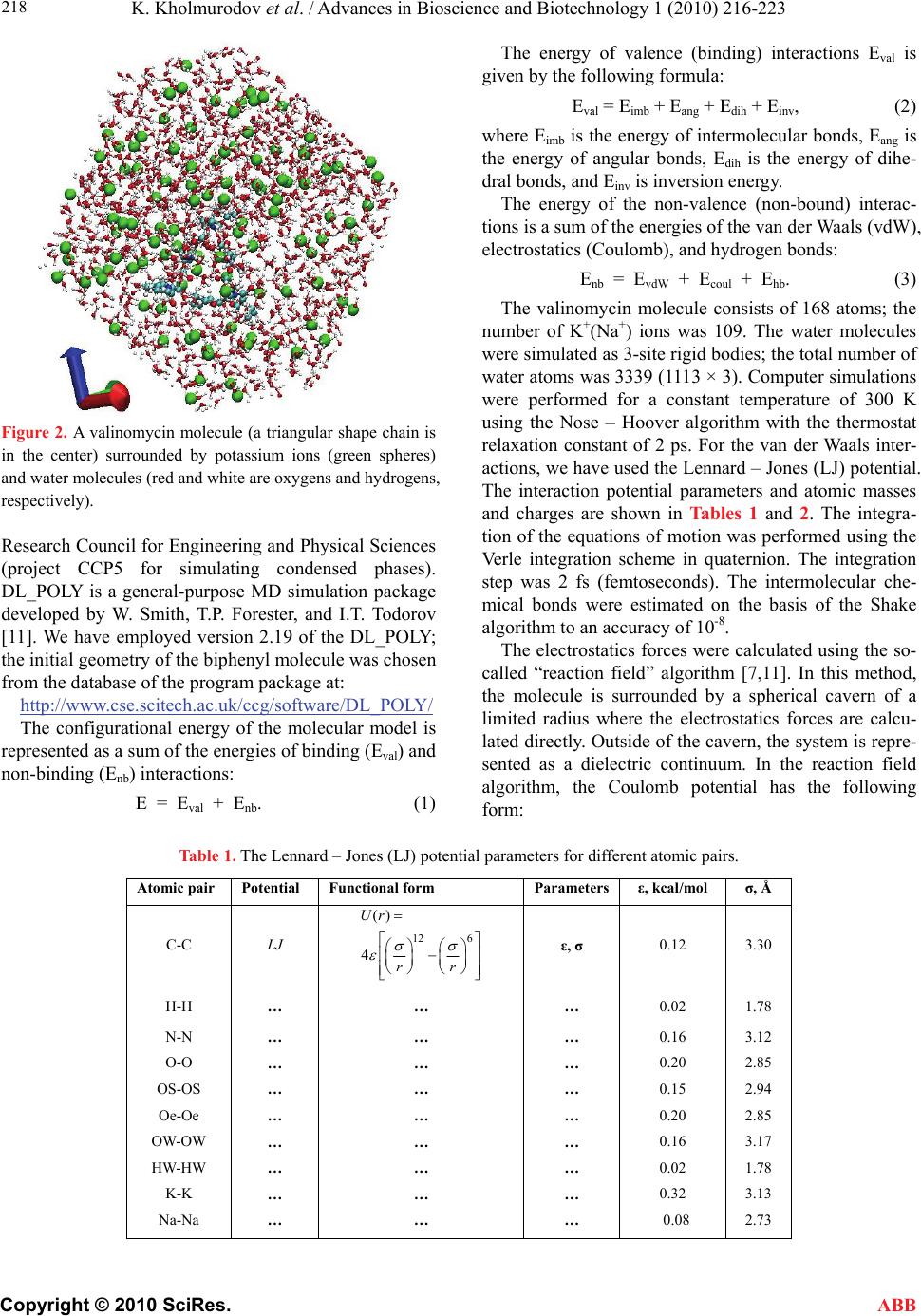
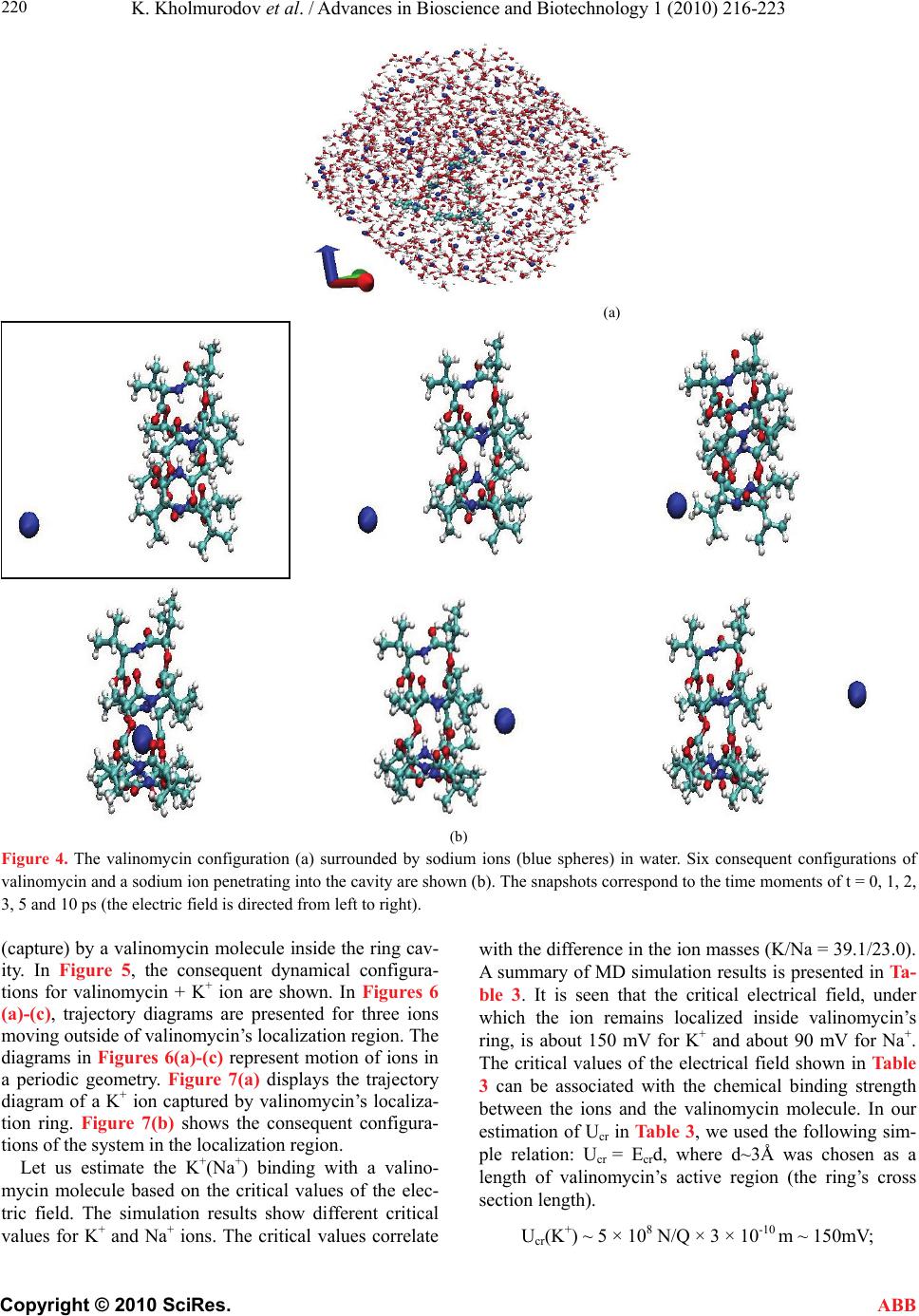
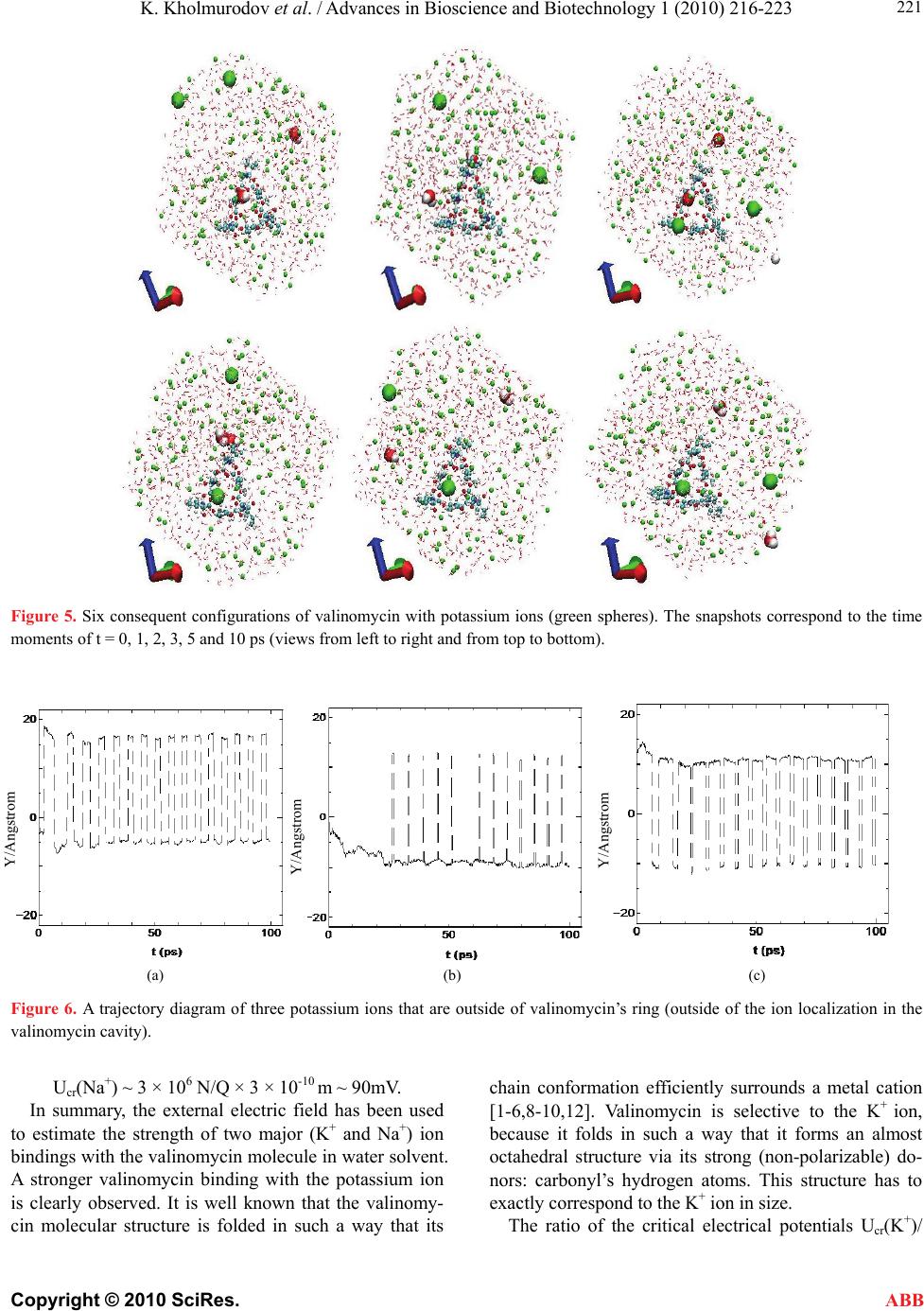
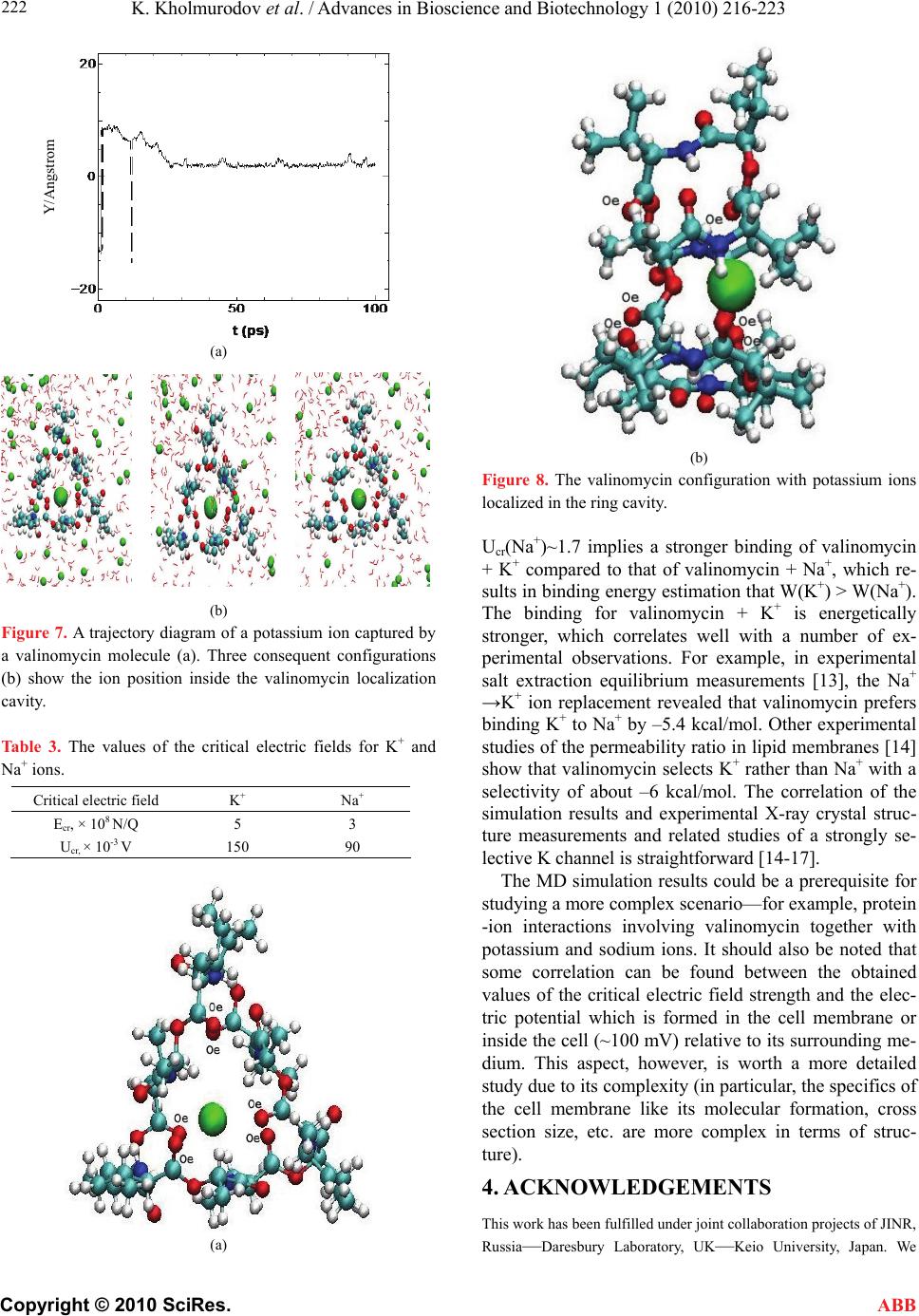
K. Kholmurodov et al. / Advances in Bioscience and Biotechnology 1 (2010) 216-223
Copyright © 2010 SciRes. ABB
223
would like to thank Prof. William Smith for the software support. This
work was supported in part by Grant in Aid for the Global Center of
Excellence Program for “Center for Education and Research of Sym-
biotic, Safe and Secure System Design” from the Ministry of Educa-
tion, Culture, Sport, and Technology in Japan.
REFERENCES
[1] Steed, I.W. and Atwood, J.L. (2000) Supramolecular
chemistry. Wiley.
[2] Doyle, D.A., Cabral, J.M., Pfuetzner, R.A., Kuo, A.,
Gulbis, J.M., Cohen, S.L., Chait, B.T. and MacKinnon, R.
(1998) The structure of the potassium channel: Molecular
basis of K+ conduction and selectivity. Science, 280
(5360), 69-77.
[3] Forester, T., Smith, W. and Clarke, H.R. (1997) Antibi-
otic activity of valinomycin. Journal of the Chemical So-
ciety Faraday Transactions, 93(4), 613-661.
[4] Ye, Q., Vincze, A., Horvai, G. and Leermakers, F.A.M.
(1998) Partial blocking of ion transport at the interface of
an ion-selective liquid membrane electrode by neutral
surfactants. Experiment and computer simulations. Elec-
trochimica Acta, 44(1), 125-132.
[5] Marrone, T.J. and Kenneth, M.M., Jr. (1995) Molecular
Recognition of K+ and Na+ by valinomycin in methanol.
Journal of American Chemical Society, 117(2), 779-791.
[6] Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K. and
Watson, Y.D. (1994) Molecular biology of the cell. Gar-
land Publishing, A member of the Taylor & Francis Group,
New York.
[7] Smith, W. and Forester, T. (1996) DL_POLY_2.0: A gen-
eral-purpose parallel molecular dynamics simulation
package. Journal of Molecular Graphics, 14(3), 136-141.
[8] Varma, S., Sabo, D. and Rempe, S.B. (2008) K+/Na+
selectivity in K-channels and valinomycin: Over-coor-
dination vs cavity-size constraints. Journal of Molecular
Biology, 376(1), 13-22.
[9] Forester, T.R., Smith, W. and Clarke, J.H.R. (1995) Cap-
ture of potassium ions by valinomycin: A molecular dy-
namics simulation study. Journal of Physical Chemistry,
99(39), 14418-14423.
[10] Eisenman, G., Alvarez, O. and Aqvist, J. (1992) Free
energy perturbation simulations of cation binding to
valinomycin. Journal of Inclusion Phenomena and Mac-
rocyclic Chemistry, 12(1-4), 23-53.
[11] Smith, W., Forester, T.R. and Todorov, I.T. (2008) The
DL poly 2 user manual. STFC Daresbury Laboratory
Daresbury, Warrington WA4 4AD Cheshire, Version
2.19.
[12] van Veggel, C.J.M.F., van Duynhoven, J.P.M., Harkema,
S., Wolbers, M.P.O. and Reinhoudt, D.N. (1996) Solid-
state structure, dynamical properties in solution and com-
putational studies of a new sodium hemispherand com-
ples. Journal of the Chemical Society Perkin Transac-
tions, 2, 449-454.
[13] Eisenman, G., Aqvist, J. and Alvarez, O. (1991) Free
energies underlying ion binding and transport in protein
channels: Free energy perturbation simulations of ion
binding and selectivity for valinomycin. Journal of the
Chemical Society Faraday Transactions, 87, 2099-2109.
[14] Eisenman, G. and Alvarez, O. (1992) Ionic selectivity of
proteins: Lessons from molecular dynamics simulations
on valinomycin. In: Gaber, B.P. and Easwaran, K.R.K.,
Eds., Biomembrane Structure and Function - The State of
the Art, Adenine Press, Schenectady, 321-351.
[15] Neilson, G.W., Mason, P.E., Ramos, S. and Sullivan, D.
(2001) Neutron and X-ray scattering studies of hydration
in aqueous solutions. Philosophical Transactions of the
Royal Society A, 359, 1575-1591.
[16] Shrivastava, I.H., Tieleman, D.P., Biggin, P.C. and San-
som, M.S.P. (2002) K(+) versus Na(+) ions in a K chan-
nel selectivity filter: A simulation study. Biophysical
Journal, 83(2), 633-645.
[17] Soper, A.K. and Weckstrom, K. (2006) Ion solvation and
water structure in potassium halide aqueous solutions.
Biophysical Chemistry, 124(3), 180-191; 39, 1023-1029.