G. Lee, B. Ge / Advances in Bioscience and Biotechnology 3 (2012) 679-685
684
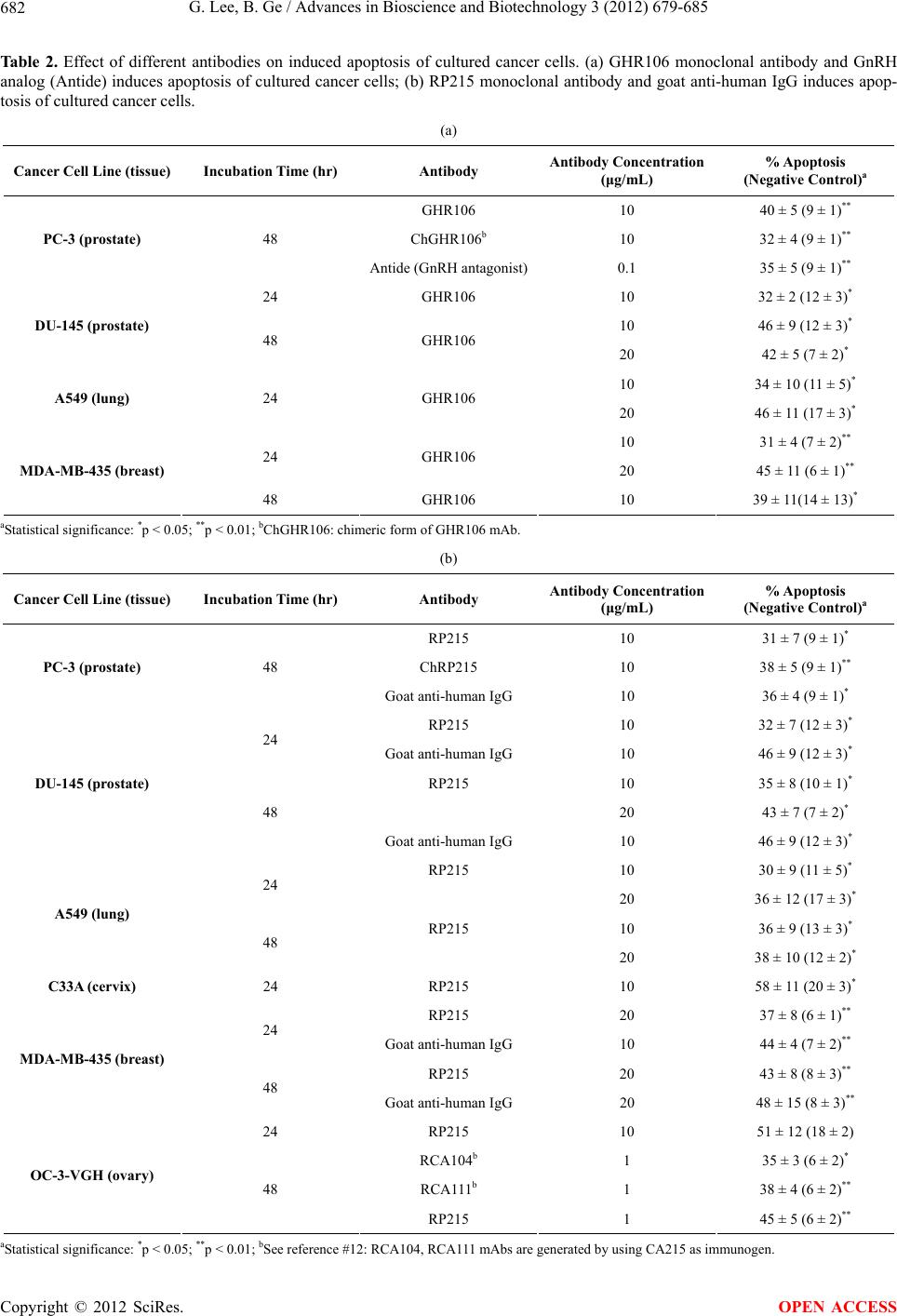
to assume that humanized forms of RP215 mAb may
have the potential to be developed as an effective anti-
body-based anti-cancer drug with multi-indication for
different types of cancer in humans.
5. CONCLUSION
GHR106 and RP215 mAbs have been evaluated, with
respect to their actions, to induce apoptosis on cultured
cancer cells, of which the GnRH/GnRH receptor and
RP215/CA215 systems are universally and abundantly
present. It remains to be shown if the efficacy of these
two mAbs in humanized forms can be demonstrated
through extensive clinical studies for cancer therapy in
humans in the near future.
6. ACKNOWLEDGEMENTS
This work was supported in parts by NSERC-IRAP of Canada. The
excellent technical support of Suefay Liu from McGill University and
Vivian Wang from the University of British Columbia is acknowledged.
REFERENCES
[1] Millar, R.P. (2005) GnRHs and GnRH receptors. Animal
Reproduction Science, 88, 5-28.
doi:10.1016/j.anireprosci.2005.05.032
[2] Harrison, G.S., Wierman, M.E., Nett, T.M. and Glode,
L.M. (2004) Gonadotropin-releasing hormone and its
receptor in normal and malignant cells. Endocrine-
Related Cancer, 11, 725-748.
doi:10.1677/erc.1.00777
[3] Cheng, C.K. and Leung P.C.K. (2005) Molecular biology
of gonadotropin-releasing hormone (GnRH)-I, GnRH-II,
and their receptors in humans. Endocrine Reviews, 26,
283-306. doi:10.1210/er.2003-0039
[4] Flanagan, C., Millar, R. and Illing, N. (1997) Advances in
understanding gonadotrophin-releasing hormone receptor
structure and ligand interactions. Reviews of Reproduction,
2, 113-120. doi:10.1530/ror.0.0020113
[5] Grundker, C. and Emons G. (2003) Role of gonadotropin-
releasing hormone (GnRH) in ovarian cancer. Reproduc-
tive Biology and Endocrinology, 1, 65.
http://www.RBEj.com/content/1/1/65
[6] Limonta, P., Moretti, R.M., Marelli, M.M. and Motta, M.
(2003) The biology of gonadotropin hormone-releasing
hormone: Role in the control of tumor growth and
progression in humans. Frontiers in Neuroendocrinology,
24, 279-295. doi:10.1016/j.yfrne.2003.doi:10.003
[7] Felberbaum, R.E., Ludwig, M. and Diedrich, K. (2000)
Clinical application of GnRH-antagonists. Molecular and
Cellular Endocrinology, 166, 9-14.
doi:10.1016/s0303-7207(00)00291-4
[8] Gründker, C., Völker, P., Schulz, K.-D. and Emons, G.
(2000) Luteinizing hormone-releasing hormone agonist
triptorelin and antagonist cetrorelix inhibit EGF-induced
c-fos expression in human gynecological cancers. Gyne-
cologic Oncology, 78, 194-202.
doi:10.1006/gyno.2000.5863
[9] Hong, I.-S., Cheung, A.P. and Leung, P.C.K. (2008) Gon-
adotropin-releasing hormones I and II induce apoptosis in
human granulosa cells. Journal of Clinical Endocrinology
and Metabolism, 93, 3179-3185.
doi:10.1210/jc.2008-0127
[10] Lee, C.-Y.G., Ho, J., Chow, S.-N., Yasojima, K., Schwab,
C. and McGeer, P.L. (2000) Immunoidentification of
gonadotropin releasing hormone receptor in human sperm,
pituitary and cancer cells. American Journal of Repro-
ductive Immunology, 44, 170-177.
doi:10.1111/j.8755-8920.2000.440307.x
[11] Lee, G. and Ge, B. (2010) Growth inhibition of tumor
cells in vitro by using monoclonal antibodies against
gonadotropin-releasing hormone receptor. Cancer Immu-
nology, Immunotherapy, 59, 1011-1019.
doi:10.1007/s00262-010-0823-3
[12] Lee, G., Zhu, M. and Ge, B. (2012). Potential monoclonal
antibody therapy for the treatment of ovarian cancer. In:
Farghaly, S.A., Ed., Ovarian Cancer—Basic Science Per-
spective, InTech, Vancouver, 385-406.
[13] Lee, G., Cheung, A., Ge, B., Zhu, M., Giolma, B., Li, B.,
et al. (2012) CA215 and GnRH receptor as targets for
cancer therapy. Cancer Immunology, Immunotherapy, 61,
1805-1817. doi:10.1007/s00262-012-1230-8
[14] Chen, Z., Qiu, X. and Gu, J. (2009) Immunoglobulin
expression in non-lymphoid lineage and neoplastic cells.
American Journal of Pathology, 174, 1139-1148.
doi:10.2353/ajpath.2009.080879
[15] Hu, D., Zheng, H., Liu, H., Li, M., Ren, W., Liao, W., et
al. (2008) Immunoglobulin expression and its biological
significance in cancer cells. Cellular and Molecular
Immunology, 5, 319-324. doi:10.1038/cmi.2008.39
[16] Qiu, X., Zhu, X., Zhang, L., Mao, Y., Zhang, J., Hao, P.,
et al. (2003) Human epithelial cancers secrete immu-
noglobulin G with unidentified specificity to promote
growth and survival of tumor cells. Cancer Research, 63,
6488-6495.
http://cancerres.aacrjournals.org/content/63/19/6488.abstr
act
[17] Lee, C.Y., Chen, K.W., Sheu, F.S., Tsang, A., Chao, K.C.
and Ng, H.T. (1992) Studies of a tumor-associated
antigen, COX-1, recognized by a monoclonal antibody.
Cancer Immunology, Immunotherapy, 35, 19-26.
doi:10.1007/BF01741050
[18] Lee, G., Laflamme, E., Chien, C.-H. and Ting, H.H.
(2008) Molecular identity of a pan cancer marker, CA215.
Cancer Biology and Therapy, 7, 2007-2014.
doi:10.4161/cbt.7.12.6984
[19] Lee, G. and Ge, B. (2009) Cancer cell expressions of
immunoglobulin heavy chains with unique carbohydrate-
associated biomarker. Cancer Biomarkers, 5, 177-188.
doi:10.3233/cbm-2009-0102
[20] Lee, G., Zhu, M., Ge, B. and Potzold, S. (2012) Wide-
spread expressions of immunoglobulin superfamily
proteins in cancer cells. Cancer Immunology, Immuno-
therapy, 61, 89-99. doi:10.1007/s00262-011-1088-1
[21] Lee, G. and Ge B. (2010) Inhibition of in vitro tumor cell
Copyright © 2012 SciRes. OPEN ACCESS