 Advances in Bioscience and Biotechnology, 2012, 3, 609-619 ABB http://dx.doi.org/10.4236/abb.2012.35079 Published Online September 2012 (http://www.SciRP.org/journal/abb/) PUB16 gene expression under abiotic stress and their putative role as an ARM repeat protein in Arabidopsis thaliana self-pollination pathway María Gabriela Acosta1,2, Miguel Ángel Ahumada2,3, Sergio Luis Lassaga2,3, Víctor Hugo Casco1,3 1Microscopy Laboratory Applied to Cell and Molecular Studies, Engineering School, National University of Entre Rios, Oro Verde, Argentina 2Biotechnology Laboratory, Genetics and Plant Breeding Department, INTA-Estación Experimental Agropecuaria Paraná, Oro Verde, Argentina 3Agricultural Sciences School, National University of Entre Rios, Oro Verde, Argentina Email: vcasco@bioingenieira.edu.ar Received 13 June 2012; revised 20 July 2012; accepted 14 August 2012 ABSTRACT The armadillo repeat super-family proteins (ARM repeat super-family proteins) possess tandem arma- dillo repeats and have been postulated to play differ- ent roles in plant development, morphogenesis, de- fense, cell death, and signal transduction through hormone signalling. In The Arabidopsis Information Resource (TAIR), we found 113 loci closely related to ARM repeat family proteins. This extensive group of proteins was studied in flowers tissues by western blot using antibodies directed against the most conserved region of the ARM repeat family proteins. The amino acid residues sequences from TAIR were aligned and the resulting phylogenetic tree allows us to inferring their evolutionary relationships. The main finding was the high similarity between the gene product of PUB16 (At5g01830, A. thaliana) and ARC1 (Brassica napus). In order to search a possible role for PUB16 we carried out stress bioassays using hormonal and saline approaches. Gene expression using RT-PCR showed that some of the ARM repeat super-family proteins are expressed both under salt or hormonal stress conditions. Particularly these studies allowed to detect and semi-quantify PUB16 gene expression in normal or stress growth conditions. In this approach it was revealed that, only in presence of GA, the ex- pression of mRNA-PUB16 became evident. To mor- phologically verify the increasing number of germi- nated pollen grain in gibberellins treated flowers, we used epi-fluorescence microscopy assay. These results suggest that PUB16 may participate in GA signaling pathway favoring self-pollination. Keywords: Self-Pollination; ARM Repeat; Gibberellins 1. INTRODUCTION ARM repeat family proteins are present in animal and plants and they are known to play key roles in several cellular processes including, signal transduction, cy- toskeletal regulation, nuclear import, transcriptional regu- lation, and ubiquitination [1]. This kind of proteins are found in the proteomes of almost all eukaryotic organ- isms and possesses ARM repeat domains, each one are constituted by multiple of 42 amino acid residues. The ARM domain is a highly conserved right handed super helix involved in protein-protein interactions. ARM re- peat domains in plants have evolved as unique domain organizations, such as the U-box and ARM domain com- bination, with specialized functions. The plant-specific U-box/ARM domain proteins are the largest family of ARM repeat proteins in all the genomes surveyed and recent data have implicated these proteins as E3 ubiq- uitin ligases [2]. While functions have not been assigned for most of the plant ARM repeat proteins, recent studies have suggested their importance in multiple processes such as self-incompatibility (SI), hormone signaling and disease resistance [3]. U-box proteins are also involved in very important plant specific pathways [4] such as SI, Pseudo-Self-Compatibility (P-SC) [5]; and abiotic stress responses [6]. Pollination is a crucial step in the life cycle of Angio- sperms, the most important cell-cell interaction in flow- ering plants and this is the mating system adopted by plants species where the pistil is fully developed and composed of stigma, style and ovary [7]. This process is influenced by several factors: stigmas types (i.e. dry stigma or wet stigma) and stigma receptivity (defined as ability to “capture” pollen by adhesion). The appropriate stage of stigma development is crucial for receptivity: on the mature stigma, mature pollen can adhere, hydrate and OPEN ACCESS  M. G. Acosta et al. / Advances in Bioscience and Biotechnology 3 (2012) 609-619 610 germinate. An efficient pollination between pollen grain and pistil is dependent on the ability of the pollen grain to adhere effectively to stigmatic surface [8]. Pollination can be classified in two categories, self- pollination (or autogamy) and cross-pollination (or al- logamy). In nature there is always support for cross-pol- lination because this process ensures the species mainte- nance and contributes to increase the genetic variability in order to provide to species more ability to adapt to new environments [9,10]. To avoid inbreeding and pro- mote out-crossing, many plants have adopted SI systems [11]. In SI plants, pollen will not develop on a stigma that expresses the same S (sterility) alleles as the pollen parent [12]. Members of the Brassicaceae have a dry stigma and one of the interesting features of this trait is the early se- lectivity of pollen capture following pollination [13,14]. Once pollen grains come into contact with stigmatic pa- pillae, only pollen grains recognized as compatible are accepted, thus allowing plants to ignore foreign pollen. These compatible interactions appear to be specific to species within the family, but clearly can occur beyond the species level [15]. For example, success pollinations, as measured by pollen tube penetration into the stigma, have been observed in interspecific and intergeneric crosses in the Brassicaceae [16-18]. Arabidopsis belongs to the Brassicaceae family and therefore has dry stigma with many large unicellular papillae that interact directly with the pollen [19]. The sequential events from pollen adhesion to the path of pollen tube growth through the pistil to the ovule for fertilization have been carefully documented at ultrastructural level in Brassica spp. and Arabidopsis thaliana [20-25]. The best characterized pol- len-pistil interaction on dry stigma is SI response in Bras- sica [13]. Breakdown of the pollination barrier SI in older flowers, a phenomenon known as P-SC or transient SI, has been described as an advantageous reproductive as- surance strategy that allows self-pollination when oppor- tunities for out-crossing have been exhausted [5,26]. The SI phenomenon seems to be controlled sporophytically by a single S locus with multiple alleles or variants and a set of complex dominance relationship between alleles [27- 29]. Its components constitute the male determinant SCR [30-34], the female determinant SLG secreted by the stigma into the cell wall, SRK located in the stigmatic plasma membrane and its ARC1target, also produced in the stigma [19]. ARC1 is a protein required in the Bras- sica pistil for rejection of pollen self-incompatible; it function downstream of the SRK. ARC1 promotes the ubiquitination and proteosomal degradation of compati- bility factors in the pistil, which in turn leads to pollen rejection [35]. ARC1, a positive regulator of Brassica SI protein, was originally identified in a screen for proteins interacting with the active SRK kinase domain and binds to the phosphorylated kinase domain through its ARM domain [36]. The active SRK kinase domain can also cause the re-localization of ARC1 from the cytosol to the ER-associated proteasomes when it is transiently ex- pressed in tobacco BY-2 cells [35]. A second kinase in Brassica SI signalling, the cytoplasmic Ser/Thr protein kinase, designated as M-Locus Protein Kinase (MLPK), also causes the re-localization of ARC1 to the perinuclear region and quite efficiently phosphorylates ARC1 in vitro, suggesting that MLPK may co-regulate ARC1 in con- junction with SRK [37-40]. The proposed model predicts seven ARM domains in ARC1 amino acid residue sequence C-terminal end [35]. ARM repeat super-family proteins shared a conserved three-dimensional structure: tandem ARM repeat form a right-handed super-helix of alpha-helices [1] and they mediate different cellular processes including signal transduction, cytoskeleton regulation, nuclear import, transcriptional regulation and ubiquitination [2]. There are hundreds of eukaryotic proteins with theses tandem structural units. The first member of the gene family to be characterized in detail was the mutant phenotype Drosophila segment polarity gene armadillo [41]. In mammals, their homologue β-catenin, it is known to function in several mechanisms during development, regulating gene expression and cell-cell adhesion [1]. The existence of ARM repeat family proteins in plants may possess very different functions in signal transduc- tion and development, including morphogenesis, defense and cell death, and allow us to predict a mechanism that has been conserved throughout eukaryotic evolution. A large subset of Arabidopsis proteins similar to β-catenin (i.e. ARABIDILLO-1 and -2) contain ARM domain; they are part of the PUB family [1,2,42]. In this work we analyze, compare and predict evolutionary relationship between ARM repeat super-family proteins, present in the Arabidopsis genome [43]. The objective of the pre- sent work was to study the gene expression of PUB16 under abiotic stress and their possible role as an ARM repeat protein in self-pollination pathway. 2. MATERIAL AND METHODS 2.1. Plant Material 2.1.1. Plant Growth Condition Arabidopsis thaliana (L.) wild-type ecotype Columbia-0 [Col-0] plants were grown on a mix of sterile soil (autoclaved before use), vermiculite-perlite and humus. Growth chamber was adjusted to 23˚C and 70% humidity with a 16/8 h light/dark photoperiod under fluorescent illumination supplemented by incandescent light yielding an intensity of 100 - 150 mE/m2·s. The modified Hoag- land solution used to irrigate the plants was done according [44]: (NO3)2Ca·4H2O 0.55 mM; NO3K 0.52 Copyright © 2012 SciRes. OPEN ACCESS  M. G. Acosta et al. / Advances in Bioscience and Biotechnology 3 (2012) 609-619 611 mM; SO4Mg·7H2O 0.22 mM; PO4H2K 0.11 mM; BO3H3 0.046 µM; SO4Zn·7H2O 0.00076 µM; SO4Cu·5H2O 0.00031 µM; Cl2Mn·4H 2O 0.0078 µM; MoO4Na2·2H2O 0.00045 µM; SO4Fe·7H2O 9 mM; NaOH 25 mM. The irrigation frequency was two times weekly with 200 ml of modified Hoagland solution per plate (15 cm × 55 cm) containing 64 pots (one plant/pot). 2.1.2. S tr e ss Induction For the ABA treatments, (100 µM and 200 µM during 2, 4, 8, 11 or 24 hours) 50 open flowers randomly selected from different plants (stage 13 according to [45]), were placed on a Styrofoam disc with small holes containing the ABA solutions. All the setup was placed into a Petri disc. For both ABA concentrations, and the five times assayed, the experiment was replicated twice. The ABA treated flowers were frozen and stored in liquid N2 until use for RNA and proteins extraction. As experimental control, ABA was replaced by vehicle (bdH2O). The GA treatment was done in flowering plants (stage 13 according to [45]) using triple spray every 2 days with 100 µM and 1000 µM of GA3 in controlled chamber growth conditions. The experiment was replicated twice. From each replicate 50 stressed flowers were randomly collected, frozen and stored in liquid N2 until use for RNA and proteins extraction. As experimental control, GA was replaced by vehicle (bdH2O). Salt stresses were performed using 50 mM and 100 mM NaCl for 10 days from rosette of 8 leaves according to [46]. Subsequently, we return to the Hoagland solution irrigation. The experiment was replicated twice. From each replicate, 50 stressed flowers were randomly col- lected, frozen and stored in liquid N2 until use for RNA and proteins extraction. As experimental control, NaCl was replaced by Hoagland solution. 2.2. Identification of ARM Repeats Super-Family Proteins in the Arabidopsis Genome 2.2.1. Se qu ence Anal y s i s Sequences data were obtained from TAIR (The Arabi- dopsis Information Research: http://www.arabidopsis.org/) and BLAST (http://blast.ncbi.nlm.nih.gov/) searches were performed on NCBI (National Center for Biotechnology Investigation). Construction of multiple sequence align- ments were carried out using Clustal XI 2.0 sequence analysis software [47] (http://www.clustal.org/clustal2/). 2.2.2. Secondary Structure Prediction Secondary structure was predicted using programs and database available at website. Domains present were defined by SMART (Simple Modular Architecture Re- search Tool: http://smart.embl-h eidelberg.de/) and Pfam (domains Proteins and families protein database: http://www.sanger.ac.uk/resources/databases/pfam.html) through UniProt (Universal Protein Resource: http://www.uniprot.org/) data base. 2.2.3. Phylogene ti c Analysis Neighbor joining trees were constructed from multiple alignments using NJ plot software http://pbil.u niv-lyon1.fr/software/njplot.html [48]. 2.3. Pollination Assay and Epi-Fluorescence Microscopy For optical and fluorescence microscopy, individual open flowers [floral stages as defined elsewhere [45,49] from fresh wild-type or stressed inflorescences were dissected; outer organs were removed using stainless steel needles under dissection microscope. Pollination tests [50] were performed on 30 pistils, fixed for 1 h in ethanol/acetic acid 3:1 vol/vol]. After washing with distilled water (three times), pistils were softened in 1 N NaOH for 10 min at 65˚C then neutralized in 50 mM phosphate buffer saline (pH 7.5). Finally samples were stained for 2 hours with decolorized aniline blue (Sigma) at 50 mg/ml in 50 mM phosphate buffer saline (pH 7.5) and mounted on slides. As control of the specific stain, non-pollinated stigmas were used. Examination and quantification by epifluorescence microscopy (Olympus BX50) of adhering and penetrating pollen grain in wild-type pistils compared to GA3 treatment was performed using UV light (excita- tion filter 395 nm and emission filter 420 nm). 2.4. RT-PCR Total RNA was isolated using Trizol (Invitrogen). RNA quality and concentration was measured by UV-spec- trometry at 260, 230 and 280 nm, its integrity was checked on 1.5% agarose gel and treated with DNaseI, Rnase Free (Fermentas). First-strand cDNA was synthe- sized from 2 µg of total RNA treated with DNase in 20 µl of reaction volume, using M-MuLV reverse transcript- tase (Fermentas). One-tenth of the first-strand cDNA was used as a template in a 25 µl PCR of 25 cycles (96˚C for 2 min, 94˚C for 1 min, 66.8˚C for 1 min and 72˚C for 1 min) using gene-specific primers. PCR products were analyzed by electrophoresis in 2% agarose gels and visu- alized with UV light before cDNA synthesis (First Strand cDNA Synthesis, Fermentas). β-tubulin (At5g62700) was used as internal control. Primers F1: GAGAATGCTGA TGAGTGCATGG/R1: CAGGGAACCTCAGACAGCA AGT (for Tubulin) and F2: AATCGCCGGGATCAAG CACC/R2: GTGGCGGCGGAAATCTGGAG (for At5g 01830). DNA extraction: 50 mg of tissue was freeze in liquid nitrogen and extensively pulverized using a mi- cropestle. Genomic DNA extraction was carried out using Copyright © 2012 SciRes. OPEN ACCESS  M. G. Acosta et al. / Advances in Bioscience and Biotechnology 3 (2012) 609-619 612 the NucleoSpin® Plant II Genomic DNA kit according to manufacturer’s specifications. To check the genomic DNA integrity a 1.5% agarose gel was prepared [51]. 2.5. Western Blot Total protein extracts from Arabidopsis thaliana were obtained by N2-freezing and grinding 50 mg of floral tissue according to [52]. Total protein concentration was determined in the supernatant by the Bradford’s assay [53], using BSA as standard. Samples were loaded on two standards SDS-PAGE 10% [54], transferred to PVDF membrane and to perform the western blot assay [51]. The primary antibody choice was based on previous evaluation (WebLogo 3.0 software: http://weblogo.three plusone.com/) of the amino acids residues conservation degree in A. thaliana ARM repeats (not shown). Since it exhibits a low-level of amino acid residues conservation, we used an antibody that covers a wide area of ARM repeats. Thus we choose the polyclonal antibody anti- ARMC8 (H-300: sc-98534, Santa Cruz Biotechnology, Inc.) since it recognizes 7 ARM repeats of the 14 present in the protein. ARMC8 is a rabbit polyclonal antibody directed against amino acids residues 311 - 610 mapping within an internal region of ARMC8 of human origin. 2.6. Statistical Analysis Statistical analysis was performed using test pollination results carried out in stressed floral tissue (+GA) and control floral tissue (−GA). The data were analyzed by a one-way analysis of variance using the SAS software (Statistical Analysis Systems, SAS. Institute, Inc., 1999). Results of analysis of variance (ANOVA) and Tukey mean differences test (α = 0.05) was performed for number of pollen grains per stigma in plants with gibber- ellin (+GA) and control plants (−GA). 3. RESULTS AND DISCUSSION Earlier studies of the Arabido psis proteome have found both large number of predict ARM repeat super-family proteins as well as a variety of domain organizations associated with ARM repeats [2,42,55-57]. The largest class of ARM repeats family proteins belong to the PUB proteins and some of them share very similar 3D struc- tures (Figure 1; [2]). As was previously postulated by Mudgil et al. 2004 [42], the fewer number of ARM re- peats observed in AtPUBs and the 3D structural homol- ogy exhibited by PUB16, PUB17 and ARC1 in our stud- ies, may be related with the acquisition of new functions. Most of the ARM repeats family proteins function as E3 ubiquitin-ligases [56] in the regulation of cell death [58] and defense [59] mediating proteasome-dependent deg- radation. This ubiquitin-proteasome pathway is used by GA signaling pathway, like auxin and jasmonate ones, to control gene expression through protein degradation [60, [61]. Since GA behaves as a “florigen” for long-day plants [62] and being a class of hormone involved in the regulation of flower development in Arabidopsis [63], we have analyzed how gene expression in flowering time is affected both under normal growth conditions and GA treatment. As antagonist phytohormone to GA, we de- cided to use ABA, because it is highly linked to the ex- pression of E3 ubiquitin ligases in other models [64,65]. To characterize these ARM repeat super-family pro- teins, we performed western blot technique using ARMC8 as primary antibody. The immunoblot assay allowed us to identify most of A. thaliana ARM repeat proteins, grouped in three very well defined clusters in all stressed plants (30 to 37 kDa; 55 to 60 kDa and 75 to 80 kDa). Contrasting, only two bands were observed both under normal growth conditions (55 to 60 kDa and 75 to 80 kDa) (not shown). In order to determine whether it is a unique protein or a cluster, we compare the molecular weights of 113 loci for putative polypeptides available in TAIR website. Accordingly, the candidates were grouped into 3 major molecular weight clusters: group A, between 75 to 80 kDa; group B, between 55 to 60 kDa; and group C, be- tween 30 to 37 kDa. Group A corresponds to five puta- tive ARM repeat proteins: AT5G01830, AT1G60190, AT5G67340, AT4G31520 and AT4G36550; group B corresponds to the six putative ARM repeat proteins: AT1G23940, AT4G31890, AT5G50900, AT5G22820, AT2G45720 and AT3G1518; and group C corresponds to eight putative ARM repeat proteins: AT3G43260, AT4G15830, AT5G11550, AT3G58180, AT1G08315, AT5G14510, AT3G01450 and AT1G15165. From this study we can remark two important facts: first of all the expression of ARM repeat proteins from groups A and B were increased in stressed conditions and second, the appearance of a new band corresponding to the group C in stressed experiments. From these results we can con- clude that the group C corresponds to ARM repeat pro- teins expressed specifically in floral tissue under the stress conditions evaluated. Following was performed a bioinformatics search in order to decide which of the three putative ARM repeat protein groups must be focused for gene expression studies. The alignment of 113 loci for putative polypep- tides available in TAIR website was performed by com- paring their amino acid residue sequence with the ARC1 ones. This comparison yielded 57 final candidates, 19 of which have U-box sequences in addition to the ARM sequences. Within this group, there are two protein se- quences closely related to ARC1: AT1G29340 (PUB17) and AT5g01830 (PUB16), been the very well-known PUB17, the most closely related to ARC1, followed by Copyright © 2012 SciRes. OPEN ACCESS  M. G. Acosta et al. / Advances in Bioscience and Biotechnology 3 (2012) 609-619 Copyright © 2012 SciRes. 613 Figure 1. 3D structure and evolutionary relationship predictions of ARM repeat proteins. At the left: PyMol molecular visualization tool where can be seen similarities in the spatial C-terminal ARM re- peats folding pattern of: ACRE276, ARC1, PUB16 and PUB17 (Acosta 2010). At the right: is shown the phylogenetic tree resulting from the evolutionary relationship between them. We used the soft- ware available online iTol (Interactive tree of life [http://itol.embl.de/upload.cgi]) [77]. The Neighbour Joining algorithm has been used to allow tree construction and indicated in each node the bootstrap values. Evolutionary distance is shown in the upper right. and their possible function has been inferred [59], we decided to study PUB16, which according to the align- ment and phylogenetic analysis is the other ARC1-like A. thaliana ARM repeat protein. PUB16. PUB17 function as putative E3 ubiquitin ligase contains four ARM repeats and a U-box domain and it was widely studied. Its functional tobacco homolog ACRE276 is required for cell death and defense in So- lanaceae [59]. The BLASTp alignment ARC1/PUB17 produced 58% identity and 74% similarity (E-value = 0) and ARC1/PUB16 display 34% identity and 51% simi- larity (E-value = 1e−75). Similarly, the scores obtained from ClustalX were: PUB16/ARC1: 31; ACRE276/ PUB16: 35; PUB17/PUB16: 36; ACRE276/ARC1: 53; PUB17/ARC1: 60 and ACRE276/PUB17: 68 (Figure 2). PUB16 contains three ARM repeats and one U-box do- main, so we can infer that it could functions as E3 ubiq- uitin-ligase. However, similarity at the amino acid resi- due does not allowed us to deduce similar functions in pollen-stigma recognition mechanism in B. napus and A. thaliana. PUB16 could belong to the ARM repeat proteins from the group A detected on western blot and their hypo- thetical characterization was performed using the Uni- Prot database (http://www.uniprot.o rg/) which freely provides accessible resource of protein sequence and functional information. The five sequences belonging to the group A (AT4G31520, AT5G01830, AT1G60190, AT5G67340 and AT4G36550) have been evidenced only at transcriptional level. According to this result, we can- not establish which of the five possible peptides corre- spond to the expression band observed in our western blot results and should be clarified in future experiments. Therefore, as a first step, we decided to start the stud- ies evaluating the PUB16 transcription levels in normal and different stress conditions at the same flowering stage. Expression studies using RT-PCR technique for subsequent semi-quantification of PUB16 revealed that in presence of GA3 1000 μM, there is a significant gene expression of this molecule; while there is no expression PUB17 is an E3 ubiquitin ligase and has been postulated that it may form a signaling complex with a SRK1-like kinase analogous to the SRK/ARC1/thioredoxin complex in B. napus during rejection of self-incompatible pollen in Brassica [36,66]. Because PUB17 has already been fully characterized OPEN ACCESS 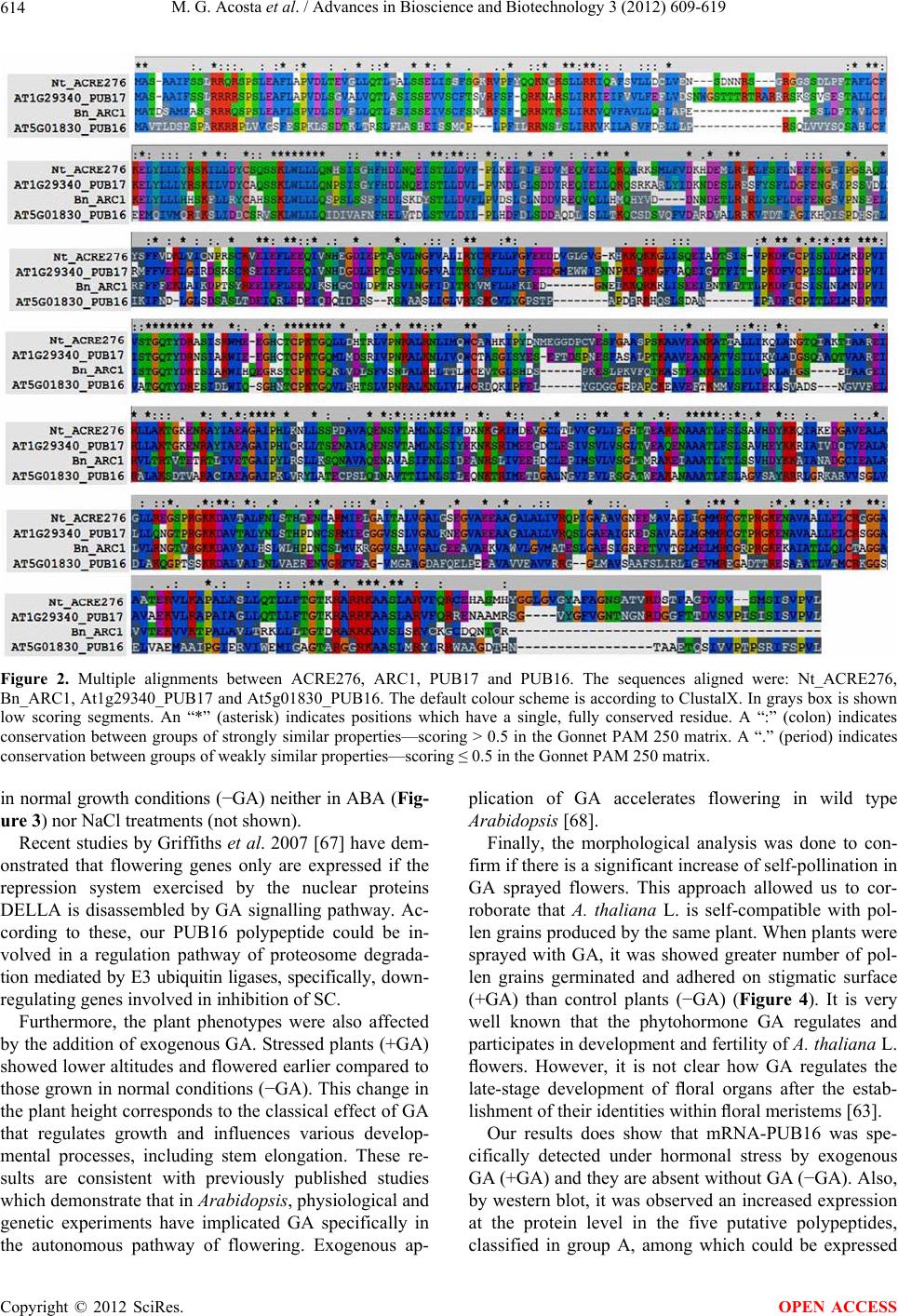 M. G. Acosta et al. / Advances in Bioscience and Biotechnology 3 (2012) 609-619 614 Figure 2. Multiple alignments between ACRE276, ARC1, PUB17 and PUB16. The sequences aligned were: Nt_ACRE276, Bn_ARC1, At1g29340_PUB17 and At5g01830_PUB16. The default colour scheme is according to ClustalX. In grays box is shown low scoring segments. An “*” (asterisk) indicates positions which have a single, fully conserved residue. A “:” (colon) indicates conservation between groups of strongly similar properties—scoring > 0.5 in the Gonnet PAM 250 matrix. A “.” (period) indicates conservation between groups of weakly similar properties—scoring ≤ 0.5 in the Gonnet PAM 250 matrix. in normal growth conditions (−GA) neither in ABA (Fig- ure 3) nor NaCl treatments (not shown). Recent studies by Griffiths et al. 2007 [67] have dem- onstrated that flowering genes only are expressed if the repression system exercised by the nuclear proteins DELLA is disassembled by GA signalling pathway. Ac- cording to these, our PUB16 polypeptide could be in- volved in a regulation pathway of proteosome degrada- tion mediated by E3 ubiquitin ligases, specifically, down- regulating genes involved in inhibition of SC. Furthermore, the plant phenotypes were also affected by the addition of exogenous GA. Stressed plants (+GA) showed lower altitudes and flowered earlier compared to those grown in normal conditions (−GA). This change in the plant height corresponds to the classical effect of GA that regulates growth and influences various develop- mental processes, including stem elongation. These re- sults are consistent with previously published studies which demonstrate that in Arabidopsis, physiological and genetic experiments have implicated GA specifically in the autonomous pathway of flowering. Exogenous ap- plication of GA accelerates flowering in wild type Arabidopsis [68]. Finally, the morphological analysis was done to con- firm if there is a significant increase of self-pollination in GA sprayed flowers. This approach allowed us to cor- roborate that A. thaliana L. is self-compatible with pol- len grains produced by the same plant. When plants were sprayed with GA, it was showed greater number of pol- len grains germinated and adhered on stigmatic surface (+GA) than control plants (−GA) (Figure 4). It is very well known that the phytohormone GA regulates and participates in development and fertility of A. thaliana L. flowers. However, it is not clear how GA regulates the late-stage development of floral organs after the estab- lishment of their identities within floral meristems [63]. Our results does show that mRNA-PUB16 was spe- cifically detected under hormonal stress by exogenous GA (+GA) and they are absent without GA (−GA). Also, by western blot, it was observed an increased expression at the protein level in the five putative polypeptides, classified in group A, among which could be expressed Copyright © 2012 SciRes. OPEN ACCESS 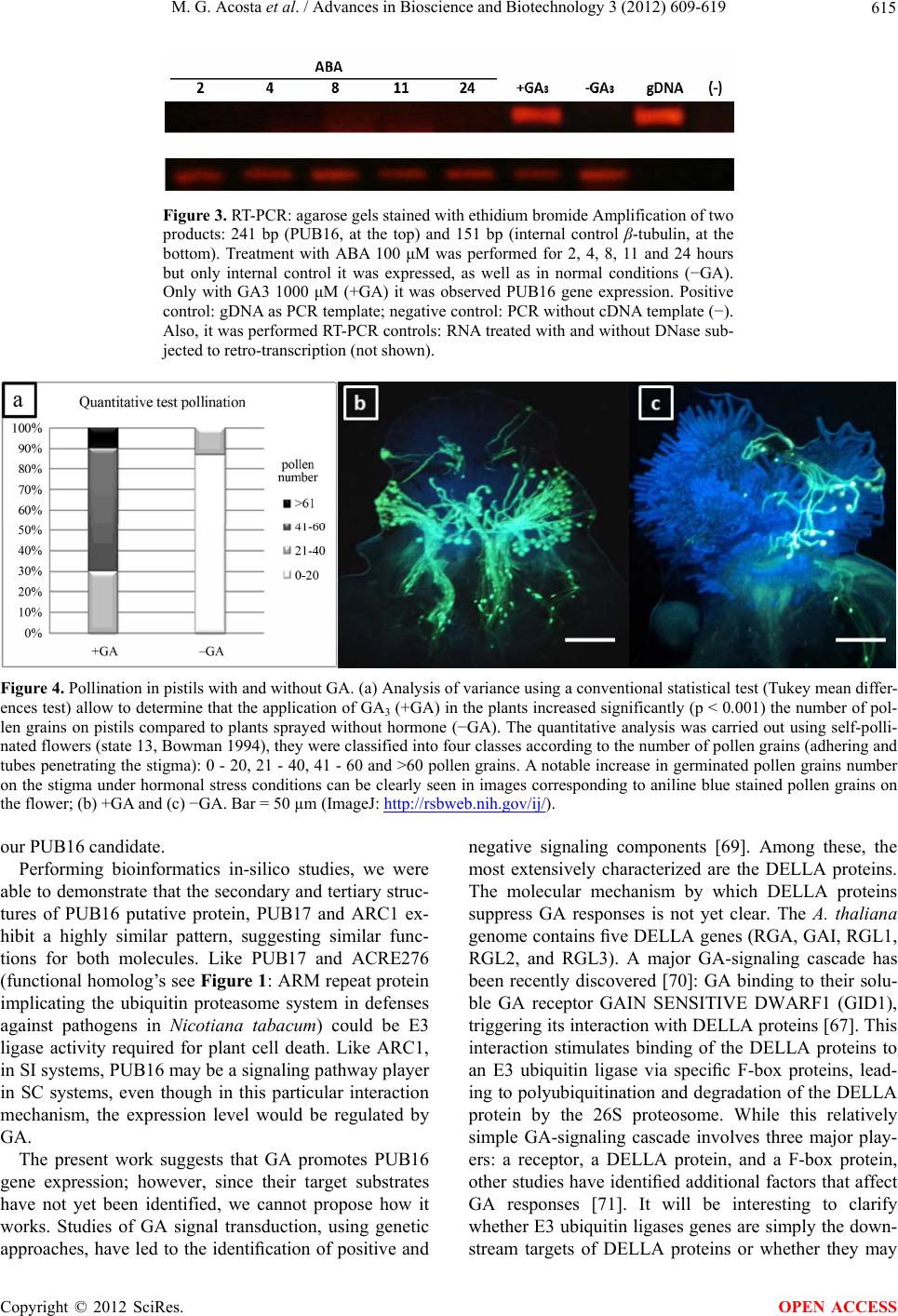 M. G. Acosta et al. / Advances in Bioscience and Biotechnology 3 (2012) 609-619 615 Figure 3. RT-PCR: agarose gels stained with ethidium bromide Amplification of two products: 241 bp (PUB16, at the top) and 151 bp (internal control β-tubulin, at the bottom). Treatment with ABA 100 μM was performed for 2, 4, 8, 11 and 24 hours but only internal control it was expressed, as well as in normal conditions (−GA). Only with GA3 1000 μM (+GA) it was observed PUB16 gene expression. Positive control: gDNA as PCR template; negative control: PCR without cDNA template (−). Also, it was performed RT-PCR controls: RNA treated with and without DNase sub- jected to retro-transcription (not shown). Figure 4. Pollination in pistils with and without GA. (a) Analysis of variance using a conventional statistical test (Tukey mean differ- ences test) allow to determine that the application of GA3 (+GA) in the plants increased significantly (p < 0.001) the number of pol- len grains on pistils compared to plants sprayed without hormone (−GA). The quantitative analysis was carried out using self-polli- nated flowers (state 13, Bowman 1994), they were classified into four classes according to the number of pollen grains (adhering and tubes penetrating the stigma): 0 - 20, 21 - 40, 41 - 60 and >60 pollen grains. A notable increase in germinated pollen grains number on the stigma under hormonal stress conditions can be clearly seen in images corresponding to aniline blue stained pollen grains on the flower; (b) +GA and (c) −GA. Bar = 50 µm (ImageJ: http://rsbweb.nih.gov/ij/). our PUB16 candidate. Performing bioinformatics in-silico studies, we were able to demonstrate that the secondary and tertiary struc- tures of PUB16 putative protein, PUB17 and ARC1 ex- hibit a highly similar pattern, suggesting similar func- tions for both molecules. Like PUB17 and ACRE276 (functional homolog’s see Figure 1: ARM repeat protein implicating the ubiquitin proteasome system in defenses against pathogens in Nicotiana tabacum) could be E3 ligase activity required for plant cell death. Like ARC1, in SI systems, PUB16 may be a signaling pathway player in SC systems, even though in this particular interaction mechanism, the expression level would be regulated by GA. The present work suggests that GA promotes PUB16 gene expression; however, since their target substrates have not yet been identified, we cannot propose how it works. Studies of GA signal transduction, using genetic approaches, have led to the identification of positive and negative signaling components [69]. Among these, the most extensively characterized are the DELLA proteins. The molecular mechanism by which DELLA proteins suppress GA responses is not yet clear. The A. thaliana genome contains five DELLA genes (RGA, GAI, RGL1, RGL2, and RGL3). A major GA-signaling cascade has been recently discovered [70]: GA binding to their solu- ble GA receptor GAIN SENSITIVE DWARF1 (GID1), triggering its interaction with DELLA proteins [67]. This interaction stimulates binding of the DELLA proteins to an E3 ubiquitin ligase via specific F-box proteins, lead- ing to polyubiquitination and degradation of the DELLA protein by the 26S proteosome. While this relatively simple GA-signaling cascade involves three major play- ers: a receptor, a DELLA protein, and a F-box protein, other studies have identified additional factors that affect GA responses [71]. It will be interesting to clarify whether E3 ubiquitin ligases genes are simply the down- stream targets of DELLA proteins or whether they may Copyright © 2012 SciRes. OPEN ACCESS  M. G. Acosta et al. / Advances in Bioscience and Biotechnology 3 (2012) 609-619 616 also interact with DELLA proteins as region-specific cofactors. Interactions studies have been used to identify candidates [72] and in vitro ubiquitination assays can be used to confirm the ability of E3 ligases to ubiquitinate these potential substrates [73-75]. However, at present, have not been identified yet the ARC1 potential targets. Additionally, with the identification of the biological roles of putative PUB proteins, the understanding about how these E3 ligases are activated is an equally impor- tant step in the elucidation of their physiological func- tions. 4. CONCLUTIONS ARM repeat super-family proteins, like related proteins possessing this domain, may be involved in protein-pro- tein interactions. The ARM repeat super-family proteins analysis in this plant model, will allow a better under- standing of the pollination cell biology and its possible participation in their signaling pathways. Since there is very little knowledge about GA signaling pathway, even though they are proposed to be related to plants fertility, it is very challenging to study which genes are expressed under both stress or normal growth conditions, and also it is important to temporal and functionally characterize their behavior. In this work, sequence comparison revealed significant structural homology between ARC1 and PUB16. This close relationship allows us to infer that similar transduc- tion pathways might exist in two Brassicaceae species differently involved in pollen hydration regulation and their signaling mechanism of self and non-self pollen recognition. Also, the results obtained here show the ARM repeat super-family proteins clustering of A. thaliana which is composed of three groups according to molecular weights. According to our gene expression studies, it appears that exogenous addition of GA cause PUB16 gene expression, but this not happen in normal condition growth or in ABA presence. Also, these results are consistent with previous reports that postulate that, plants ARM repeat super-family proteins expression is subjected to both salt [50] or hormonal stresses [76]. Furthermore, these outcomes suggest that A. thaliana could use some GA-signaling pathway which favors self- pollination, fruit set and great seed production under hormonal stress. Further analysis of ARM repeat super- family proteins will improve understanding of their bi- ology role related to its possible involvement in different signaling pathways. REFERENCES [1] Coates, J.C. (2007) Plant cell monographs: Plant growth signaling. Springer, Berlin, 299-314. [2] Samuel, M., Salt, J., Shiu, S. and Goring, D. (2006) Mul- tifunctional arm repeat domains in plants. International Review of Cytology, 253, 1-26. doi:10.1016/S0074-7696(06)53001-3 [3] Coates, J.C., Laplaze, L. and Haseloff, J. (2006) Arma- dillo-related proteins promote lateral root development in Arabidopsis. Proceedings of the National Academy of Sciences USA, 103, 1621-1626. doi:10.1073/pnas.0507575103 [4] González-Lamothe, R., Tsitsigiannis, D.I., Ludwig, A.A., Panicot, M., Shirasu, K. and Jones, J.D.G. (2006) The U-box protein CMPG1 is required for efficient activation of defense mechanisms triggered by multiple resistance genes in tobacco and tomato. The Plant Cell, 18, 1067- 1083. doi:10.1105/tpc.106.040998 [5] Liu, P., Sherman-Broyles, S. and Nasrallah, M.E. (2007) A cryptic modifier causing transient self-incompatibility in Arabidopsis thaliana. Current Biology, 17, 734-740. doi:10.1016/j.cub.2007.03.022 [6] Yan, J., Wang, J., Li, Q., Hwang, J.R., Patterson, C. and Zhang, H. (2003) AtCHIP, a U-box-containing E3 ubiq- uitin ligase, plays a critical role in temperature stress tol- erance in Arabidopsis. Plant Physiology, 132, 861-869. doi:10.1104/pp.103.020800 [7] Dong, J., Kim, S.T. and Lord, E.M. (2005) Plantacyanin plays a role in reproduction in Arabidopsis. Plant Physi- ology, 138,778-789. doi:10.1104/pp.105.063388 [8] Wheeler, M.J., Franklin-Tong, V.E. and Franklin, F.C.H. (2001) The molecular and genetic basis of pollen-pistil interactions. New Phytologist, 151, 565-584. doi:10.1046/j.0028-646x.2001.00229.x [9] Nasrallah, M.E., Liu, P., Sherman-Broyles, S., Boggs, N.A. and Nasrallah, J.B. (2004) Natural variation in ex- pression of self-incompatibility in Arabidopsis thaliana: Implications for the evolution of selfing. Proceedings of the National Academy of Sciences USA, 101, 16070- 16074. doi:10.1073/pnas.0406970101 [10] Sanchez, A.M., Bosch, M., Bots, M., Nieuwland, J., Feron, R. and Mariani, C. (2004) Pistil factors controlling pollination. The Plant Cell, 16, S98-S106. doi:10.1105/tpc.017806 [11] Huang, J., Zhao, L., Yang, Q. and Xue, Y. (2006) AhSSK1, a novel SKP1-like protein that interacts with the S-locus F-box protein SLF. The Plant Journal, 46, 780-793. doi:10.1111/j.1365-313X.2006.02735.x [12] Kusaba, M., Dwyer, K., Hendershot, J., Vrebalov, J., Nasrallah, J.B. and Nasrallah, M.E. (2001) Self-incom- patibility in the genus Arabidopsis: Characterization of the S locus in the outcrossing A. lyrata and its autoga- mous relative A. thaliana. The Plant Cell, 13, 627-643. [13] Heslop-Harrison, Y. and Shivanna, K.R. (1977) The re- ceptive surface of the angiosperm stigma. Annals of Bot- any, 41, 1233-1258. [14] Dickinson, H. (1995) Dry stigmas, water and self-in- compatibility in Brassica. Sexual Plant Reproduction, 8, 1-10. doi:10.1007/BF00228756 [15] Hulskamp, M., Kopczak, S.D., Horejsi, T.F., Kihl, B.K. and Pruitt, R.E. (1995) Identification of genes required for pollen-stigma recognition in Arabidopsis thaliana. Copyright © 2012 SciRes. OPEN ACCESS  M. G. Acosta et al. / Advances in Bioscience and Biotechnology 3 (2012) 609-619 617 The Plant Journal, 8, 703-714. doi:10.1046/j.1365-313X.1995.08050703.x [16] Sampson, D.R. (1962) Intergeneric pollen-stigma incom- patibility in Cruciferae. Canadian Journal of Genetics and Cytology, 4, 38-49. [17] Hiscock, S.J. and Dickinson, H.G. (1993) Unilateral in- compatibility within the Brassicaceae: Further evidence for the involvement of the self incompatibility (S)-locus. Theoretical and Applied Genetics, 86, 744-753. doi:10.1007/BF00222665 [18] Lelivelt, C.L.C. (1993) Studies of pollen grain germina- tion, pollen-tube growth, micropylar penetration and seed set in intraspecific and intergeneric crosses within three Cruciferae species. Euphytica, 67, 185-197. doi:10.1007/BF00040620 [19] Nasrallah, J.B. (2000) Cell-cell signaling in the self-in- compatibility response. Current Opinion of Plant Biology, 3, 368-373. doi:10.1016/S1369-5266(00)00098-4 [20] Hill, J.P. and Lord, E.M. (1987) Dynamics of pollen tube growth in the wild radish, Raphanus raphanistrum (Bras- sicaceae) 2.Morphology, cytochemistry, and ultrastruc- ture of transmitting tissues, and path of pollen tube growth. American Journal of Botany, 74, 988-997. doi:10.2307/2443938 [21] Elleman, C.J. and Dickinson, H.G. (1990) The role of the exine coating in pollen-stigma interactions in Brassica oleracea L. New Phytologist, 114, 511-518. doi:10.1111/j.1469-8137.1990.tb00419.x [22] Elleman, C.J., Franklin-Tong, V. and Dickinson, H.G. (1992) Pollination in species with dry stigmas: The nature of the early stigmatic response and the pathway taken by pollen tubes. New Phytologist, 121, 413-424. doi:10.1111/j.1469-8137.1992.tb02941.x [23] Kandasamy, M.K., Nasrallah, J.B. and Nasrallah, M.E. (1994) Pollen-pistil Interactions and developmental regu- lation of pollen tube growth in Arabidopsis. Development, 120, 3405-3418. [24] Hulskamp, M., Schneitz, K. and Pruitt, R.E. (1995b) Ge- netic evidence for along-range activity that directs pollen tube guidance in Arabidopsis. The Plant Cell, 7, 57-64. [25] Lennon, K.A., Roy, S., Hepler, P.K. and Lord, E.M. (1998) The structure of the transmitting tissue of Arabi- dopsis thaliana (L.) and the path of pollen tube growth. Sexual Plant Reproduction, 11, 49-59. doi:10.1007/s004970050120 [26] Tsuchimatsu, T., Suwabe, K., Shimizu-Inatsugi, R., Iso- kawa, S., Pavlidis, P., Stadler, T., Suzuki, G., Takayama, S., Watanabe, M. and Shimizu, K. (2010) Evolution of self-compatibility in Arabidopsis by a mutation in the male specificity gene. Nature, 464, 1342-1346. doi:10.1038/nature08927 [27] Bateman, A.J. (1955) Self-incompatibility systems in an- giosperms. III. Cruciferae. Heredity, 9, 52-68. doi:10.1038/hdy.1955.2 [28] Thompson, K.F. and Taylor, J.P. (1966) Non-linear domi- nance relationships between S-alleles. Heredity, 21, 345- 362. doi:10.1038/hdy.1966.36 [29] Tarutani, Y., Shiba, H., Iwano, M., Kakizaki, T., Suzuki, G., Watanabe, M., Isogai, A. and Takayama, S. (2010) Trans-acting small RNA determines dominance relation- ships in Brassica self-incompatibility. Nature, 466, 983- 986. doi:10.1038/nature09308 [30] Suzuki, G., Kai, N., Hirose, T., Fukui, K., Nishio, T., Takayama, S., Isogai, A., Watanabe, M. and Hinata, K. (1999) Genomic organization of the S locus: Identifica- tion and characterization of genes in SLG/SRK region of S9 haplotype of Brassica campestris (syn. rapa). Genetics, 153, 391-400. [31] Schopfer, C.R., Nasrallah, M.E. and Nasrallah, J.B. (1999) The male determinant of self-incompatibility in Brassica. Science, 286, 1697-1700. doi:10.1126/science.286.5445.1697 [32] Takayama, S., Shiba, H., Iwano, M., Shimosato, H., Che, F.S., Kai, N., Watanabe, M., Suzuki, G., Hinata, K. and Isogai, A. (2000) The pollen determinant of self-incom- patibility in Brassica campestris. Proceedings of the Na- tional Academy of Sciences USA, 97, 1920-1925. [33] Takayama, S., Shimosato, H., Shiba, H., Funato, M., Che, F.S., Watanabe, M., Iwano, M. and Isogai, A. (2001) Di- rect ligand-receptor complex interaction controls Bras- sica self-incompatibility. Nature, 413, 534-538. doi:10.1038/35097104 [34] Shiba, H., Takayama, S., Iwano, M., Shimosato, H., Fu- nato, M., Nakagawa, T., Che, F., Suzuki, G., Watanabe, M., Hinata, K. and Isogai, A. (2001) A pollen coat pro- tein, SP11/SCR, determines the pollen S-specificity in the self-incompatibility of Brassica species. Plant Physiology, 125, 2095-2103. doi:10.1104/pp.125.4.2095 [35] Stone, S.L., Anderson, E.M., Mullen, R.T. and Goring, D.R. (2003) ARC1 is an E3 ubiquitin ligase and promotes the ubiquitination of proteins during the rejection of self- incompatible Brassica pollen. The Plant Cell, 15, 885- 898. doi:10.1105/tpc.009845 [36] Gu, T.S., Mazzurco, M., Sulaman, W., Matias, D.D. and Goring, D.R. (1998) Binding of an arm repeat protein to the kinase domain of the S-locus receptor kinase. Pro- ceedings of the National Academy of Sciences USA, 95, 382-387. [37] Samuel, M.A., Mudgil, Y., Salt, J.N., Delmas, F., Ramachandran, S., Chilelli, A. and Goring, D.R. (2008) Interactions between the S-domain receptor kinases and AtPUB-ARM E3 ubiquitin ligase ssuggest a conserved signaling pathway in Arabidopsis. Plant Physiology, 147, 2084-2095. doi:10.1104/pp.108.123380 [38] Murase, K., Shiba, H., Iwano, M., Che, F.S., Watanabe, M., Isogai, A. and Takayama, S. (2004) A membrane- anchored protein kinase involved in Brassica self-in- compatibility signaling. Science, 303, 1516-1519. doi:10.1126/science.1093586 [39] Kakita, M., Murase, K., Iwano, M., Matsumoto, T., Wa- tanabe, M., Shiba, H., Isogai, A. and Takayama, S. (2007) Two distinct forms of M-locus protein kinase localize to the plasma membrane and interact directly with S-locus receptor kinase to transducer self-incompatibility signal- ing in Brassica rapa. The Plant Cell, 19, 3961-3973. doi:10.1105/tpc.106.049999 [40] Kakita, M., Shimosato, H., Murase, K., Isogai, A. and Copyright © 2012 SciRes. OPEN ACCESS  M. G. Acosta et al. / Advances in Bioscience and Biotechnology 3 (2012) 609-619 618 Takayama, S. (2007) Direct interaction between the S- locus receptor kinase and M locus protein kinase in- volved in Brassica self-incompatibility signaling. Plant Biotechnology, 24, 185-190. doi:10.5511/plantbiotechnology.24.185 [41] Riggleman, B., Wieschaus, E. and Schedl, P. (1989) Mo- lecular analysis of the armadillo locus: Uniformly distrib- uted transcripts and a protein with novel internal repeats are associated with a Drosophila segment polarity gene. Genes Development, 3, 96-113. doi:10.1101/gad.3.1.96 [42] Mudgil, Y., Shiu, S.H., Stone, S.L., Salt, J.N. and Goring, D.R. (2004) A large complement of the predicted Arabi- dopsis ARM repeat proteins are members of the U-box E3 ubiquitin ligase family. Plant Physiology, 134, 59-66. doi:10.1104/pp.103.029553 [43] Acosta, M.G., Langhi, D., Lassaga, S.L. and Casco, V.H. (2010a) Bioinformatics and morphological studies of pol- lination mechanism as a process of cell-cell adhesion in Arabidopsis thaliana. Agricultural Science Magazine, 14, 3-15. [44] Epstein, E. (1972) Mineral nutrition of plants: Principles and perspectives. J. Wiley and Sons, Inc., New York, 68- 82. [45] Bowman, J. (1994) Arabidopsis: An atlas of morphology and development. Springer-Verlag, New York, 258-259. [46] Banzai, T., Hershkovits, G., Katcoff, D.J., Hanagata, N., Dubinsky, Z. and Karube, I. (2002) Identification and characterization of mRNA transcripts differentially ex- pressed in response to high salinity by means of differen- tial display in the mangrove, Bruguiera gymnorrhiza. Plant Science, 162, 499-505. doi:10.1016/S0168-9452(01)00601-X [47] Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994) CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22, 4673-4680. doi:10.1093/nar/22.22.4673 [48] Perriere, G. and Gouy, M. (1996) www-query: An on-line retrieval system for biological sequence banks. Biochimie, 78, 364-369. doi:10.1016/0300-9084(96)84768-7 [49] Smyth, D.R., Bowman, J.L. and Meyerowitz, E.M. (1990) Early flower development in Arabidopsis. The Plant Cell, 2, 755-767. [50] Eschrich, W. and Currier, H.B. (1964) Identification of callose by it diachrome and fluorchrome reactions. Stain Technology, 39, 308-309. [51] Sambrook, D.W. and Russell, J. (2001) Molecular clon- ing: A laboratory manual. Cold Spring Harbor Laboratory Press, New York, 102-114. [52] Weigel, D. (2002) Arabidopsis: A laboratory manual. Cold Spring Harbor Laboratory Press, New York, 102- 145. [53] Bradford, M.M. (1976) Rapid and sensitive method for the quantitation of microgram quantities of protein utiliz- ing the principle of protein-dye binding. Analytical Bio- chemistry, 72, 248-254. doi:10.1016/0003-2697(76)90527-3 [54] Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227, 680-685. doi:10.1038/227680a0 [55] Azevedo, C., Santos-Rosa, M.J. and Shirasu, K. (2001) The U-box protein family in plants. Trends in Plant Sci- ence, 6, 354-358. doi:10.1016/S1360-1385(01)01960-4 [56] Andersen, P., Kragelund, B.B., Olsen, A.N., Larsen, F.H., Chua, N., Poulsen, F.M. and Skriver, K. (2004) Structure and biochemical function of aprototypical Arabidopsis U-box domain. Journal of Biological Chemistry, 279, 40053-40061. doi:10.1074/jbc.M405057200 [57] Wiborg, J., O’Shea, C. and Skriver, K. (2008) Biochemi- cal function of typical and variant Arabidopsis thaliana U-box E3 ubiquitin-protein ligases. Biochemical Journal, 413, 447-457. doi:10.1042/BJ20071568 [58] Thomas, S.G. and Franklin-Tong, V.E. (2004) Self-in- compatibility triggers programmed cell death in Papaver pollen. Nature, 429, 305-309. doi:10.1038/nature02540 [59] Yang, C.W., González-Lamothe, R., Ewan, R.A., Row- land, O., Yoshioka, H., Shenton, M., Ye, H., O’Donnell, E., Jones, J.D.G. and Sadanandom, A. (2006) The E3 ubiquitin ligase activity of Arabidopsis PLANTU-BOX17 and its functional tobacco homolog ACRE276 are re- quired for cell death and defense. The Plant Cell, 18, 1084-1098. doi:10.1105/tpc.105.039198 [60] Itoh, H., Ueguchi-Tanaka, M., Sato, Y., Ashikari, M. and Matsuoka, M. (2002) The gibberellin signaling pathway is regulated by the appearance and disappearance of SLENDER RICE1 in nuclei. The Plant Cell, 14, 57-70. doi:10.1105/tpc.010319 [61] Spartz, A.K., Lee, S.H., Wenger, J.P., Gonzalez, N., Itoh, H., Inzé, D., Peer, W.A., Murphy, A.S., Overvoorde, P.J. and Gray, W.M. (2012) The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. The Plant Journal, 70, 978-990. doi:10.1111/j.1365-313X.2012.04946.x [62] Komeda, Y. (2004) Genetic regulation of time to flower- ing Arabidopsis thaliana. Annual Review of Plant Biol- ogy, 55, 521-535. doi:10.1146/annurev.arplant.55.031903.141644 [63] Yu, H., Ito, T., Wellmer, F. and Meyerowitz, E.M. (2004) Floral homeotic genes are targets of gibberellins signaling in flower. Proceedings of the National Academy of Sci- ences USA, 101, 7827-7832. [64] Weiss, D. and Ori, N. (2007) Mechanisms of cross talk between gibberellin and other hormones. Plant Physiol- ogy, 144, 1240-1246. doi:10.1104/pp.107.100370 [65] Acosta, M.G., Ahumada, M.A., Lassaga, S.L. and Casco, V.H. (2010b) Abiotic stress effect on gene expression of ARM repeats proteins in A. thaliana. Book of Abstracts— XXIII Meeting Argentina Plant Physiology, 1, 205. [66] Cabrillac, D., Cock, J.M., Dumas, C. and Gaude, T. (2001) The S locus receptor kinase is inhibited by thioredoxins and activated by pollen coat proteins. Nature, 410, 220- 223. doi:10.1038/35065626 [67] Griffiths, J., Murase, K., Rieu, I., Zentella, R., Zhang, Z.L., Powers, S.J., Gong, F., Phillips, A.L., Hedden, P. and Sun, T.P. (2007) Genetic characterization and func- Copyright © 2012 SciRes. OPEN ACCESS  M. G. Acosta et al. / Advances in Bioscience and Biotechnology 3 (2012) 609-619 Copyright © 2012 SciRes. 619 OPEN ACCESS tional analysis of the GID1 gibberellin receptors in Arabi- dopsis. The Plant Cell, 18, 3399-3414. doi:10.1105/tpc.106.047415 [68] Blazquez, M.A., Green, R., Nilsson, O., Sussman, M.R. and Weigel, D. (1998) Gibberellins promote flowering of Arabidopsis by activating the LEAFY promoter. The Plant Cell, 10, 791-800. [69] Sun, T.P. and Gubler, F. (2004) Molecular mechanism of gibberellin signaling in plants. Annual Review of Plant Biology, 55, 197-223. doi:10.1146/annurev.arplant.55.031903.141753 [70] Ueguchi-Tanaka, M., Ashikari, M., Nakajima, M., Itoh, H., Katoh, E., Kobayashi, M., Chow, T.Y., Hsing, Y.I., Kitano, H. and Yamaguchi, I. (2005) GIBBERELLIN INSENSITIVE DWARF1 encodes a soluble receptor for gibberellin. Nature, 437, 693-698. doi:10.1038/nature04028 [71] Hartweck, L.M. and Olszewski, N.E. (2006) GIBBEREL- LIN INSENSITIVE DWARF1 is a gibberellin receptor that illuminates and raises questions about GA signaling. The Plant Cell, 18, 278-282. doi:10.1105/tpc.105.039958 [72] Yee, D. and Goring, D.R. (2009) The diversity of PUB E3 ubiquitin ligases: From upstream activators to down- stream target substrates. Journal of Experimental Botany, 60, 1109-1121. doi:10.1093/jxb/ern369 [73] Cho, S.K., Chung, H.S., Ryu, M.Y., Park, M.J., Lee, M.M., Bahk, Y.Y., Kim, J., Pai, H.S. and Kim, W.T. (2006) Heterologous expression and molecular and cellular characterization of CaPUB1 encoding a hot pepper U- BoxE3 ubiquitin ligase homolog. Plant Physiology, 142, 1664-1682. doi:10.1104/pp.106.087965 [74] Cho, S.K., Ryu, M.Y., Song, C., Kwak, J.M. and Kim, W.T. (2008) Arabidopsis PUB22 and PUB23 are ho- mologous U-box E3 ubiquitin ligases that play combina- tory roles in response to drought stress. The Plant Cell, 20, 1899-1914. doi:10.1105/tpc.108.060699 [75] Shen, G., Yan, J., Pasapula, V., Luo, J., He, C., Clarke, A.K. and Zhang, H. (2007) The chloroplast protease sub- unit ClpP4 is a substrate of the E3 ligase AtCHIP and plays an important role in chloroplast function. The Plant Journal, 49, 228-237. doi:10.1111/j.1365-313X.2006.02963.x [76] Alonso-Ramírez, A., Rodríguez, D., Reyes, D., Jiménez, J.A., Nicolás, G., López-Climent, M., Gómez-Cadenas, A. and Nicolás, C. (2009) Cross-talk between gibberellins and salicylic acid in early stress responses in Arabidopsis thaliana seeds. Plant Signal Behavior, 4, 750-751. doi:10.4161/psb.4.8.9175 [77] Letunic, I. and Bork, P. (2007) Interactive tree of life (iTOL): An on line tool for phylogenetic tree display and annotation. Bioinformatics, 23, 127-128. doi:10.1093/bioinformatics/btl529 ABBREVIATIONS ARM: armadillo PUB: plant U-box ARC1: armadillo-repeats containing protein-1 SRK: S-locus receptor kinase SLG: S-locus glycoprotein
|