Paper Menu >>
Journal Menu >>
 Advances in Microbiology, 2012, 2, 201-207 http://dx.doi.org/10.4236/aim.2012.23024 Published Online September 2012 (http://www.SciRP.org/journal/aim) Specific Antigens to Distinguish M. tuberculosis from M. avium Qun Liang1,2*, Lingxia Zhang3*, Zeng Tu1,4*, Jingyu Wang1,2, Tao Hu1, Pengzhi Wang1, Weili Wu1, Qi Liu5, Yanlin Zhao6, Yan Li7#, Weijun Chen1,5# 1Key Laboratory of Genome Sciences and Information, Beijing Institute of Genomics, Chinese Academy of Sciences, Beijing, China 2Graduate University of the Chinese Academy of Sciences, Beijing, China 3Beijing 309th Hospital of PLA, Beijing, China 4Department of Pathogenic Biology, Chongqing Medical University, Chongqing, China 5Beijing Genomics Institute in Shenzhen, Shenzhen, China 6Beijing Tuberculosis and Thoracic Tumor research Institute, Beijing, China 7The Affiliated Hospital of the Academy of Millitary Medical Sciences, Beijing, China Email: liangqun82623@gmail.com, zhanglingxia7@yahoo.com.cn, Tuz1980@126.com, icandowjy@yahoo.com.cn, hutao8426@126.com, wangpzh@genomics.org.cn, wuwl@big.ac.cn, hellofqaa@126.com, zhaoyanlin@tb123.org, #liyanmd@yahoo.com.cn, #chenwj@genomics.org.cn Received April 15, 2012; revised May 23, 2012; accepted June 4, 2012 ABSTRACT To distinguish Mycobacterium tuberculosis from Mycobacterium avium, specific M. tuberculosis antigens had been studied for improving the early differential diagnosis effect of tuberculosis caused by different Mycobacterium. The rabbit anti-M. avium sera and anti-M. tuberculosis sera were analyzed for antibody-based reactivity by matrix-assisted laser desorption-ionization mass spectrometry (MALDI-TOF Mass) against M. tuberculosis proteins. The immunoreac- tive spots, which were attributed to the proteins HspX, GroES and CFP-10, were mostly located at 10 - 60 kDa and PI 4 - 6, subsequently Western blotting result proved that HspX and CFP-10 were specific to M. tuberculosis and ELISA test- ing result of 30 M. a vium positive sera showed that GroES were cross-reactive to M. avium. Lastly, positive and nega- tive tuberculosis reference sera and based on the mechanism of indirect ELISA, the specificity and the sensitivity of the methods targeting the antibodies HspX, GroES or CFP-10 were evaluated at 37% and 26%, 12% and 97%, 81% and 98%, respectively. The combination of these three antibody detection methods allowed to reached a specificity of 42%, and of 39% without taken into account of the method targeting the GroES antibody. Using proteomics approach, we found three M. tuberculosis specific antigens showed good potential in tuberculosis diagnosis, providing basic study for serodiagnosis of tuberculosis. Keywords: Mycobacterium tuberculosis; Mycobacterium avium; Mass Spectrometry; Immunodetection 1. Introduction Tuberculosis (TB) remains a major cause of death in de- veloping countries. It is also rising throughout the indus- trialized countries, partly as the cause of human immu- nodeficiency virus (HIV) infection. Mycobacterium tu- berculosis complex (MTC) members such as M. tuber- culosis are the pathogens of TB infection and Nontuber- culous mycobacteria (NTM) such as Mycobacterium avium are responsible of mycobacteria [1,2]. Unfortu- nately, NTM is resistant to many general anti-TB drugs, leading the patients to suffer a years prolonged course and finally become chronic or refractory cases of myco- bacteria [3,4]. However, clinically it is difficult to dis- tinguish between them, because they are similar in clini- cal manifestations, imaging study, smear and culture, tuberculin tests and pathological examination [5-7]. There- fore, the rapid identification of M. tuberculosis and M. avium is of extreme importance to diagnosis, effective chemotherapy and control transmission of TB. Represented by two-dimensional gel electrophoresis (2-D) and mass spectrometry (MS) technology, pro- teomics has provided some encouraging results in TB research [8-10]. So far, important antigens of M. tuber- culosis have been identified such as the 38-kDa antigen, early secreted antigenic target (ESAT-6), antigen 85B, the proteins encoded by Rv3872 [11,12]. However, the cross-reaction between M. tuberculosis and M. avium makes these antigens unable to distinguish these myco- *These authors contributed equally to this article. #Corresponding authors. C opyright © 2012 SciRes. AiM  Q. LIANG ET AL. 202 bacterial species and consequently differentiate those respective infections [13]. Only a few secreted proteins such as 14-kDa protein and CFP-10 have been further characterized which was demonstrated by Western blot- ting in culture filtrates of M. avium but was not detected in M. tuberculosis [14]. CFP-10, molecular weight of 10 kDa, mainly existed in M. tuberculosis culture filtrate. Previous data have shown that it is a specific potential antigen to M. tuberculosis [15,16]. Renshaw et al. [17] reported that BCG and NTM were lack of CFP-10 coding. HspX-14 was predicted to be an important membrane antigen, which increased expression along with M. tu- berculosis growing into stable phase from logarithmic phase [18] and in anaerobic condition [19]. Some data also showed HspX-14 could induce humoral immune response [20,21]. GroES, a molecular chaperone protein, joints participation in protein folding, assembly, transport and degradation [22]. The high abundance of GroES possibly led antigen presenting cells to secrete many small peptides which could induce the host produced a strong immunological response [23]. These reports sug- gested that these proteins may be the M. tuberculosis- specific antigenicity. Comparison of immunoblot profiles of rabbit anti-M. tuberculosis sera and rabbit anti-M. avium sera reacting with extracts of M. tuberculosis could detect specific antigens allowing to distinguish easily these two myco- bacteria species and helped the serodiagnostic test for tuberculosis. 2. Materials Mycobacterium tuberculosis H37Rv, rabbit anti-M. tu- berculosis sera and rabbit anti-M. avium sera were ob- tained from Tuberculosis Research Institute of the Peo- ple’s Liberation Army Hospital 309 (Beijing, China). 300 sera from healthy blood donators were collected from Beijing Red-cross blood Central which were tested negatively to TB by Elisa kits (Chengdu Yongan Phar- maceutical Co. Ltd). 100 sera from M. tuberculosis patients were collected in 309 Hospital (Beijing, China). Basis of diagnosis: Two sputum samples examined by smear microscopy-stained for acid-fast bacilli or AFB are positive and chest X-ray visualizes the chest shadow caused by tubercular lesion. Then used COBAS AMPLICOR™ M. tuberculosis Kit (Roche) to confirm. 30 sera from M. avium patients were collected in 309 Hospital (Beijing, China). Basis of diagnosis: Used COBAS AMPLICOR™ M. avium Kit (Roche) to detect M. avium and used COBAS AMPLICOR™ M. tubercu- losis Kit (Roche) to confirm they were not infected by M. tuberculosis. 200 sera from purified protein derivative (PPD)-posi- tive healthy body-examining people were collected in Affiliated Hospital of the Academy of Millitary Medical Sciences. 3. Methods 3.1. Protein Preparation M. tuberculosis H37Rv was cultured in modified Sauton medium for 4 weeks and heat-inactivated at 80˚C for 1 hr. The bacteria were harvested by centrifugation and were washed three times with 10 mM Tris buffer (pH 7.4), then suspended in 500 µl lysate (0.3% SDS, 200 mM DTT, 50 mM Tris, pH 7) and sonicated for 30 min (250 W, 2 sec pulse-on, 4 sec pulse-off intervals), subse- quently added the Benzonase nuclease (1:20; Novagen, Merck KgaA, Darmstadt, Germany) and kept at 4˚C for 1 hr, and then centrifuged at 22,300 g for 30 min. Used 2-D Clean-up Kit (GE Healthcare, Fairfield, USA) to purified protein. 3.2. 2-D PAGE Samples of 200 μg protein suspended in rehydration buffer (6 M Carbamide, 0.71 M SDS, 0.375 M Tris pH 8.8, 20% glycerol) were applied on immobilized pH 4 - 7 linear gradient strips (13 cm; GE Healthcare). Focusing at 20˚C using the following four-step program: a) 50 V, 6 hr; b) 500 V, 1 hr; c) 1000 V, 1 hr; d) 8000 V constant until 66,000 Vh. The current limit was set at 50 μA/strip. After isoelectric focusing (IEF), each strip was equili- brated for 15 min in equilibration buffer I (rehydration buffer, 0.13 M DTT) followed by equilibration buffer II (rehydration buffer, 0.14 M iodoacetamide) for 15 min. The second dimension separation was performed in uni- form 12.5% SDS-PAGE gels. Then silver stained the gels. 3.3. Western Blotting Preparing for Western blotting, proteins was electropho- retically transferred by using TE70 Semi-Dry Transfer (Amersham Pharmacia Biotech, Uppsala, Sweden) to 0.2-µm-pore-size polyvinylidene difluoride membrane (Immun-Blot PDVF membrane; Bio-Rad, Hercules, CA), which was then washed and blocked in Tris-buffered saline-Tween 20 (TBS-T) containing 1% Tween 20 and 5% non-fat dry milk for 120 min, next incubated in the rabbit anti-M. tuberculosis and anti-M. avium sera (both 1:250 TBS-T diluted) for 2 hr, then reacted with horse- radish peroxidase labeled goat anti-rabbit IgG antibody (Santa Cruz Biotechnology, Santa Cruz, USA) (1:2500 TBS-T diluted) for 1 hr, subsequently washed exten- sively in TBS-T and then add freshly prepared DAB so- lution (Beijing Saichi Shengwu Keji, Beijing, China ) for color. Signal was detected by using UMAX scanner and Copyright © 2012 SciRes. AiM 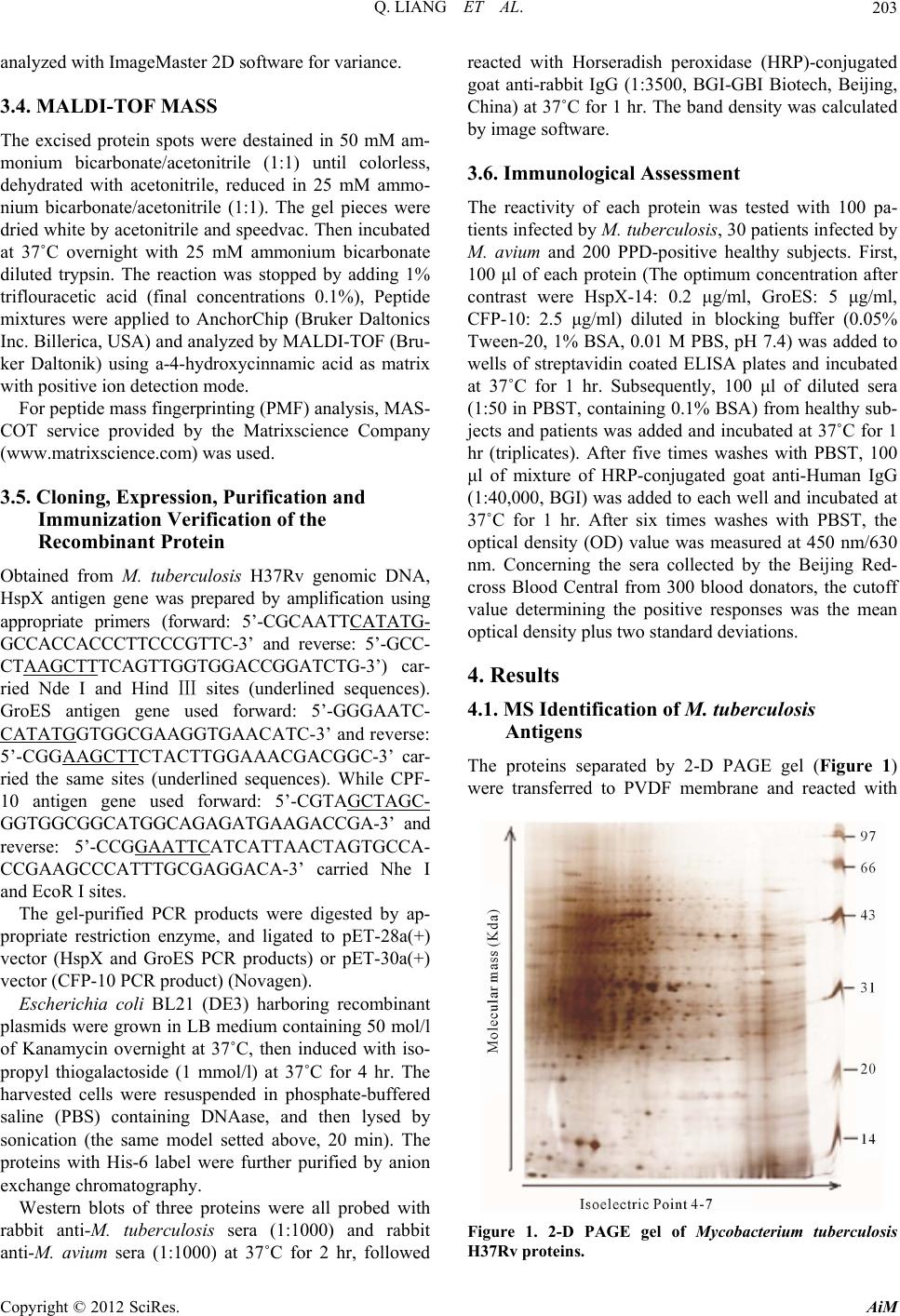 Q. LIANG ET AL. 203 analyzed with ImageMaster 2D software for variance. 3.4. MALDI-TOF MASS The excised protein spots were destained in 50 mM am- monium bicarbonate/acetonitrile (1:1) until colorless, dehydrated with acetonitrile, reduced in 25 mM ammo- nium bicarbonate/acetonitrile (1:1). The gel pieces were dried white by acetonitrile and speedvac. Then incubated at 37˚C overnight with 25 mM ammonium bicarbonate diluted trypsin. The reaction was stopped by adding 1% triflouracetic acid (final concentrations 0.1%), Peptide mixtures were applied to AnchorChip (Bruker Daltonics Inc. Billerica, USA) and analyzed by MALDI-TOF (Bru- ker Daltonik) using a-4-hydroxycinnamic acid as matrix with positive ion detection mode. For peptide mass fingerprinting (PMF) analysis, MAS- COT service provided by the Matrixscience Company (www.matrixscience.com) was used. 3.5. Cloning, Expression, Purification and Immunization Verification of the Recombinant Protein Obtained from M. tuberculosis H37Rv genomic DNA, HspX antigen gene was prepared by amplification using appropriate primers (forward: 5’-CGCAATTCATATG- GCCACCACCCTTCCCGTTC-3’ and reverse: 5’-GCC- CTAAGCTTTCAGTTGGTGGACCGGATCTG-3’) car- ried Nde I and Hind Ⅲ sites (underlined sequences). GroES antigen gene used forward: 5’-GGGAATC- CATATGGTGGCGAAGGTGAACATC-3’ and reverse: 5’-CGGAAGCTTCTACTTGGAAACGACGGC-3’ car- ried the same sites (underlined sequences). While CPF- 10 antigen gene used forward: 5’-CGTAGCTAGC- GGTGGCGGCATGGCAGAGATGAAGACCGA-3’ and reverse: 5’-CCGGAATTCATCATTAACTAGTGCCA- CCGAAGCCCATTTGCGAGGACA-3’ carried Nhe I and EcoR I sites. The gel-purified PCR products were digested by ap- propriate restriction enzyme, and ligated to pET-28a(+) vector (HspX and GroES PCR products) or pET-30a(+) vector (CFP-10 PCR product) (Novagen). Escherichia coli BL21 (DE3) harboring recombinant plasmids were grown in LB medium containing 50 mol/l of Kanamycin overnight at 37˚C, then induced with iso- propyl thiogalactoside (1 mmol/l) at 37˚C for 4 hr. The harvested cells were resuspended in phosphate-buffered saline (PBS) containing DNAase, and then lysed by sonication (the same model setted above, 20 min). The proteins with His-6 label were further purified by anion exchange chromatography. Western blots of three proteins were all probed with rabbit anti-M. tuberculosis sera (1:1000) and rabbit anti-M. avium sera (1:1000) at 37˚C for 2 hr, followed reacted with Horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG (1:3500, BGI-GBI Biotech, Beijing, China) at 37˚C for 1 hr. The band density was calculated by image software. 3.6. Immunological Assessment The reactivity of each protein was tested with 100 pa- tients infected by M. tuberculosis, 30 patients infected by M. avium and 200 PPD-positive healthy subjects. First, 100 μl of each protein (The optimum concentration after contrast were HspX-14: 0.2 μg/ml, GroES: 5 μg/ml, CFP-10: 2.5 μg/ml) diluted in blocking buffer (0.05% Tween-20, 1% BSA, 0.01 M PBS, pH 7.4) was added to wells of streptavidin coated ELISA plates and incubated at 37˚C for 1 hr. Subsequently, 100 μl of diluted sera (1:50 in PBST, containing 0.1% BSA) from healthy sub- jects and patients was added and incubated at 37˚C for 1 hr (triplicates). After five times washes with PBST, 100 μl of mixture of HRP-conjugated goat anti-Human IgG (1:40,000, BGI) was added to each well and incubated at 37˚C for 1 hr. After six times washes with PBST, the optical density (OD) value was measured at 450 nm/630 nm. Concerning the sera collected by the Beijing Red- cross Blood Central from 300 blood donators, the cutoff value determining the positive responses was the mean optical density plus two standard deviations. 4. Results 4.1. MS Identification of M. tuberculosis Antigens The proteins separated by 2-D PAGE gel (Figure 1) were transferred to PVDF membrane and reacted with Figure 1. 2-D PAGE gel of Mycobacterium tuberculosis H37Rv proteins. Copyright © 2012 SciRes. AiM 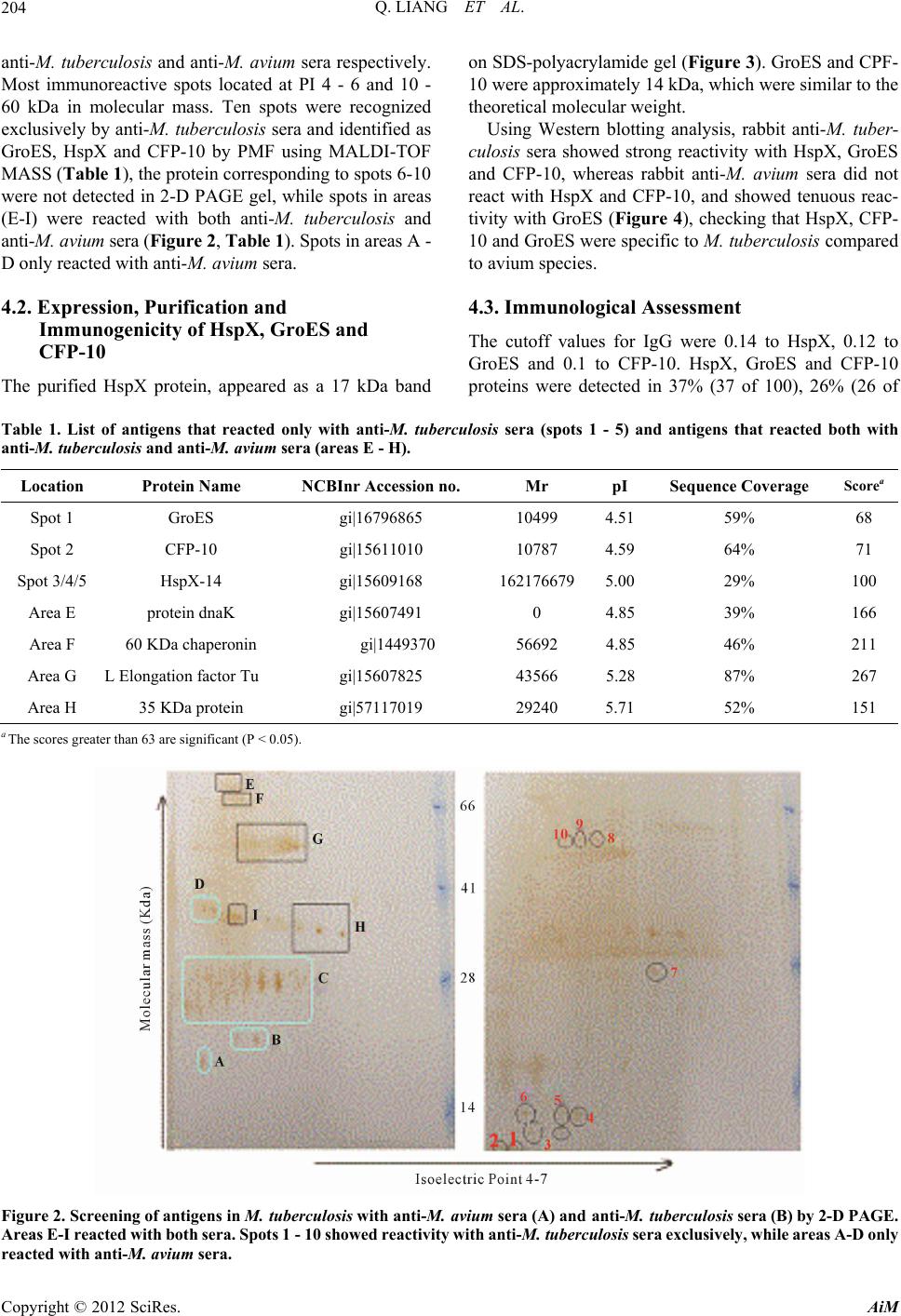 Q. LIANG ET AL. AiM 204 anti-M. tuberculosis and anti-M. avium sera respectively. Most immunoreactive spots located at PI 4 - 6 and 10 - 60 kDa in molecular mass. Ten spots were recognized exclusively by anti-M. tuberculosis sera and identified as GroES, HspX and CFP-10 by PMF using MALDI-TOF MASS (Table 1), the protein corresponding to spots 6-10 were not detected in 2-D PAGE gel, while spots in areas (E-I) were reacted with both anti-M. tuberculosis and anti-M. avium sera (Figure 2, Table 1). Spots in areas A - D only reacted with anti-M. avium sera. Copyright © 2012 SciRes. 4.2. Expression, Purification and Immunogenicity of HspX, GroES and CFP-10 The purified HspX protein, appeared as a 17 kDa band on SDS-polyacrylamide gel (Figure 3). GroES and CPF- 10 were approximately 14 kDa, which were similar to the theoretical molecular weight. Using Western blotting analysis, rabbit anti-M. tuber- culosis sera showed strong reactivity with HspX, GroES and CFP-10, whereas rabbit anti-M. avium sera did not react with HspX and CFP-10, and showed tenuous reac- tivity with GroES (Figure 4), checking that HspX, CFP- 10 and GroES were specific to M. tuberculosis compared to avium species. 4.3. Immunological Assessment The cutoff values for IgG were 0.14 to HspX, 0.12 to GroES and 0.1 to CFP-10. HspX, GroES and CFP-10 proteins were detected in 37% (37 of 100), 26% (26 of Table 1. List of antigens that reacted only with anti-M. tuberculosis sera (spots 1 - 5) and antigens that reacted both with anti-M. tuberculosis and anti-M. avium sera (areas E - H). Location Protein Name NCBInr Accession no. Mr pI Sequence Coverage Scorea Spot 1 GroES gi|16796865 10499 4.5159% 68 Spot 2 CFP-10 gi|15611010 10787 4.5964% 71 Spot 3/4/5 HspX-14 gi|15609168 1621766795.0029% 100 Area E protein dnaK gi|15607491 0 4.8539% 166 Area F 60 KDa chaperonin gi|1449370 56692 4.8546% 211 Area G L Elongation factor Tu gi|15607825 43566 5.2887% 267 Area H 35 KDa protein gi|57117019 29240 5.7152% 151 a The scores greater than 63 are significant (P < 0.05). Figure 2. Screening of antigens in M. tuberculosis with anti-M. avium sera (A) and anti-M. tuberculosis sera (B) by 2-D PAG E. Areas E-I react ed with both sera. Spots 1 - 10 showed reactivit y with anti-M. tuberculosis sera ex clusively, while areas A-D only reacted with anti-M. avium sera.  Q. LIANG ET AL. 205 Figure 3. Expression of HspX, GroES and CFP-10 antigens (12.5% SDS-PAGE). (a) (b) Figure 4. Validation of the antigens specific to M. tuberculo- sis. Western blotting analysis of the equal amount expressed HspX, GroES and CFP-10 proteins separated by SDS- PAGE, were performed with either anti-M. tuberculosis or anti-M. avium sera for 3 times. 100) and 12% (12 of 100) of M. tuberculosis patients sera respectively (Table 2). Moreover, two sera of the samples were tested positive with CFP-10 but negative with the other two proteins and another three sera were positive with GroES protein solely. While only GroES Table 2. Serological reactivity of CFP-10, GroES and HspX-14 in M. tuberculosis patients, M. avium patients and healthy PPD+ individuals. No. of samples positive by ELISA for: Status No. of persons CaGbHc C+G C+H G+HC+G+H M. tuberculosis+100 122637 28 39 4042 M. avium+30 070 7 0 7 7 Healthy, PPD+ 200 3355 36 6 3738 aC: CFP-10; bG: GroES; cH: HspX-14. protein (7 of 30) was detected in sera of M. avium pa- tients. To evaluate the specificity, two hundred of sera from PPD-positive health body-examining people were tested. The specificity of HspX, GroES and CFP-10 were 97.5% (195 of 200), 82.5% (165 of 200) and 98.5% (197 of 200), respectively (Table 2). Similarly, 1 sera was tested positive only with CFP-10, while 2 sera only with HspX- 14. 5. Discussion At present, TB remains a serious threat to human health. A rapid diagnostic method of early stage of TB infection will undoubtedly play a decisive role in TB control [24], yet various similar pathological manifestations make them difficult to distinguish [5-7]. Recently, more and more M. avium infection has been reported [25-28]. However, the research about the distinction between M. tuberculosis and M. avium is rare [14]. Using a serological proteomic approach to detect spe- cifically by identifying candidate antigen has been re- ported by previous research [8-10]. Due to high lipid content of M. tuberculosis, we chose 2-D Clean-up kit to purify the crude protein and optimized some other pro- teomics experimental condition of M. tuberculosis, in- cluding the IEF voltage settings, staining method, trans- fer membrane current and time settings. Three proteins (CFP-10, GroES, HspX-14) were identified as potentially specific antigen of anti-M. tuberculosis by 2-D PAGE, Western blotting and MALDI-TOF methods in our study. CFP10, one of the antigens selected by our method was consistent with what was reported [14], which suggested the approach could screen out specific antigens. The reuslt of Immunoblotting assays showed that CFP-10 and HspX-14 only reacted with the anti-M. tu- berculosis sera while the GroES protein cross-reacted with the anti-M. avium sera slightly (Figure 4). The result of testing for specificity of these recombi- nant proteins showed that the specific response to protein GroES was relatively lower. M. tuberculosis GroES, Copyright © 2012 SciRes. AiM  Q. LIANG ET AL. 206 having a part homology sequence with human homolo- gous protein, often causes the emergence of spontaneous autoimmune diseases such as rheumatoid arthritis, sys- temic sclerosis, psoriasis, and ankylosing spondylitis [28, 29]. It may be the causes of lower specificity. The result of evaluating the sensitivity of theses re- combinant proteins showed a low sensitivity (37%, 26%, 12%). This was consitent with the previous M. tubercu- losis antibody detection studies [16,30,31]. Nonetheless, CFP-10, GroES and HspX-14 proteins as test antigens were complementary in detecting M. tuberculosis spe- cific antibodies and could effectively improve the detec- tion of M. tuberculosis antibodies (42%). Ignoring the contribution of GroES, the detection ratio of CFP-10 and HspX-14 proteins as a test antigen could reach 39%. In summary, we detected three M. tuberculosis spe- cific antigens, which could be used in order to distin- guish M. tuberculosis from M. avium. By system re- search of HspX-14, GroES and CFP-10, we found that HspX-14 and CFP-10 showed better potentiality in TB diagnosis. 6. Acknowledgements The work was supported by the research funding from Infectious Diseases Special Project, Minister of Health of China (2008ZX10003—009) and Beijing Science and Technology Program (Z08050703080801). 7. Disclosure I certify that all my affiliations with or financial in- volvement in, within the past 5 years and foreseeable future, any organization or entity with a financial interest in or financial conflict with the subject matter or materi- als discussed in the manuscript are completely disclosed. REFERENCES [1] D. S. Prince, D. D. Peterson and R. M. Steiner, “Infection with Mycobacterium avium Complex in Patients without Predisposing Conditions,” New England Journal of Medi- cine, Vol. 321, No. 13, 1989, pp. 863-868. doi:10.1056/NEJM198909283211304 [2] A. I. Zumla and J. Grange, “Nontuberculous mycobacte- rial Pulmonary Infections,” Clinics in Chest Medicine, Vol. 23, No. 2, 2002, pp. 369-376. doi:10.1016/S0272-5231(01)00011-9 [3] M. Payton, R. Auty, R. Delgoda, M. Everett and E. Sim, “Cloning and Characterization of Arylamine n-Acetyl- transferase Genes from Mycobacterium smegmatis and Mycobacterium tuberculosis: Increased Expression Re- sults in Isoniazid Resistance,” Journal of Bacteriology, Vol. 184, No. 4, 1999, pp. 1343-1347. [4] D. E. Griffith, B. A. Brown, P. Cegielski, D. T. Murphy and R. J. Wallace Jr., “Early Results (at 6 Months) with Intermittent Clarithromycin Including Regimens for Lung Disease Due to Mycobacterium avium Complex,” Cli- nical Infectious Diseases, Vol. 30, No. 2, 2000, pp. 288- 292. doi:10.1086/313644 [5] R. J. Seballos, A. L. Walsh and A. C. Mehta, “Clinical Evaluation of a Liquid Chemical Sterilization System for the Flexible Bronchoscope,” Journal of Bronchology, Vol. 2, No. 3, 1995, pp. 192-199. doi:10.1097/00128594-199507000-00005 [6] C. F. Von Reyn, D. E. Williams, C. R. Horsburgh, A. S. Jaeger, B. J. Marsh, K. Haslovk and M. Magnusson, “Dual Skin Testing with Mycobacterium avium Sensitin and Purified Protein Derivative to Discriminate Pulmo- nary Disease Due to M. avium Complex from Pulmonary Disease Due to Mycobacterium tuberculosis,” Journal of Infection Diseases, Vol. 177, No. 3, 1998, pp. 730-736. doi:10.1086/514225 [7] A. M. Middleton, M. V. Chadwick, A. G. Nicholson, R. Wilson, D. J. Thomton, S. Kirkham and J. K. Sheehan, “Interaction between Mycobacteria and Mucus on a Hu- man Respiratory Tissue Organ Culture Model with an Air Interface,” Experimental Lung Research, Vol. 30, No. 1, 2004, pp. 17-29. doi:10.1080/01902140490252876 [8] P. R. Jungblut, E. C. Muller, J. Mattow and S. H. E. Kaufmann, “Proteomics Reveals Open Reading Frames in Mycobacterium tuberculosis H37Rv Not Predicted by Genomics,” Infection and Immunity, Vol. 69, No. 9, 2001, pp. 5905-5907. doi:10.1128/IAI.69.9.5905-5907.2001 [9] J. Starck, G. Källenius, B. I. Marklund, D. I. Andersson and T. Åkerlund, “Comparative Proteome Analysis of Mycobacterium tuberculosis Grown under Aerobic and Anaerobic Conditions,” Microbiology, Vol. 150, No. 11, 2004, pp. 3821-3829. doi:10.1099/mic.0.27284-0 [10] S. W. Ryoo, Y. K. Park, S. N. Park, Y. S. Shim, H. Liew, S. Kang and G. H. Bai, “Comparative Proteomic Analysis of Virulent Korean Mycobacterium tuberculosis K-Strain with Other Mycobacteria Strain Following Infection of U-937 Macrophage,” Journal of Microbiology, Vol. 45, No. 3, 2007, pp. 268-271. [11] P. Andersen, A. B. Andersen, A. L. Sorensen and S. Na- gai, “Recall of Longlived Immunity to Mycobacterium tuberculosis Infection in Mice,” Journal of Immunology, Vol. 154, No. 7, 1995, pp. 3359-3372. [12] E. D. Chan, R. Reves, J. T. Belisle, P. J. Brennan and W. E. Hahn, “Diagnosis of Tuberculosis by a Visually De- tectable Immunoassay for Lipoarabinomannan,” Ameri- can Journal of Respiratory and Critical Care Medicine, Vol. 161, No. 5, 2000, pp. 1713-1719. [13] U. Demkow, M. Filewska, B. Bialas, M. Szturmowicz, T. Zielonka, S. Wesołowski, J. Kuś, J. Ziołkowski, E. Aug- ustynowicz-Kopeć, Z. Zwolska, E. Skopińiska-Rábewska and E. Rowińska-Zakrzewska, “Antimycobacterial Anti- body Level in Pleural, Pericardial and Cerebrospinal Fluid of Patients with Tuberculosis,” Pneumonologia I Alergologia Polska, Vol. 72, No. 3, 2004, pp. 105-110. [14] I. Olsen, L. J. Reitan and H. G. Wiker, “Distinct Differ- ences in Repertoires of Low-Molecular-Mass Secreted Antigens of Mycobacterium avium Complex and Myco- bacterium tuberculosis,” Journal of Clinical Microbiol- Copyright © 2012 SciRes. AiM  Q. LIANG ET AL. Copyright © 2012 SciRes. AiM 207 ogy, Vol. 38, No. 12, 2000, pp. 4453-4458. [15] D. C. Dillon, M. R. Alderson, C. H. Day, T. Bement, A. Campos-Neto, Y. A. W.Skeiky, T. Vedvick, R. Badaro, S. G. Reed and R. Houghton, “Molecular and Immunologi- cal Characterization of Mycobacterium tuberculosis CFP- 10, an Immunodiagnostic Antigen Missing in Mycobacte- rium bovis BCG,” Journal of Clinical Microbiology, Vol. 38, No. 9, 2000, pp. 3285-3290. [16] L. A. H. van Pinxteren, P. Ravn, E. M. Agger, J. Pollock and P. Andersen, “Diagnosis of Tuberculosis Based on the Two Specific Antigens ESAT-6 and CFP10,” Clinical and Vaccine Immunology, Vol. 7, No. 2, 2000, pp. 155- 160. doi:10.1128/CDLI.7.2.155-160.2000 [17] P. S. Renshaw, K. L. Lightbody, V. Veverka, F. W. Muskett, G. Kelly, T. A. Frenkiel, S. V. Gordon, R. G. Hewinson, B. Burke, J. Norman, R. A. Williamson and M. D. Carr, “Structure and Function of the Complex Formed by the Tuberculosis Virulence Factors CFP-10 and ESAT-6,” The EMBO Journal, Vol. 24, No. 23, 2005, pp. 2491-2498. doi:10.1038/sj.emboj.7600732 [18] Y. Hu, F. Movahedzadeh, N. G. Stoker and A. R. M. Coates, “Deletion of the Mycobacterium tuberculosis Al- pha-Crystallin-Like HspX Gene Causes Increased Bacte- rial Growth in Vivo,” Infection and Immunity, Vol. 74, No. 2, 2006, pp. 861-868. doi:10.1128/IAI.74.2.861-868.2006 [19] R. Colangeli, J. S. Spencer, P. Bifani, A. Williams, K. Lyashchenko, M. A. Keen, P. J. Hill, J. Belisle and M. L. Gennaro, “MTSA-10, the Product of the Rv3874 Gene of Mycobacterium tuberculosis, Elicits Tuberculosis-Spe- cific, Delayed-Type Hypersensitivity in Guinea Pigs,” Infection and Immunity, Vol. 68, No. 2, 2000, pp. 990- 993. doi:10.1128/IAI.68.2.990-993.2000 [20] G. H. Bothamley, “Epitope-Specific Antibody Levels De- monstrate Recognition of New Epitopes and Changes in titer but Not Affinity during Treatment of Tuberculosis,” Clinical and Vaccine Immunology, Vol. 11, No. 5, 2004, pp. 942-951. doi:10.1128/CDLI.11.5.942-951.2004 [21] A. Geluk, M. Y. Lin, K. E. Meijgaarden, E. M. S. Leyten, K. L. M. C. Franken, T. H. M. Ottenhoff and M. R. Klein, “T-Cell Recognition of the HspX Protein of Mycobacte- rium tuberculosis Correlates with Latent M. tuberculosis Infection but Not with M. bovis BCG Vaccination,” In- fection and Immunity, Vol. 75, No. 6, 2007, pp. 2914- 2921. doi:10.1128/IAI.01990-06 [22] J. C. Ranford, A. R. Coates and B. Henderson, “Chaper- onins Are Cell-Signalling Proteins: The Unfolding Biol- ogy of Molecular Chaperones,” Molecular Medicine, Vol. 2, No. 8, 2000, pp. 1-17. [23] I. Rosenkrands, K. Weldingh, P. Ravn, L. Brandt, P. Ho- jrup, P. B. Rasmussen, A. R. Coates, M. Singh, P. Mas- cagni and P. Andersen, “Differential T-Cell Recognition of Native and Recombinant Mycobacterium tuberculosis GroES,” Infection and Immunity, Vol. 67, No. 11, 1999, pp. 5552-5558. [24] P. Nunn, “The Global Epidemic. The Present Epidemiol- ogy of Tuberculosis,” Scottish Medical Journal, Vol. 45, No. 5, 2000, pp. 6-7. [25] R. J. Wilkinson, K. Haslov, R. Rappuoli, F. Giovannoni, P. R. Narayanan, C. R. Desai, H. M. Vordermeier, J. Paulsen, G. Pasvol, J. Lvanyi and M. Singh, “Evaluation of the Recombinant 38-Kilodalton Antigen of Mycobac- terium tuberculosis as a Potential Immunodiagnostic Re- agent,” Journal of Clinic Microbiology, Vol. 35, No. 3, 1997, pp. 553-557. [26] K. Lyashchenko, R. Colangeli, M. Houde, H. A. Jahdali, D. Menzies and M. L. Gennaro, “Heterogeneous Anti- body Responses in Tuberculosis,” Infection and Immunity, Vol. 66, No. 8, 1998, pp. 3936-3940. [27] K. R. U. Devi, B. Ramalingam and A. Raja, “Antibody Response to Mycobacterium tuberculosis 30 and 16 kDa Antigens in Pulmonary Tuberculosis with Human Immu- nodeficiency Virus Coinfection,” Diagnostic Microbiol- ogy & Infectious Disease, Vol. 6, No. 3, 2002, pp. 205- 209. [28] G. V. Kanaujia, M. A. Garcia, D. M. Bouley, R. Peters and M. L. Gennaro, “Detection of Early Secretory Anti- genic Target-6 Antibody for Diagnosis of Tuberculosis in Non-Human Primates,” Comparative Medicine, Vol. 53, No. 6, 2003, pp. 602-606. [29] U. Zugel and S. H. E. Kaufmann, “Role of Heat Shock Proteins in Protection from and Pathogenesis of Infec- tious Diseases,” Clinical Microbiology Reviews, Vol. 12, No. 1, 1999, pp. 19-39. [30] M. J. Elhay, T. Oettinger and P. Andersen, “Delayed- Type Hypersensitivity Responses to ESAT-6 and MPT64 from Mycobacterium tuberculosis in the Guinea Pig,” In- fection and Immunity, Vol. 66, No.7, 1998, pp. 3454- 3456. [31] H. Målen, F. S. Berven, K. E. Fladmark and H. G. Wiker, “Comprehensive Analysis of Exported Proteins from My- cobacterium tuberculosis H37Rv,” Proteomics, Vol. 7, No. 10, 2007, pp. 1702-1718. doi:10.1002/pmic.200600853 |