 Advances in Microbiology, 2012, 2, 295-302 http://dx.doi.org/10.4236/aim.2012.23035 Published Online September 2012 (http://www.SciRP.org/journal/aim) Growth and Antagonism of Trichoderma spp. and Conifer Pathogen Heterobasidion annosum s.l. in Vitr o at Different Temperatures Vizma Nikolajeva1, Zaiga Petrina1, Livija Vulfa1, Laura Alksne1, Daina Eze1, Lelde Grantina1, Talis Gaitnieks2, Anita Lielpetere1 1Faculty of Biology, University of Latvia, Riga, Latvia 2Latvian State Forest Research Institute “Silava”, Salaspils, Latvia Email: vizma.nikolajeva@lu.lv Received April 15, 2012; revised May 18, 2012; accepted June 1, 2012 ABSTRACT Variations in the radial growth rate of 24 isolates belonging to ten species of Trichoderma, three isolates of conifer pathogen Heterobasidion anno sum s.s. and four isolates of H. parviporum were evaluated by incubation on a solid malt extract medium at a temperature of 4˚C, 15˚C and 21˚C. Trichoderma antagonism against Heterobasidion was investi- gated in dual culture in vitro. The slowest rate of growth was referable to all seven strains of Heterobasidion spp. All Heterobasidion spp. strains were overgrown by 63% of Trichoderma spp. strains after two weeks at 21˚C and by 33% of strains at 15˚C. 21% of Trichoderma strains did not grow and only four strains belonging to T. koningii, T. viride and T. viridescens demonstrated the ability to completely overgrow Heterobasidion spp. after two weeks incubation at 4˚C. According to the antagonistic efficiency, Trichoderma strains were divided into five groups with an Euclidean distance of 25. The groups contained isolates from different species. It was suggested that selected psychrotrophic fast growing T. viride, T. koningii and T. viridescens strains could be examined in different substrate conditions as suitable antagonist agents for the control of H. annos u m and H. par vip oru m. Keywords: Trichoderma; Heter ob a s idi on ; Antagonism; Growth Rate; Temperature 1. Introduction Conifer root and butt rot disease caused by Heterobasid- ion annosum (Fr.) Bref. sensu lato is an economically important disease of coniferous trees in boreal and tem- perate forests [1]. The fungus spreads through aerial basidiospores to stump surfaces and wounds, and by mycelia via root contacts from tree to tree [2,3]. Hetero- basidion spp. also has an anamorph stage preferably de- veloping in laboratory conditions and forming conidia. Conidiospores also form on the stumps in moist weather and can survive in soil up to ten months [4]. Hetero- basidion spp. grow at a wide range of temperature with a minimum of 2˚C [5]. A biocontrol method to reduce H. annosum s.l. infec- tion is to apply the wood degrading fungus Phlebiopsis gigantea in a spore suspension directly on the freshly cut stumps immediately after cutting [6]. There are also a lot of other fungi examined for Heterobasidion biocontrol potential including Trichoderma spp. [7]. Trichoderma and its teleomorphic stage Hypocrea in- clude cosmopolitan soil-borne species. At present, the International Subcommission on Trichoderma and Hypo- crea Taxonomy lists 104 species (http://www.isth.info). Trichoderma are particularly prevalent in the humic layer of hardwood forests where they represent up to 3% of all fungal propagules [8]. Some strains are commonly used to control different plant diseases. Possible mechanisms involved in disease control by Trichoderma include my- coparasitism, antibiosis, competition, and induction of plant defence responses [9]. Trichoderma are also wood- colonising fungi which restrict the growth of H. annosum in vitro and under natural conditions [10,11]. There are a number of other investigations of antagonistic action of Trichoderma against Heterobasidion [12-16]. However, only one or a few strains per species of Trichoderma were involved [7]. In general, Trichoderma species are mesophylic. How- ever, they possess different temperature minima, optima and maxima. Temperature also affects the metabolism features and antagonistic properties of Trichoderma spp. [17]. Goldfarb et al. [18] ascertained variation of growth rate substantially within and among species. The investi- gations supported the opinion that Trichoderma isolates C opyright © 2012 SciRes. AiM 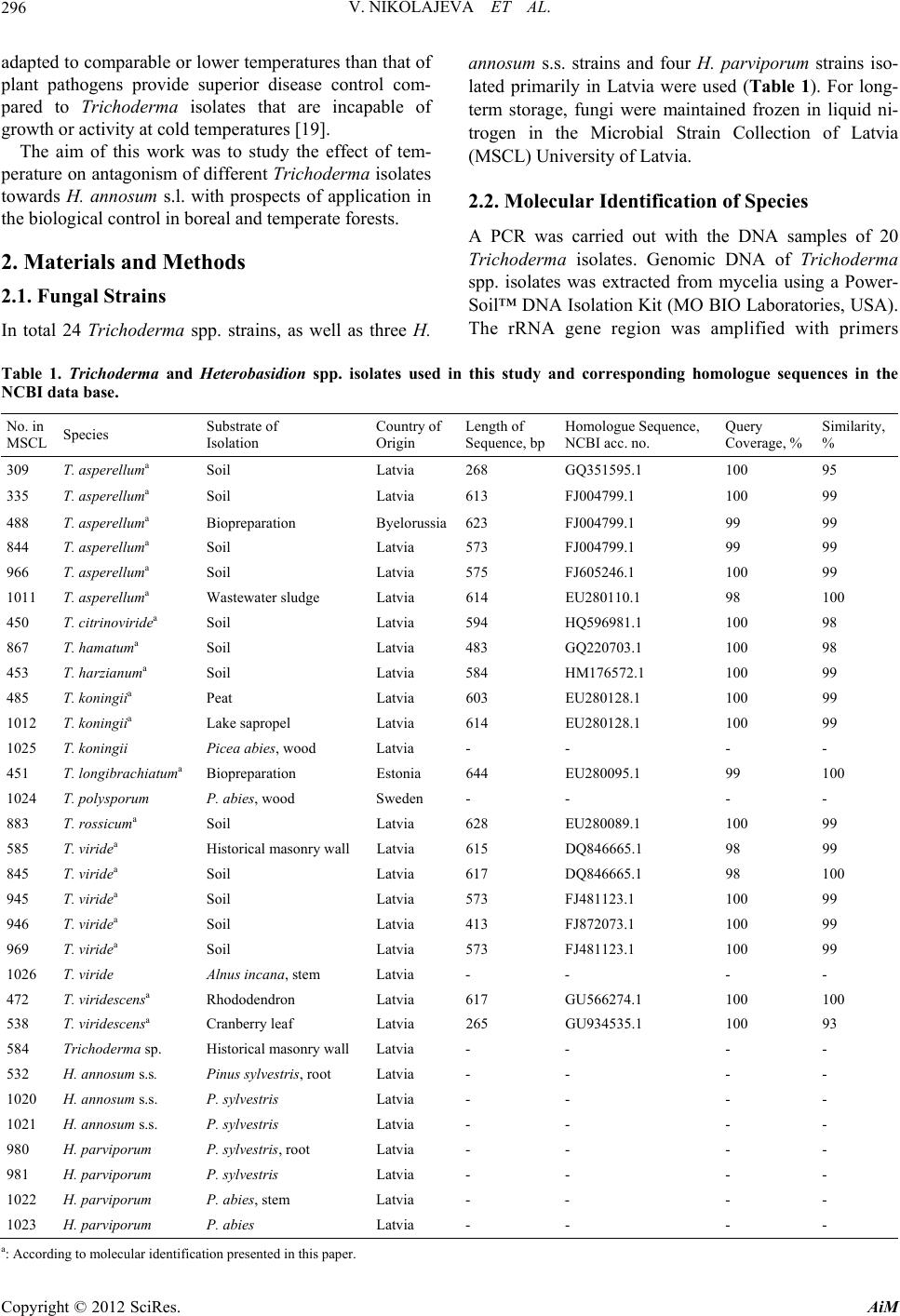 V. NIKOLAJEVA ET AL. 296 adapted to comparable or lower temperatures than that of plant pathogens provide superior disease control com- pared to Trichoderma isolates that are incapable of growth or activity at cold temperatures [19]. The aim of this work was to study the effect of tem- perature on antagonism of different Trichoderma isolates towards H. annosum s.l. with prospects of application in the biological control in boreal and temperate forests. 2. Materials and Methods 2.1. Fungal Strains In total 24 Trichoderma spp. strains, as well as three H. annosum s.s. strains and four H. parviporum strains iso- lated primarily in Latvia were used (Table 1). For long- term storage, fungi were maintained frozen in liquid ni- trogen in the Microbial Strain Collection of Latvia (MSCL) University of Latvia. 2.2. Molecular Identification of Species A PCR was carried out with the DNA samples of 20 Trichoderma isolates. Genomic DNA of Trichoderma spp. isolates was extracted from mycelia using a Power- Soil™ DNA Isolation Kit (MO BIO Laboratories, USA). The rRNA gene region was amplified with primers Table 1. Trichoderma and Heterobasidion spp. isolates used in this study and corresponding homologue sequences in the NCBI data base. No. in MSCL Species Substrate of Isolation Country of Origin Length of Sequence, bp Homologue Sequence, NCBI acc. no. Query Coverage, % Similarity, % 309 T. asperelluma Soil Latvia 268 GQ351595.1 100 95 335 T. asperelluma Soil Latvia 613 FJ004799.1 100 99 488 T. asperelluma Biopreparation Byelorussia623 FJ004799.1 99 99 844 T. asperelluma Soil Latvia 573 FJ004799.1 99 99 966 T. asperelluma Soil Latvia 575 FJ605246.1 100 99 1011 T. asperelluma Wastewater sludge Latvia 614 EU280110.1 98 100 450 T. citrinoviridea Soil Latvia 594 HQ596981.1 100 98 867 T. hamatuma Soil Latvia 483 GQ220703.1 100 98 453 T. harzianuma Soil Latvia 584 HM176572.1 100 99 485 T. koningiia Peat Latvia 603 EU280128.1 100 99 1012 T. koningiia Lake sapropel Latvia 614 EU280128.1 100 99 1025 T. koningii Picea abies, wood Latvia - - - - 451 T. longibrachiatuma Biopreparation Estonia 644 EU280095.1 99 100 1024 T. polysporum P. abies, wood Sweden - - - - 883 T. rossicuma Soil Latvia 628 EU280089.1 100 99 585 T. viridea Historical masonry wall Latvia 615 DQ846665.1 98 99 845 T. viridea Soil Latvia 617 DQ846665.1 98 100 945 T. viridea Soil Latvia 573 FJ481123.1 100 99 946 T. viridea Soil Latvia 413 FJ872073.1 100 99 969 T. viridea Soil Latvia 573 FJ481123.1 100 99 1026 T. viride Alnus incana, stem Latvia - - - - 472 T. viridescensa Rhododendron Latvia 617 GU566274.1 100 100 538 T. viridescensa Cranberry leaf Latvia 265 GU934535.1 100 93 584 Trichoderma sp. Historical masonry wall Latvia - - - - 532 H. annosum s.s. Pinus sylvestris, root Latvia - - - - 1020 H. annosum s.s. P. sylvestris Latvia - - - - 1021 H. annosum s.s. P. sylvestris Latvia - - - - 980 H. parviporum P. sylvestris, root Latvia - - - - 981 H. parviporum P. sylvestris Latvia - - - - 1022 H. parviporum P. abies, stem Latvia - - - - 1023 H. parviporum P. abies Latvia - - - - a: Ac cording to molecular identification presented in this paper. Copyright © 2012 SciRes. AiM  V. NIKOLAJEVA ET AL. 297 ITS1F [20] and ITS4 [21]. The reactions in Eppendorf Mastercycler Personal were carried out in 25 μl volume. The mixture contained 0.2 μl Hot Start Taq DNA Polymerase, 2.5 μl 10X Hot Start PCR Buffer, 2.5 μl dNTP Mix, 2 mM each, 2 μl 25 mM MgCl2 (all reagents from Fermentas, Lithuania), 0.5 μl of each 25 μM primer (OPERON Biotechnologies), 15.43 μl sterile distilled water and 1 μl of DNA template. The PCR conditions were as follows: the initial denaturation step of 4 min at 95˚C, 40 s of denaturation at 95˚C, 40 s of annealing at 52˚C, 1 min of primer extension at 72˚C (30 cycles) and final extension 10 min at 72˚C. PCR products (20 - 25 µl) were purified using 0.5 μl Exonuclease I (20 u·μl–1) and 2.0 µL Shrimp Alkaline Phosphatase (1 u·μl–1) (both from Fermentas, Lithuania). Samples were further pro- cessed in a PCR machine 30 min at 37˚C, 15 min at 85˚C. Both strands of the amplification products were sequenced by using an ABI Prism Big Dye terminator cycle sequencing ready reaction kit (version 3.1, Applied Biosystems, Foster City, USA) and primers ITS1F and ITS4. Sequencing of strains 309 - 845 and 1011 - 1012 was done in CBS, Utrecht, the Netherlands. Sequencing of strains 867 - 969 was done in the Biomedical Study and Research Center at the Univer- sity of Latvia. Obtained sequences were analyzed using Staden Package 1.6.0. release. The resulting consensus sequences were used in the BLASTN homology search against The National Center for Biotechnology Infor- mation (NCBI) nucleotide database (http://www.ncbi.nlm.nih.gov). 2.3. Growth Kinetics and Antagonism Assay Paired cultures of Trichoderma and Heterobasidion spp. were grown from inocula placed 5 cm apart, on malt ex- tract agar (MEA; Becton Dickinson, USA) at 4˚C, 15˚C and 21˚C. In controls, single cultures were grown in the centre of Petri dishes. Radii of fungal colonies have been recorded daily after 3rd - 28th days. The rate of colony growth was calculated from measurements of radii after three days long incubation of fungi at 21˚C and 15˚C or after 10 days of incubation at 4˚C when the growth cor- responded to the exponential growth phase. The results were expressed in mm·h–1. The experiment was per- formed twice. The efficiency of Trichoderma in sup- pressing radial growth was calculated as follows: 100CTC, where C is radial growth measurement of the pathogen in the control and T is radial growth of the pathogen in the presence of Trichoderma [22]. 2.4. Conidiospore Germination Conidia from H. annosum 1020 were obtained from fully sporulated agar cultures. Droplets (100 l) of spore sus- pension (about 107 conidia·ml–1) were transferred to ster- ile cellophane strips and placed onto a surface of Tri- choderma spp. growing in the MEA medium for three days. Germinated spores were counted after one, two and three days of incubation at 21˚C using a microscope. Germination of 100 randomly selected conidia in each droplet was evaluated with a microscope at ×600 mag- nification. Enumeration of germinated conidia was per- formed in five replicates (on five cellophane strips). A conidium was considered germinated, if the length of the germ tube was not shorter than the diameter of the co- nidium. The experiment was repeated three times. An average of measurements from 15 replicates was calcu- lated. 2.5. Statistical Analysis The significance of the difference in values was deter- mined through ANOVA analysis at a significance level of 0.05. The efficiency of different Trichoderma strains in suppressing of Heterobasidion spp. was analysed us- ing hierarchial cluster analysis with complete linkage method and Euclidean distance matrix. 3. Results 3.1. Identity of Trichoderma Species According to molecular identification, six Trichoderma strains proved to be T. asperellum and five belonged to T. viride, two strains belonged to T. koningii and two be- longed to T. viridescens but five further species (T. citri- noviride, T. hamatum, T. harzianum, T. longibrachiatum and T. rossicum) were represented by single isolates (Table 1). 3.2. Growth and Antagonism between Trichoderma and Heterobasidion spp., in Vitro 17% of the investigated Trichoderma spp. strains grew at all the temperatures tested (4˚C, 15˚C, and 21˚C), but the growth was comparatively slow at 4˚C. The observed mean radial growth rate was from less than 0.010 (five Trichoderma spp. strains) to 0.139 (T. viride 969) mm·h–1 for Trichoderma spp. incubated at 4˚C, from 0.094 (Trichoderma sp. 584) to 0.260 (T. viride 1026) mm·h–1 at 15˚C and from 0.187 (T. rossicum 883) to 0.451 (T. koningii 1025) mm·h–1 at 21˚C. The growth rate of Het- erobasidion strains was slower and ranged only from less than 0.010 (H. annosum 1020 and 1021) to 0.021 (H. parviporum 1023), from 0.017 (H. annosum 1020) to 0.045 (H. parviporum 1022) and from 0.061 (H. anno- sum 1020) to 0.133 (H. parviporum 981) correspondingly at 4˚C, 15˚C and 21˚C. Examining the differences among the species (Figure 1, able 2), we can see that the slowest rate of growth T Copyright © 2012 SciRes. AiM  V. NIKOLAJEVA ET AL. 298 0 10 20 30 40 50 60 358 1012151719 Time, days Radiuss, mm T.472 T.945 T.946 T.969 T.1025 T.1026 H.532 H.980 H.1020 H.1021 H.1022 H.1023 Figure 1. Mycelial radii of Trichoderma (T.) and Heterobasidion (H.) colonies during incubation on MEA at 4˚C. Isolate numbers are shown in Table 1. Data are presented as means for each variant. Error bars indicate SD. Table 2. Radial growth rate of colonies of Trichoderma and Heterobasidion species on MEA at different temperatures. Rate of Growth, mm·h–1 ±SD Species Number of Strains 21˚C 15˚C 4˚C T. asperellum 6 0.269 ± 0.015c 0.158 ± 0.017d,e 0.016 ± 0.026a,b T. citrinoviride 1 0.326 ± 0.008d 0.155 ± 0.008d 0.055 ± 0.006d T. hamatum 1 0.293 ± 0.009c 0.181 ± 0.010e 0.039 ± 0.005c T. harzianum 2 0.274 ± 0.011c 0.152 ± 0.027c,d,e 0.014 ± 0.014a,b T. koningii 3 0.344 ± 0.077c,d 0.194 ± 0.040d,e 0.080 ± 0.015e T. longibrachiatum 1 0.284 ± 0.005c 0.185 ± 0.009e <0.010a T. polysporum 1 0.194 ± 0.005b 0.117 ± 0.006b 0.055 ± 0.008d T. rossicum 1 0.187 ± 0.007b 0.138 ± 0.005c 0.064 ± 0.006d T. viride 6 0.338 ± 0.039d 0.219 ± 0.031e 0.080 ± 0.009e T. viridescens 2 0.284 ± 0.014c 0.184 ± 0.009e 0.073 ± 0.032c,d,e H. annosum s.s. 3 0.085 ± 0.019a 0.033 ± 0.012a 0.009 ± 0.008a H. parviporum 4 0.110 ± 0.014a 0.041 ± 0.003a 0.018 ± 0.005b Data are presented as means for all strains of each species at each temperature. Significant difference (P < 0.05) is established in all of the rows. The means in the same column with different superscripts are significantly (P < 0.05) different. was referable to all seven strains of two Heterobasidion species. Only some strains formed a sterile zone between antagonistic colonies. For example, it was in the case of T. asperellum 1011 and H. parviporum 981 (Figure 2). We also observed changes in the Trichoderma pigmenta- tion from green to yellow near of the pathogen while the pathogen colony restricted their growth. No inhibition zone formed, for example, between colonies of T. rossi- cum 883 and H. parviporum 981 (Figures 3(a) and (b)) at any incubation temperature. Heterobasidion spp. strains were overgrown (100% efficiency of Trichoderma) by 63% of Trichoderma spp. Copyright © 2012 SciRes. AiM  V. NIKOLAJEVA ET AL. 299 (a) (b) (b) Figure 2. Dual growth of T. asperellum 1011 (on the left) and H. parviporum 981 (on the right) after one week (a), two weeks (b) and four weeks (c) incubation at 15˚C. (a) (b) (b) Figure 3. Dual growth of T. rossicum 883 (on the left) and H. parviporum 981 (on the right) after one week (a) and four weeks (b) incubation at 21˚C, and (c) T. asperellum 309 (on the left) and H. parviporum 981 (on the right) after four weeks incubation at 4˚C. strains after two weeks at 21˚C and by 33% of strains at 15˚C (Table 3). The restriction of Heterobasidion was especially difficult and slow at 4˚C. 21% of Trichoderma strains did not grow and only four strains (T. koningii 585, T. viride 595 and 969, and T. viridescens 472) clus- tered in the group (Figure 4) what demonstrated the abil- ity to completely overgrow Heterobasidion spp. also after two weeks incubation at 4˚C. T. asperellum 309 was not able to grow at 4 ˚C and allowed to extend H. parvi- porum 981 (Figure 3(c)) with minimal restriction. 3.3. Trichoderma Impact on the Germination of Heterobasidion Species All eight of the randomly chosen Trichoderma spp. strains (T. koningii 1025, T. polysporum 1024, T. viride 585, 945, 946, 969 and 1026, and T. viridescens 472) caused inhibition of H. annosum 1020 conidia germina- tion after one day of incubation (data not shown). After two days, the inhibition effect was retained by only four T. viride strains and T. polysporum 1024. Both T. viride 585 and T. viride 969 demonstrated the ability to signifi- cantly (P < 0.05) slowing down the conidiospore germi- nation of pathogen (Ta ble 4), especially on the first day. Table 3. In vitro efficiency of Trichoderma isolates in suppressing linear growth of Heterobasidion spp. in paired cultures measured after two weeks of growth at different temperatures. Strain of Trichoderma 21˚C 15˚C 4˚C T. asperellum 309 72.2 ± 9.0 66.4 ± 12.2 5.0 ± 5.0 T. asperellum 335 77.5 ± 3.8 62.7 ± 11.4 0.0 ± 0.0 T. asperellum 488 74.6 ± 10.2 66.9 ± 10.2 0.0 ± 0.0 T. asperellum 844 88.2 ± 10.2 69.9 ± 8.3 0.0 ± 0.0 T. asperellum 966 100.0 ± 0.0 90.3 ± 5.3 47.7 ± 5.1 T. asperellum 1011 76.8 ± 11.2 60.8 ± 11.5 0.0 ± 0.0 T. citrinoviride 450 100.0 ± 0.0 81.9 ± 5.5 29.3 ± 11.8 T. hamatum 867 90.5 ± 3.8 80.4 ± 10.2 21.7 ± 4.7 T. harzianum 453 100.0 ± 0.0 75.4 ± 7.9 12.8 ± 10.8 T. koningii 485 100.0 ± 0.0 100.0 ± 0.0 100.0 ± 0.0 T. koningii 1012 100.0 ± 0.0 100.0 ± 0.0 47.3 ± 9.4 T. koningii 1025 100.0 ± 0.0 100.0 ± 0.0 55.2 ± 9.3 T. longibrachiatum 451 78.1 ± 6.4 68.8 ± 7.2 0.0 ± 0.0 T. polysporum 1024 66.7 ± 11.5 55.6 ± 5.9 34.9 ± 3.7 T. rossicum 883 55.9 ± 12.4 43.7 ± 6.8 32.1 ± 8.2 T. viride 585 100.0 ± 0.0 100.0 ± 0.0 100.0 ± 0.0 T. viride 845 100.0 ± 0.0 85.9 ± 8.1 47.2 ± 1.5 T. viride 945 100.0 ± 0.0 100.0 ± 0.0 45.6 ± 14.3 T. viride 946 100.0 ± 0.0 94.7 ± 4.1 51.7 ± 2.4 T. viride 969 100.0 ± 0.0 100.0 ± 0.0 100.0 ± 0.0 T. viride 1026 100.0 ± 0.0 100.0 ± 0.0 36.7 ± 11.6 T. viridescens 472 100.0 ± 0.0 100.0 ± 0.0 100.0 ± 0.0 T. viridescens 538 100.0 ± 0.0 95.3 ± 4.1 9.6 ± 4.0 Trichoderma sp. 584 100.0 ± 0.0 81.9 ± 8.5 7.0 ± 6.3 Data are presented as means of efficiency according to three Heterobasidion strains (H. 521, H. 980 and H. 981) SD. The efficiency of Trichoderma was calculated as follows: 100CTC , where C is radial growth measurement of the pathogen in the control and T is radial growth of the pathogen in the resence of Trichoderma. p Copyright © 2012 SciRes. AiM  V. NIKOLAJEVA ET AL. 300 Figure 4. Dendrogram, based on the Euclidean distance, of the Trichoderma strains according to efficiency in sup- pressing linear growth of Heterobasidion spp. in paired cul- tures at a temperature of 21˚C, 15˚C and 4˚C (Table 3). Table 4. Percentage of germinated H. annosum 1020 conidia ± SD affected by Trichoderma colonies. Time, Days Variant 1 2 3 Control 31.1 ± 1.1 ND ND T. viride 585 1.1 ± 0.8a 10.0 ± 3.8a ND T. viride 969 1.7 ± 2.5a 10.9 ± 1.0a 22.8 ± 3.6a aSignificant difference in comparison with control (P < 0.05). ND—the mycelium was formed and therefore the enumeration of germinated conidia was not possible. 4. Discussion 4.1. Growth of Fungi at Different Temperatures Our studies were carried out to determine the biocontrol potential of different Trichoderma strains against H. an- nosum and H. parviporum in vitro at different tempera- tures with emphasis on psychrotrophic growth at low temperature. It is known that the intensive infection of conifers with root rot occurs when the mean day tem- perature exceeds 5˚C because then conditions are suitable for germination of Heterobasidion basidiospores [2]. An optimum for H. annosum is located at 17˚C and 22˚C and for H. parviporum at 27˚C. The slowest growth takes place at 2˚C for both species (0.65 mm·d–1 for H. parvi- porum and 0.60 mm·d–1 for H. annosum). Our absolute values always were about 30% - 40% lower (Figure 1) and the obtained means differed significantly (P < 0.05) between species only at 4˚C (Table 2). The significant influence of the temperature (P < 0.05) was estimated on the growth rate of different Tricho- derma strains and species allowing their division into three, four and five groups respectively at a temperature of 21˚C, 15˚C and 4˚C (Table 2). Already Danielson and Davey [23] recognized that some Trichoderma species are typical for cool geographic regions (T. viride, T. polysporum) and others are typical for warm climatic regions (T. koningii, T. hamatum, T. harzianum, T. pseudokoningii, T. saturnisporum). Today, T. polyspo- rum is identified as one of the psychrotrophic Tricho- derma species present in temperate [18] and arctic envi- ronments [24]. Our fungal strains were isolated from the temperate region mainly from Latvia (Table 1). In addi- tion, our single T. polysporum strain (origin—Sweden) showed a high rate of growth (0.055 mm·h–1) at 4˚C. However, three other species, T. viride, T. koningii and T. viridescens, demonstrated a growth rate of 0.073 - 0.080 mm·h–1 and overcame T. polysporum (Table 2). Jaklitsch et al. [25] estimated that T. viride and T. viri- descens is characterised by north- and south-temperate distribution, relatively slow growth and temperature op- timum of about 25˚C. They showed very little variation in growth rate among their many isolates that corre- sponds to the results of our study. Lieckfeldt et al. [26] found that T. viride has an optimum temperature of 22.5˚C but T. asperellum has an optimum of 30˚C. We did not attempt to estimate optimum temperature, how- ever, all six of our investigated T. viride strains showed significantly faster growth (P < 0.05) than all six of T. asperellum strains at all of the used temperature regimes. 4.2. Fungal Antagonism at Different Temperatures The interaction between Trichoderma and Heterobasid- ion spp. was highly dependent on temperature. It is rec- ognized that different Trichoderma spp. strains have dif- ferent thermal requirements for biocontrol [27,28]. Köhl and Schlösser [29] established the potential of some Trichoderma isolates to decay sclerotia of Botrytis cine- rea at low temperatures, even at 5˚C. In the present in- vestigation we found out that all 24 of the investigated Trichoderma strains showed antagonistic activity against Heterobasidion. The number of Trichoderma strains over- growing pathogens decreased with a decrease in tem- perature from 21˚C to 15˚C and especially to 4˚C (Table 3). According to the antagonistic efficiency, Tricho- Copyright © 2012 SciRes. AiM  V. NIKOLAJEVA ET AL. 301 derma strains were divided into five groups with an Euclidean distance of 25 (Figure 4). The groups con- tained isolates from different species. We did not find any Trichoderma strain that demon- strated a significant difference in antagonism against several strains of Heterobasidion. This is in contrary to the investigation of Bell et al. [30], which showed that a single isolate of the antagonist can be highly effective against one isolate (in particular Rhizoctonia solani) but may have only minimal effects on other isolates of the same species. Napierala-Filipiak and Werner [31], who studied interactions between 42 mycorrhizal fungi and six strains of Heterobasidion spp., ascertained that al- though antagonism is dependent on growth rates of fungi, the fungi do not display antagonistic properties to all strains of the pathogen. Obtained results show that Trichoderma strains inhib- ited germination of Heterobasidion especially in the be- ginning (Table 4). A direct influence of hyphae and mac- romolecular compounds on the conidia was excluded by plating cellophane strips onto the surface of the colonies. Therefore, we can assume that soluble low molecular and/or volatile substances produced this impact. Possible mechanisms of antagonism among our iso- lates of Trichoderma and Heterobasidion such as myco- parasitism and antibiosis will be studied in our further experiments. The experiments will also include investi- gation of the ability of Trichoderma to replace Hetero- basidion in different substrate conditions including wood media. It was suggested that selected psychrotrophic fast growing T. viride, T. koningii and T. viridescens strains could be examined in different substrate conditions as suitable antagonist agents for the control of H. annosum and H. parvipor um . 5. Acknowledgements We would like to thank Centraalbureau voor Schimmel- cultures (Utrecht, The Netherlands) and the Biomedical Study and Research Center of the University of Latvia for sequencing of fungal strains. This study was sup- ported by the European Regional Development Fund (ERDF) (2010/0277/2DP/2.1.1.1.0/10/APIA/VIAA/129). REFERENCES [1] F. O. Asiegbu, A. Adomas and J. Stenlid, “Conifer Root and Butt Rot Caused by Heterobasidion annosum (Fr.) Bref. s.l.,” Molecular Plant Pathology, Vol. 6, No. 4, 2005, pp. 395-409. doi:10.1111/j.1364-3703.2005.00295.x [2] D. B. Redfern and J. Stenlid, “Spore Dispersal and Infec- tion,” In: S. Woodward, J. Stenlid, R. Karjalainen and A. HuØttermann, Eds., Heterobasidion annosum: Biology, Ecology, Impact and Control, CAB International, Wal- lingford, 1998, pp. 105-124. [3] T. Piri and K. Korhonen, “The Effect of Winter Thinning on the Spread of Heterobasidion parviporum in Norway Spruce Stands,” Canadian Journal of Forest Research, Vol. 38, No. 10, 2008, pp. 2589-2595. doi:10.1139/X08-103 [4] W. A. Sinclair and H. H. Lyon, “Diseases of Trees and Shrubs,” Cornell University Press, Ithaca, New York, 2005. [5] J. Taubert, “Temperature Requirements for Germination of Conidiospores and Growth of Mycelia of Hetero- basidion annosum s.s. and Heterobasidion parviporum,” Master Thesis No. 123. Swedish University of Agricul- tural Sciences, Alnarp, 2008. http://ex-epsilon.slu.se:8080/archive/00002863/01/Tauber t_J_081006.pdf [6] N. Samils, A. Olson and J. Stenlid, “The Capacity in Heterobasidion annosum s.l. to Resist Overgrowth by the Biocontrol Agent Phlebiopsis gigantea Is a Heritable Trait,” Biological Control, Vol. 45, No. 3, 2008, pp. 419- 426. doi:10.1016/j.biocontrol.2008.03.010 [7] O. Holdenrieder and B. J. W. Greig, “Biological Methods of Control,” In: S. Woodward, J. Stenlid, R. Karjalainen and A. HuØttermann, Eds., Heterobasidion annosum: Bi- ology, Ecology, Impact and Control, CAB International, Wallingford, 1998, pp. 235-258. [8] G. C. Papavizas, “Trichoderma and Gliocladium—Bio- logy, Ecology, and Potential for Biocontrol,” Annual Re- view of Phytopathology, Vol. 23, No. 1, 1985, pp. 23-54. doi:10.1146/annurev.py.23.090185.000323 [9] G. E. Harman, C. R. Howell, A. Viterbo, I. Chet and M. Lorito, “Trichoderma Species—Opportunistic, Avirulent Plant Symbionts,” Nature Reviews Microbiology, Vol. 2, No. 1, 2005, pp. 43-56. doi:10.1038/nrmicro797 [10] Z. Sierota, “Influence of Acidity on the Growth of Trichoderma viride Pers. ex Fr. and on the Inhibitory Ef- fect of its Filtrates Against Fomes annosus (Fr.) Cke in Artificial Cultures,” European Journal of Forest Pathol- ogy, Vol. 6, No. 5, 1976, pp. 302-311. doi:10.1111/j.1439-0329.1976.tb00540.x [11] H. Kwaśna, “Fungi on the Surface of Roots of Scots Pine and Its Stumps and their Effect on Heterobasidion anno- sum (Fr.) Bref. and Armillaria ostoyae (Romagn.) Herink Growth,” Roczniki Nauk Rolniczych Seria E, Ochrona Roslin, Vol. 26, No. 1, 1997, pp. 109-123. [12] O. Holdenrieder, “Investigations on Biological Control of Heterobasidion annosum in Norway Spruce (Picea abies) with Antagonistic Fungi. II. Interaction Experiments in Wood,” European Journal of Forest Pathology, Vol. 14, No. 3, 1984, pp. 137-153. doi:10.1111/j.1439-0329.1984.tb00938.x [13] M. Werner, A. Werner and R. Andrzejak, “Antagonistic Effects of Some Fungi on Fusarium oxysporum and Het- erobasidion annosum in Laboratory Experiment,” In: M. Manka, Ed., Environmental Biotic Factors in Integrated Plant Disease Control, The Polish Phytopathological So- ciety, Poznan, 1995, pp. 611-616. Copyright © 2012 SciRes. AiM  V. NIKOLAJEVA ET AL. Copyright © 2012 SciRes. AiM 302 [14] G. Nicolotti, P. Gonthier and G. C. Varese, “Effectiveness of Some Biocontrol and Chemical Treatments against Heterobasidion annosum on Norway Spruce Stumps,” European Journal of Forest Pathology, Vol. 29, No. 5, 1999, pp. 339-346. doi:10.1046/j.1439-0329.1999.00159.x [15] M. Berglund, J. Rönnberg, L. Holmer and J. Stenlid, “Comparison of Five Strains of Phlebiopsis gigantea and two Trichoderma Formulations for Treatment against Natural Heterobasidion Spore Infections on Norway Spruce Stumps,” Scandinavian Journal of Forest Re- search, Vol. 20, No. 1, 2005, pp. 12-17. doi:10.1080/02827580510008202 [16] A. Lehtijärvi, H. T. Doğmuş-Lehtijärvi, A. G. Aday and F. Oskay, “The Efficacy of Selected Biological and Chemi- cal Control Agents against Heterobasidion abietinum on Abies cilicica,” Forest Pathology, Vol. 41, No. 6, 2011. pp. 470-476. doi:10.1111/j.1439-0329.2010.00705.x [17] A. Tronsmo and C. Dennis, “Effect of Temperature on Antagonistic Properties of Trichoderma Species,” Trans- actions of the British Mycological Society, Vol. 71, No. 3, 1978, pp. 469-474. doi:10.1016/S0007-1536(78)80075-8 [18] B. Goldfarb, E. E. Nelson and E. M. Hansen, “Tricho- derma spp.: Growth Rates and Antagonism to Phellinus weirii in Vitro,” Mycologia, Vol. 81, No. 3, 1989, pp. 375-381. doi:10.2307/3760075 [19] Z. Antal, L. Manczinger, G. Szakacs, R. P. Tengerdy and L. Ferenczy, “Colony Growth, in Vitro Antagonism and Selection of Extracellular Enzymes in Cold-Tolerant Strains of Trichoderma Species,” Mycological Research, Vol. 104, No. 5, 2000, pp. 545-549. doi:10.1017/S0953756299001653 [20] M. Gardes and T. D. Bruns, “ITS Primers with Enhanced Specificity for Basidiomycetes—Application to the Iden- tification of Mycorrhizae and Rusts,” Molecular Ecology, Vol. 2, No. 2, 1993, pp. 113-118. doi:10.1111/j.1365-294X.1993.tb00005.x [21] T. J. White, T. Bruns, S. Lee and J. Taylor, “Amplifica- tion and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics,” In: M. A. Innis, D. H. Gelfand, J. J. Sninsky and T. J. White, Eds., PCR Protocols. A Guide to Methods and Applications, Academic Press, San Diego, 1990, pp. 315-322. [22] A. Asran-Amal, K. A. Abd-Elsalam, M. R. Omar and A. A. Aly, “Antagonistic Potential of Trichoderma spp. against Rhizoctonia solani and Use of M13 Microsatel- lite-Primed PCR to Evaluate the Antagonist Genetic Var- iation,” Journal of Plant Diseases and Protection, Vol. 112, No. 6, 2005, pp. 550-561. [23] R. M. Danielson and C. B. Davey, “Non Nutritional Fac- tors Affecting the Growth of Trichoderma in Culture,” Soil Biology and Biochemistry, Vol. 5, No. 5, 1973, pp. 495-504. doi:10.1016/0038-0717(73)90039-4 [24] Y. Yamazaki, M. Tojo, T. Hoshino, K. Kida, T. Saka- moto, et al., “Characterization of Trichoderma poly- sporum from Spitsbergen, Svalbard Archipelago, Norway, with Species Identity, Pathogenicity to Moss, and Poly- galacturonase Activity,” Fungal Ecology, Vol. 4, No. 1, 2011, pp. 15-21. doi:10.1016/j.funeco.2010.06.002 [25] W. M. Jaklitsch, G. J. Samuel, S. L. Dodd, B. S. Lu and I. S. Druzhinina, “Hypocrea rufa/Trichoderma viride: A Reassessment, and Description of Five Closely Related Species with and without Warted Conidia,” Studies in Mycology, Vol. 55, No. 1, 2006, pp. 135-177. doi:10.3114/sim.2006.56.04 [26] E. Lieckfeldt, G. J. Samuels, H. I. Nirenberg and O. Petrini, “A Morphological and Molecular Perspective of Trichoderma viride: Is It One or Two Species?” Applied and Environmental Microbiology, Vol. 65, No. 6, 1999, pp. 2418-2428. [27] P. K. Mukherjee and K. Raghu, “Effect of Temperature on Antagonistic and Biocontrol Potential of Trichoderma sp. on Sclerotium rolfsii,” Mycopathologia, Vol. 139, No. 3, 1997, pp. 151-155. doi:10.1023/A:1006868009184 [28] C. Guigon-Lopez, E. Carvajal-Millan, N. P. de Leon- Renova, F. Vargas-Albores, L. Bravo-Luna and V. M. Guerrero-Prieto, “Microcalorimetric Measurement of Tri- choderma spp. Growth at Different Temperatures,” Ther- mochimica Acta , Vol. 509, No. 1-2, 2010, pp. 40-45. doi:10.1016/j.tca.2010.05.018 [29] J. Köhl and E. Schlösser, “Decay of Sclerotia of Botrytis cinerea by Trichoderma spp. at Low Temperatures,” Journal of Phytopathology, Vol. 125, No. 4, 1989, pp. 320-326. doi:10.1111/j.1439-0434.1989.tb01076.x [30] D. K. Bell, H. D. Wells and C. R. Markham, “In Vitro Antagonism of Trichoderma Species against Six Fungal Plant Pathogens,” Phytopathology, Vol. 72, No. 4, 1982, pp. 379-382. doi:10.1094/Phyto-72-379 [31] A. Napierala-Filipiak and A. Werner, “Antagonism of Higher Fungi to Heterobasidion annosum (Fr.) Bref. in Laboratory Conditions,” Dendrobiology, Vol. 45, 2000, pp. 65-81. http://www.idpan.poznan.pl/images/stories/dendrobiology /vol45/45_65_81.pdf
|