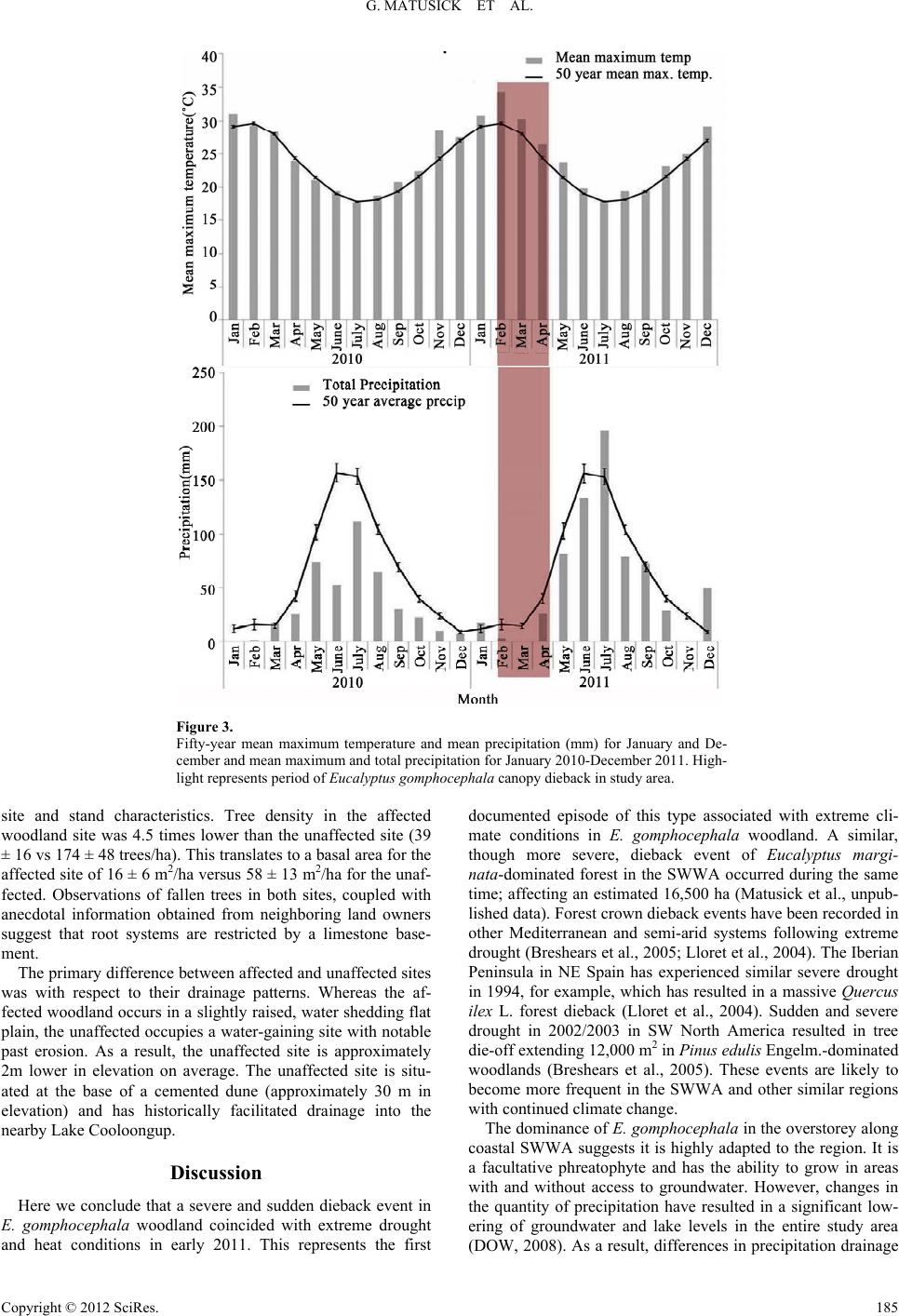
G. MATUSICK ET AL.
patterns between the affected (water-shedding) and unaffected
(water-gaining) may explain the strong contrast in woodland
canopy impacts. Trees in water-limited forest ecosystems are
highly dependent on soil type, slope, and drainage patterns
(Costa et al., 2008). We therefore hypothesize the cumulative
effects of long-term rainfall reductions (leading to dropping
groundwater levels), inadequate winter precipitation in 2010/
2011 (leading to inadequate soil recharge), and severe heat
conditions in February 2011(leading to high canopy water
stress) led to canopy dieback in the water-shedding E. gom-
phocephala woodland. During extreme periods of dryness,
eucalypts located on water-shedding sites, with low soil water
holding capacity are susceptible to rapid drying and dieback
(Pook et al., 1966). We suspect the neighboring unaffected E.
gomphocephala woodland, although transpiring more water,
receives higher rates of precipitation runoff and therefore is
capable of withstanding stressful summertime conditions.
Recent climate change modeling suggests that Mediterranean
ecosystems will exhibit a range of responses to future climate
changes (Klausmeyer & Shaw, 2009). Specifically, this model-
ing has predicted that E. gomphocephala-dominated ecosys-
tems from the study area north will contract, while areas south
of the study are expected to stay stable or expand (Klausmeyer
& Shaw, 2009). Observations from this study are able to con-
firm these modeled projections are correct for the study area.
Additionally, they provide important field evidence for how
future contractions in E. gomphocephala ecosystems may occur.
An accumulation of this type of data is required for field vali-
dation of modeling tools as well as understanding the cascade
of immediate, short- and long-term changes in ecosystem
structure and functioning following dieback (Ellison et al.,
2005). Ultimately, this information will be required to facilitate
mitigation strategies.
Acknowledgements
The authors would like to thank the West Australian State
Department of Environment and Conservation and the City of
Rockingham for their support.
REFERENCES
Bates, B. C., Hope, P., Ryan, B., Smith, I., & Charles, S. (2008). Key
findings from the Indian Ocean Climate Initiative and their impact on
policy development in Australia. Climate Change, 89, 339-354.
doi:10.1007/s10584-007-9390-9
Breshears, D. D., Cobb, N. S., Rich, P. M., Price, K. P., Allen, C. D.,
Balice, R. G., Romme, W. H., Kastens, J. H., Floyd, M. L., Belnap, J.,
Anderson, J. J., Myers, O. B., & Meyer, C. W. (2005). Regional
vegetation die-off in response to global-change-type drought. Pro-
ceedings of the National Academy of Sciences, 102, 15144-15148.
doi:10.1073/pnas.0505734102
Cai, Y. F., Barber, P., Dell, B., O’Brien, P., Williams, N., Bowen, B., &
Hardy, G. (2010). Soil bacterial functional diversity is associated
with the decline of Eucalyptus gomphocephala. Forest Ecology and
Management, 260, 1047- 1057. doi:10.1016/j.foreco.2010.06.029
Close, D. C., Davidson, N. J., Swanborough, P. W., & Corkrey, R.
(2011). Does low-intensity surface fire increase water- and nutri-
ent-availability to overstorey Eucalyptus gomphocephala? Plant Soil,
349, 203-214. doi:10.1007/s11104-011-0862-3
Commonweath of Australia, Bureau of Meteorology (2011). Perth in
2010: One of the hottest and driest years on record. URL (last
checked 19 September 2012).
http://www.bom.gov.au/climate/current/annual/wa/archive/2010.pert
h.html
Costa, A., Madeira, M., & Oliveira, A. C. (2008). The relationship
between cork oak growth patterns and soil, slope and drainage in a
cork oak woodland in Southern Portugal. Forest Ecology and Man-
agement, 255, 1525-1535. doi:10.1016/j.foreco.2007.11.008
Croton, J. T., & Reed, A. J. (2007). Hydrology and bauxite mining on
the Darling Plateau. Restoration Ecology, 15, S40-S47.
doi:10.1111/j.1526-100X.2007.00291.x
Curry, S. J. (1980). The association of insects with eucalypt dieback in
South Australia. In K. M. Old, G. A. Kile, & C. P. Ohmart (Eds.),
Proceedings of CSIRO conference on Eucalypt Dieback in Forests
and Woodlands, Canberra, 4-6 August 1980.
Damesin, C., Rambal, S., & Joffre, R. (1998). Co-occurrence of trees
with different leaf habit: A functional approach on Mediterranean
oaks. Acts Oecologia, 19, 195-204.
doi:10.1016/S1146-609X(98)80024-6
Department of Environment and Conservation, City of Rockingham
(2010). Proposed final management plan, Rockingham Lakes re-
gional park. Perth: Conservation Commission of Western Australia.
Department of Water, Government of Western Australia (2008). Rock-
ingham-Stakehill Groundwater Management Plan. Perth: Depart-
ment of Water, Government of Western Australia.
Edwards (2004). Environmental correlates and associations of tuart
(Eucalyptus gomphocephala DC.) decline. Master’s Thesis, Perth:
Edith Cowen University.
Ellison, A. M., Bank, M. S., Clinton, B. D., Colburn, E. A., Elliott, K.,
Ford, C. R., Foster, D. R., Kloeppel, B. D., Knoepp, J. D., Lov ett, H.
M., Mohan, J., Orig, D. A., Rodenhouse, N. L., Sobczak, W. V.,
Stinson, K. A., St one, J. K., Swan, C. M., Thompso n, J., Con Holle,
B., & Webster, J. R. (2005). Loss of foundation species: Conse-
quences for the structure and dynamics of forested ecosystems.
Frontiers in Ecology and the Environment, 3, 479-486.
doi:10.1890/1540-9295(2005)003[0479:LOFSCF]2.0.CO;2
Fox, J. E. D., & Curry, S. J. (1980). Notes on the tuart tree (Eucalyptus
gomphocephala) in the Perth area. Western Australian Naturalist, 14,
174-186.
Galiano, L., Martínez-Vilalta, J., Sabaté, S., & Lloret, F. (2012). De-
terminants of drought effects on crown condition and their relation-
ship with depletion of carbon reserves in a Mediterranean holm oak
forest. Tree Physiology, 32, 478-489.
doi:10.1093/treephys/tps025
Giorgi, F., & Lionello, P. (2008). Climate change projections for the
Mediterranean region. Global Planet Change, 63, 90-104.
doi:10.1016/j.gloplacha.2007.09.005
Jump, A., Hunt, J. M., & Peñuelas, J. (2006). Rapid climate change-
related growth decline at the southern edge of Fagus sylvatica.
Global Change Biology, 12, 2163-2174.
doi:10.1111/j.1365-2486.2006.01250.x
Klausmeyer, K. R., & Shaw, M. R. (2009). Climate change, habitat loss,
protected areas and the climate adaptation potential of species in
Mediterranean ecosystems worldwide. PLoS ONE, 4, e6392.
doi:10.1371/journal.pone.0006392
Lloret, F., Siscart, D., & Dalmases, C. (2004) Canopy recovery after
drought dieback in holm-oak Mediterranean forests of Catalonia (NE
Spain). Global Change Bio lo gy , 1, 2092-2099.
doi:10.1111/j.1365-2486.2004.00870.x
Mitchell, K. (2001). Quantitative analysis by the point-centered quarter
method. New York: Hobart and Will iam Smith Colleges.
Petrone, K. C., Hughes, J. D., Van Biel, T. G., & Silberstein, R. P.
(2010). Streamflow decline in southwestern Australia, 1950-2008.
Geophysical Research Letters, 37, L11401.
doi:10.1029/2010GL043102
Pook, E. W., Costin, A. B., & Moore, C. W. E. (1966). Water stress in
native vegetation during the drought of 1965. Australian Journal of
Botany, 14, 257-26 7. doi:10.1071/BT9660257
Sommer, B., & Froend, R. (2011). Resilience of phreatophytic vegeta-
tion to groundwater drawdown: is recovery possible under a drying
climate? Ecohydrology, 4, 67-82. doi:10.1002/eco.124
Copyright © 2012 SciRes.
186