S. SHUKLA, A. GOYAL 283
Characteristics of Mutants of Leuconostoc mesenteroides
B-742 Constitutive for Dextransucrases,” Enzyme and
Microbial Technology, Vol. 17, No. 8, 1995, pp. 689-695.
doi:10.1016/0141-0229(94)00021-I
[7] D. Kim and J. F. Robyt, “Dextransucrase Constitutive
Mutans of Leuconostoc mesenteroides B-1299,” Enzyme
and Microbial Technology, Vol. 17, No. 12, 1995, pp.
1050-1056. doi:10.1016/0141-0229(95)00039-9
[8] G. L. Cote, J. A. Ahlgren and M. R. Smith, “Some Struc-
tural Features of an Insoluble D-Glucan from a Mutant
Strain of Leuconostoc mesenteroides NRRL B-1355,”
Journal of Industrial Microbiology and Biotechnology,
Vol. 23, No. 1, 1999, pp. 656-660.
doi:10.1038/sj.jim.2900678
[9] H. Eifuku, A. Yoshimitsu-Narita, S. Sato, T. Yakushiji
and M. Inoue, “Production and Partial Characterization of
the Extracellular Polysaccharides from Oral Streptococ-
cus salivarius,” Carbohydrate Research, Vol. 194, 1989,
pp. 247-260. doi:10.1016/0008-6215(89)85023-2
[10] V. Monchois, R.-M. Willemot and P. Monsan, “Glucan-
sucrases: Mechanism of Action and Structure-Function
Relationships,” FEMS Microbiology Reviews, Vol. 23,
No. 2, 1999, pp. 131-151.
doi:10.1111/j.1574-6976.1999.tb00394.x
[11] V. Monchois, P. R.-S. Monsan and R. M. Willemot,
“Cloning and Sequencing of an Extracellular Dextransu-
crase (DSRB) from Leuconostoc mesenteroides NRRL
B-1299 Synthesizing Only α(1-6) Glucan,” FEMS Micro-
biology Letters, Vol. 159, No. 2, 1998, pp. 307-315.
doi:10.1111/j.1574-6968.1998.tb12876.x
[12] H. Abo, T. Matsumura, T. Kodama, H. Ohta, K. Fukui, K.
Kato and H. Kagawa, “Peptide Sequences for Sucrose
Splitting and Glucan Binding within Streptococcus so-
brinus Glucosyltransferase (Water-Insoluble Glucan Syn-
thetase),” Journal of Bacteriolology, Vol. 173, 1991, pp.
989-996.
[13] J. J. Ferretti, M. L. Gilpin and R. R. B. Russell, “Nucleo-
tide Sequence of a Glucosyltransferase Gene from Strep-
tococcus sobrinus Mfe28,” Journal of Bacteriology, Vol.
169, 1987, pp. 4271-4278.
[14] S. Patel, A. Majumder and A. Goyal, “Potentials of
exopolysaccharides from Lactic Acid Bacteria,” Indian
Journal of Microbiology, Vol. 52, No. 1, 2012, pp. 3-12.
doi:10.1007/s12088-011-0148-8
[15] R. K. Purama, A. Goyal, “Dextransucrase Production by
Leuconostoc mesenteroides,” Indian Journal of Microbi-
ology, Vol. 2, 2005, pp. 89-101.
[16] A. Singh, A. Majumder and A. Goyal, “Artificial Intelli-
gence Based Optimization of Exocellular Glucansucrase
Production from Leuconostoc dextranicum NRRL B-
1146,” Bioresource Technology, Vol. 99, No. 17, 2008,
pp. 8201-8206. doi:10.1016/j.biortech.2008.03.038
[17] R. K. Purama and A. Goyal, “Screening and Optimization
of Nutritional Factors for Higher Dextransucrase Produc-
tion by Leuconostoc mesenteroides NRRL B-640 Using
Statistical Approach,” Bioresource Technology, Vol. 99,
No. 15, 2008, pp. 7108-7114.
doi:10.1016/j.biortech.2008.01.032
[18] S. Patel, D. Kothari and A. Goyal, “Enhancement of Dex-
transucrase Activity of Pediococcus pentosaceus Mutant
SPAm1 by Response Surface Methodology,” Indian Jour-
nal of Microbiology, Vol. 10, 2011, pp. 346-351.
[19] S. Shukla and A. Goyal, “16S rRNA Based Identification
of a Glucan Hyper-Producing Weissella confuse,” Enzyme
Research, 2011, Article ID: 250842.
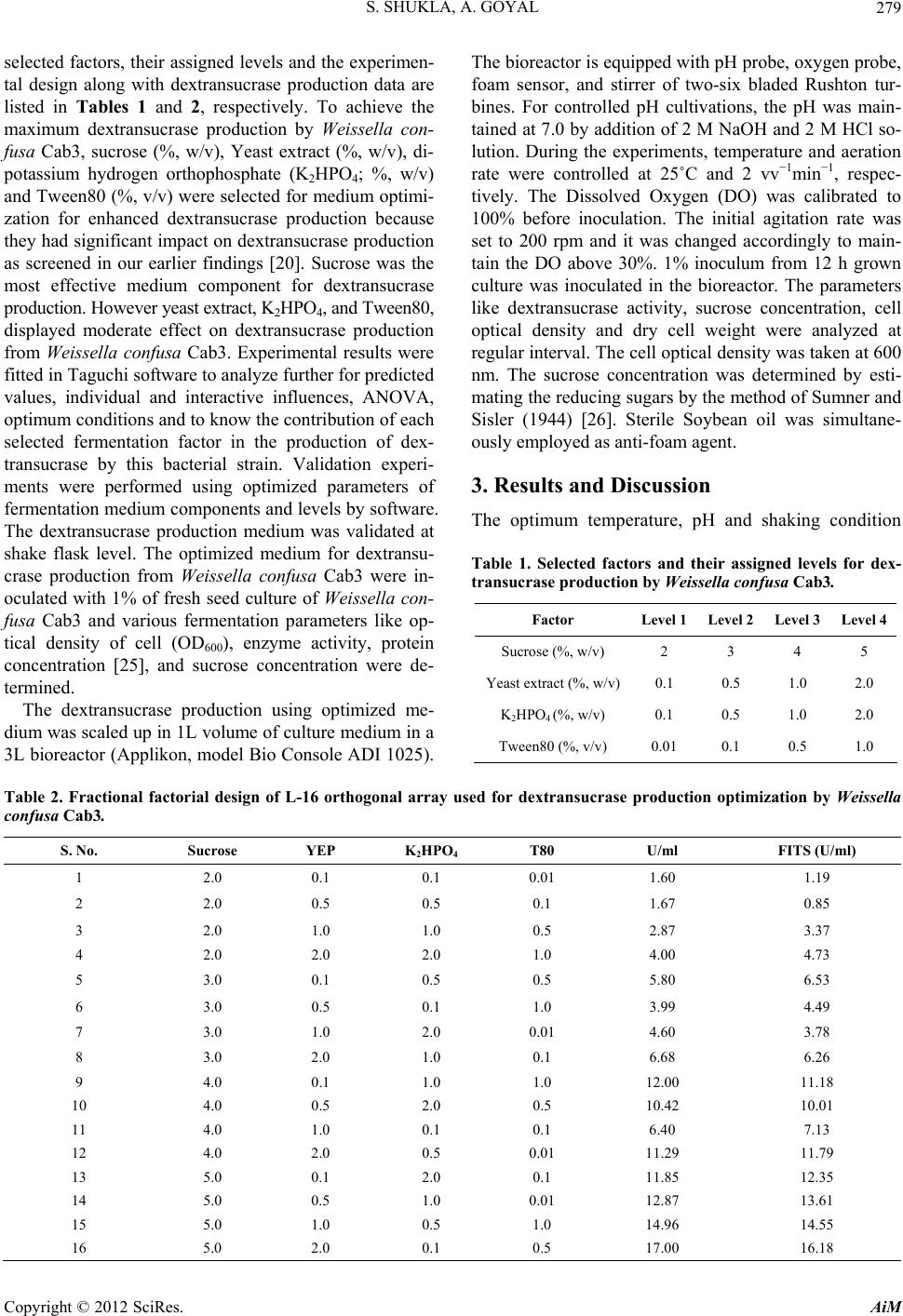
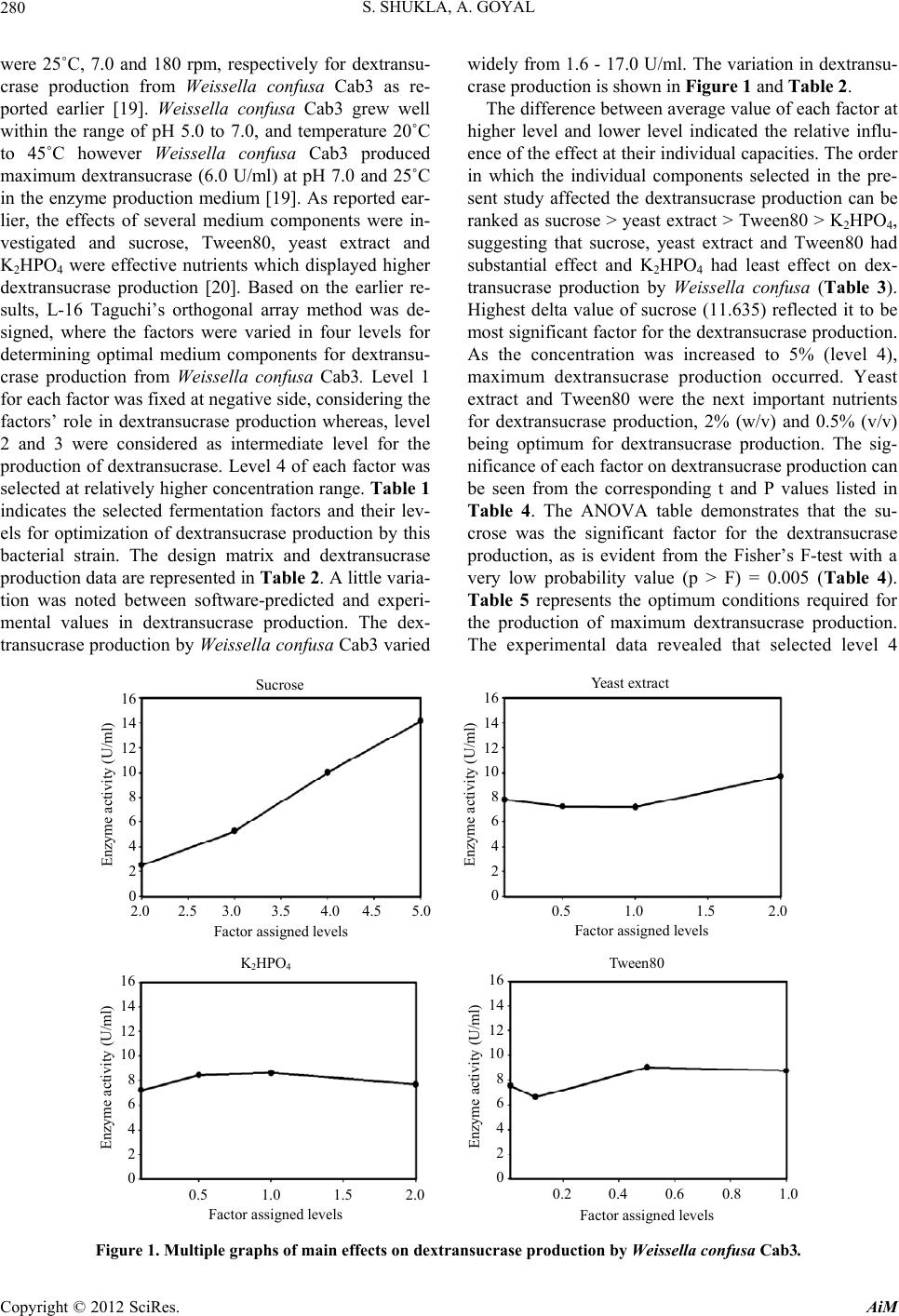
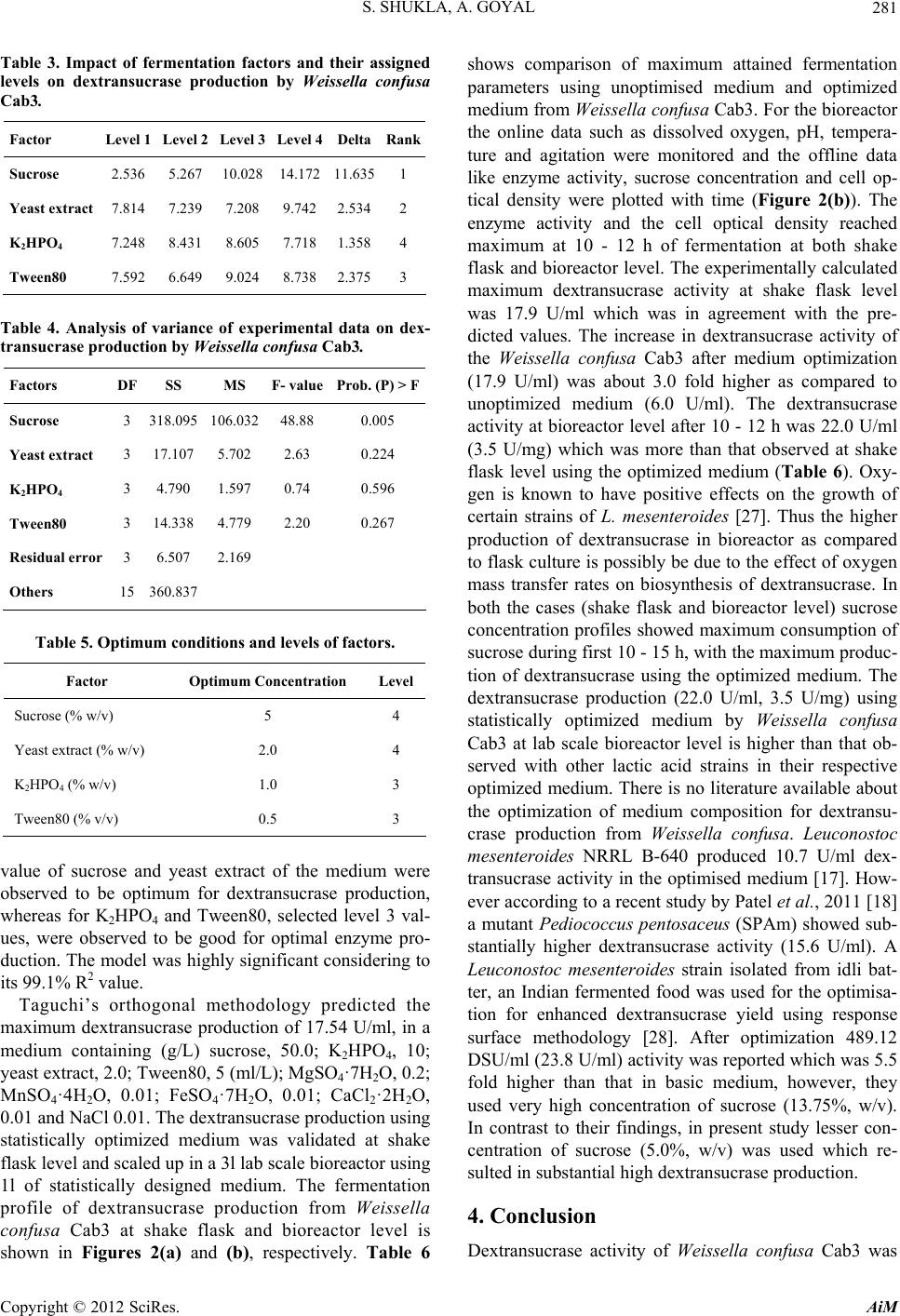
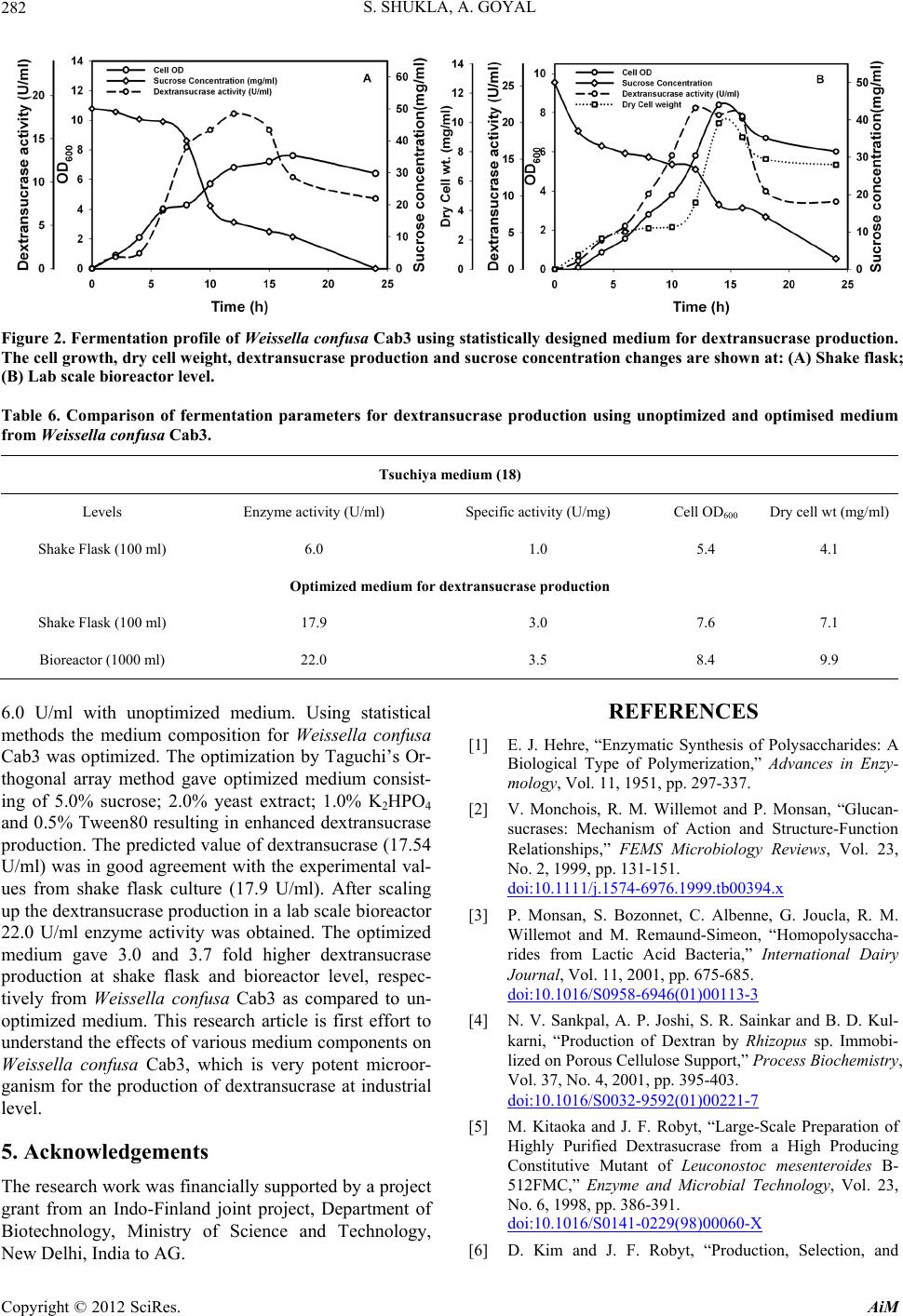
[20] S. Shukla and A. Goyal, “Optimization of Fermentation
Medium for Enhanced Glucansucrase and Glucan Pro-
duction from Weissella confusa,” Brazilian Archives of
Biology and Technology, Vol. 54, No. 6, 2011, pp. 1117-
1124.
[21] J. C. De Man, M. Rogosa and M. E. Sharpe, “A Medium
for the Cultivation of lactobacilli,” Journal of Applied
Bacteriology, Vol. 23, No. 1, 1960, pp. 130-135.
doi:10.1111/j.1365-2672.1960.tb00188.x
[22] H. M. Tsuchiya, H. J. Koepsell, J. Corman, G. Bryant, M.
O. Bogard, V. H. Feger and R. W. Jackson, “The Effect
of Certain Culture Factors on Production on Dextransu-
crase by Leuconostoc mesenteroides,” Journal of Bacte-
riology, Vol. 64, 1952, pp. 521-526.
[23] N. Nelson, “A Photometric Adaptation of the Somoyogi
Method for the Determination of Glucose,” Journal of
Biological Chemistry, Vol. 153, 1944, pp. 375-380.
[24] M. Somogyi, “A New Reagent for the Determination of
Sugars,” Journal of Biological Chemistry, Vol. 160, 1945,
pp. 61-68.
[25] O. H. Lowry, N. J. Rosebrough, A. L. Farr and R. J.
Randall, “Protein Measurement with the Folin Phenol
Reagent,” Journal of Biological Chemistry, Vol. 193, 1951,
pp. 265-275.
[26] J. B. Sumner and E. B. Sisler, “A Simple Method for
Blood Sugar,” Archives of Biochemistry, Vol. 4, 1944, pp.
333-336.
[27] V. B. Veljkovic, M. L. Lazic, D. J. Rutic, S. M. Jovano-
vic and D. U. Skala, “Effect of Aeration on Extracellular
Dextran Production by Leuconostoc mesenteroides,” En-
zyme and Microbial Technology, Vol. 14, No. 8, 1992, pp.
665-668. doi:10.1016/0141-0229(92)90044-O
[28] S. D. Sawale and S. S. Lele, “Increased Dextransucrase
Production by Response Surface Methodology from Leu-
conostoc Species; Isolated from Fermented Idli Batter,”
Global Journal of Biotechnology and Biochemistry, Vol.
4, No. 2, 2009, pp. 160-167.
Copyright © 2012 SciRes. AiM