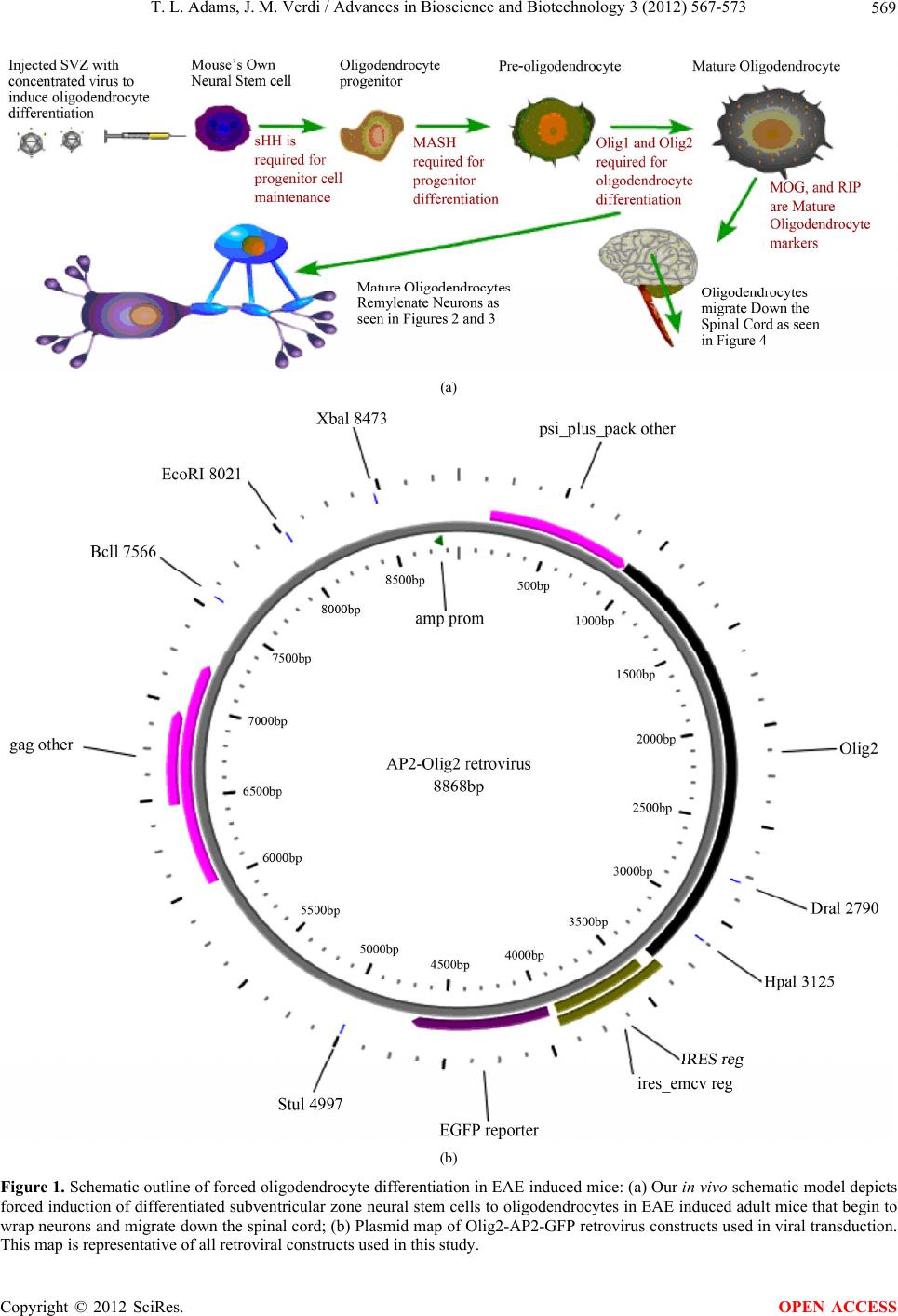
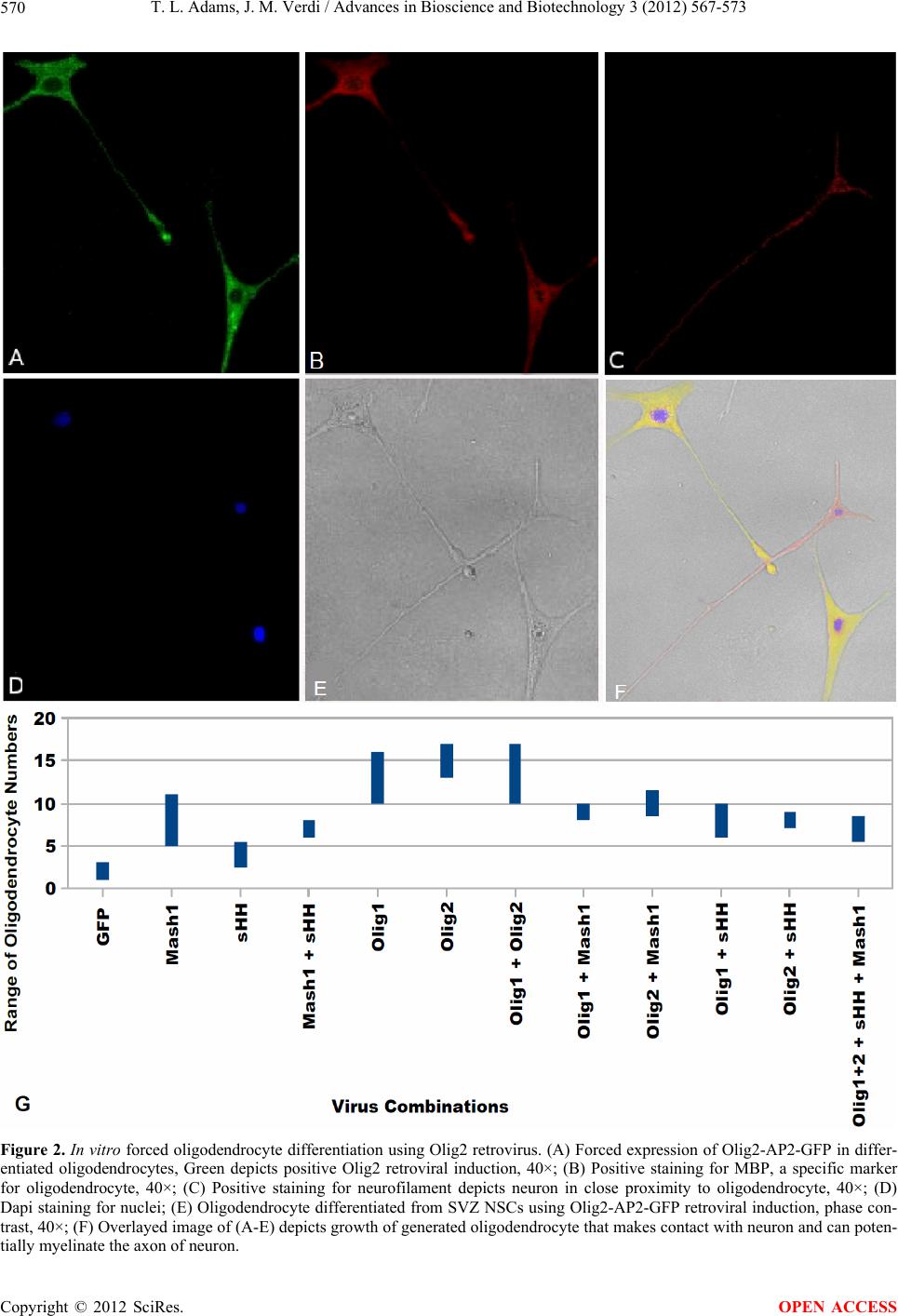
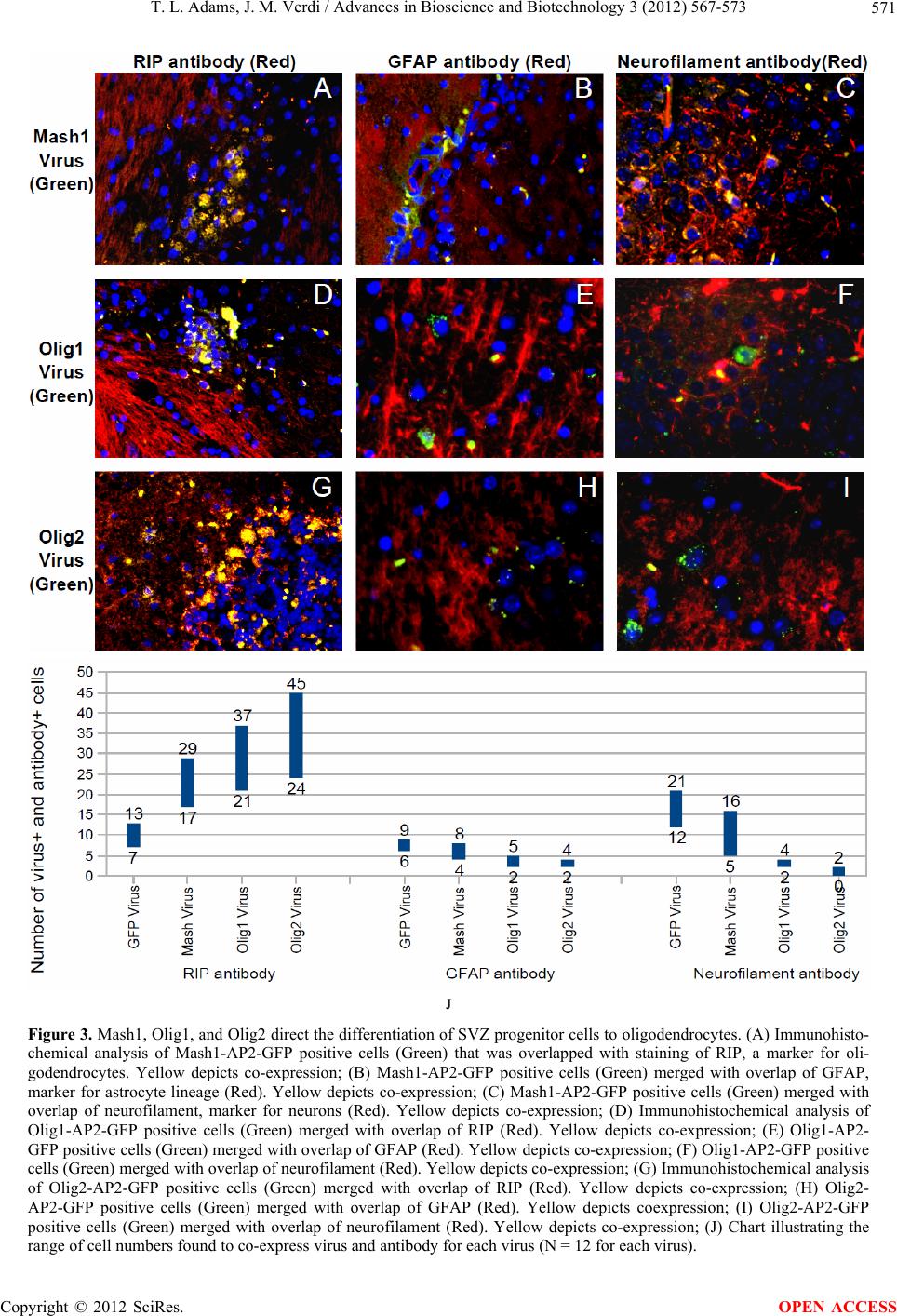
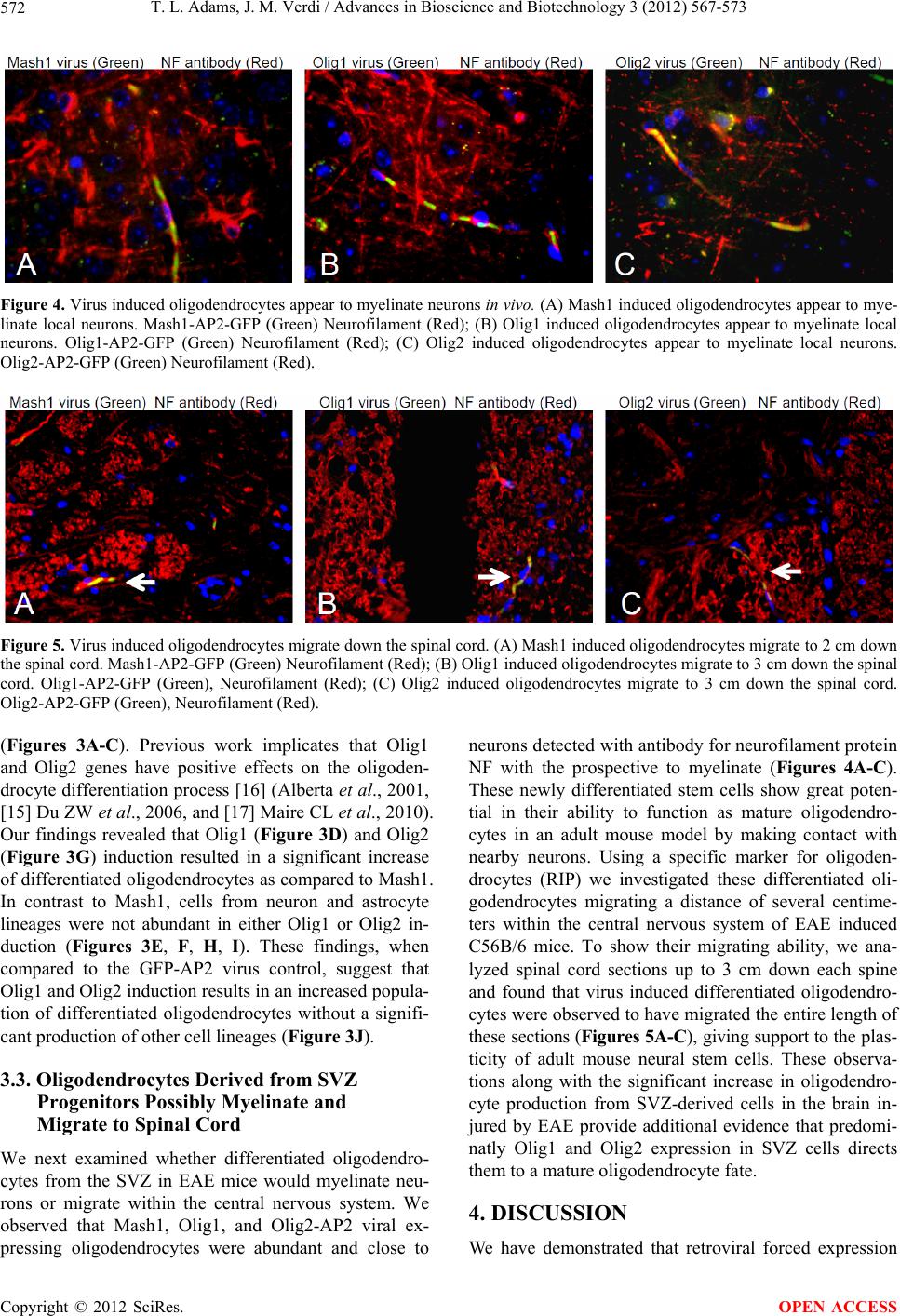
T. L. Adams, J. M. Verdi / Advances in Bioscience and Biotechnology 3 (2012) 567-573 573
of Mash1, Olig1, and Olig2 genes, involved in enhanced
oligodendrogenesis, can contribute to directing the dif-
ferentiation of adult subventricular zone neural stem cells
to functional oligodendrocytes. Mash1 alone or with Shh
produced oligodendrocytes, but it was Olig1 and Olig2
that gave us a much larger increase in oligodendrocyte
numbers while creating fewer cells from the neuronal
lineage. These oligodendrocytes are also capable of mi-
gration and possible myelination. These findings could
allow us to design small molecules that would be used to
force ones own neural stem cells to make oligodendro-
cytes and remyelinate the damaged axons and conse-
quently restore neuronal function.
These reports show that targeting oligodendrocyte
production and possible remylination is a viable thera-
peutic strategy for restoration of neuronal function. We
envision that results of the proposed study may extend to
several neurological disorders where cellular replace-
ment is needed. Future work should be done to find
whether all the oligodendrocytes formed are mature
enough to partake in full remyelination of axons that
have loss of function due to EAE.
5. ACKNOWLEDGEMENTS
The authors would like to thank Dr. Jennifer Rochira and Dr. Aldona
Karaczyn for their knowledge and insight for experimental design.
Thank you to Dr. Josephine Nalbantoglu for sharing the AP2 retrovirus
construct. T.A. carried out experiments, analyzed data, and wrote the
manuscript. J.M.V. conceived the experimental design. Authors have
no conflict of interest.
REFERENCES
[1] Raff, M., Apperly, J., Kondo, T., Tokumoto, Y. and Tang,
D. (2001) Timing cell-cycle exit and differentiation in
oligodendrocyte development. Novartis Found Symposia,
237, 100-107; discussion 107-112, 158-163.
[2] Raff, M.C., Durand, B. and Gao, F.B. (1998) Cell number
control and timing in animal development: The oligoden-
drocyte cell lineage. International Journal of Develop-
ment Biology, 42, 263-267.
[3] Stemple, D.L. and Anderson, D.J. (1992) Isolation of a
stem cell for neurons and glia from the mammalian neural
crest. Cell, 71, 973-985.
doi:10.1016/0092-8674(92)90393-Q
[4] Galipeau, J., Andre, H.L., Paquin, A., Sicilia, F., Karpati,
G. and Nalbantoglu, J. (1999) Vasicular stomatitis virus g
pseudotyped retrovector mediates effective in vivo sui-
cide gene delivery in experimental brain cancer. Cancer
Research, 59, 2384-2394.
[5] Olsson, T. (1995) Critical influences of the cytokine or-
chestration on the outcome of myelin antigenspecific
T-cell autoimmunity in experimental autoimmune enceph-
alomyelitis and multiple sclerosis. Immunological Re-
views, 144, 245-268.
doi:10.1111/j.1600-065X.1995.tb00072.x
[6] Matluk, N., Rochira, J., Karaczyn, A., Adams, T. and
Verdi, J.M. (2010) A role for NRAGE in NF-κB activa-
tion through the non-canonical BMP pathway. BMC Bi-
ology, 8, 7. doi:10.1186/1741-7007-8-7
[7] Parras, C.M., Hunt, C., Sugimori, M., Nakafuku, M.,
Rowitch, D. and Guillemot, F. (2007) The proneural gene
Mash1 specifies an early population of telencephalic oli-
godendrocytes. The Journal of Neuroscience, 27, 4233-
4242. doi:10.1523/JNEUROSCI.0126-07.2007
[8] Johnson, J. E., Birren, S. J., Saito, T. and Anderson, D. J.
(1992) DNA binding and transcriptional regulatory activ-
ity of mammalian achaete-scute homologous (MASH)
proteins revealed by interaction with a muscle-specific
enhancer. Proceedings of the National Academy of Sci-
ences, 89, 3596-3600. doi:10.1073/pnas.89.8.3596
[9] Rogister, B., Ben-Hur, T. and Dubois-Dalcq, M. (1999)
From neural stem cells to myelinating oligodendrocytes.
Molecular and Cellular Neuroscience, 14, 287-300.
doi:10.1006/mcne.1999.0790
[10] Alberta, J.A., Park, S.K., Mora, J., Yuk, D., Pawlitzky, I.,
Iannarelli, P., Vartanian, T., Stiles, C.D. and Rowitch,
D.H. (2001) Sonic hedgehog is required during an early
phase of oligodendrocyte development in mammalian
brain. Molecular and Cellular Neuroscience, 18, 434-441.
doi:10.1006/mcne.2001.1026
[11] Rogister, B., Ben-Hur, T. and Dubois-Dalcq, M. (1999)
From neural stem cells to myelinating oligodendrocytes.
Molecular and Cellular Neuroscience, 14, 287-300.
doi:10.1006/mcne.1999.0790
[12] Othman, A., Frim, D.M., Polak, P., Vujicic, S., Arnason,
B.G. and Boullerne, A.I. (2011) Olig1 is expressed in hu-
man oligodendrocytes during maturation and regeneration.
Glia, 59, 914-926. doi:10.1002/glia.21163
[13] Du, Z.W., Li, X.J., Ngyen, G.D. and Zhang, S.C. (2006)
Induced expression of Olig2 is sufficient for oligoden-
drocyte specification but not for motoneuron specifica-
tion and astrocyte repression. Molecular and Cellular
Neuroscience, 33, 371-380.
doi:10.1016/j.mcn.2006.08.007
[14] Jessberger, S. and Gage, F.H. (2009) Fate plasticity of
adult hippocampal progenitors: Biological relevance and
therapeutic use. Trends in Pharmacological Sciences, 30,
61-65. doi:10.1016/j.tips.2008.11.003
[15] Lubetzki, C., Demerens, C., Anglade, P., Villarroya, A.,
Frankfurter, A., Lee, M.Y. and Zalc, B. (1993) Even in
culture, oligodendrocytes meyelinate solely axons. Pro-
ceedings of the National Academy Sciences, 40, 6820-
6824.
[16] Zhu, G., Mehler, M.F., Zhao, J., Yu Yung, S. and Kessler,
J.A. (1999) Sonic hedgehog and BMP2 exert opposing
actions on proliferation and differentiation of embryonic
neural progenitor cells. Developmental Biology, 215, 118-
129. doi:10.1006/dbio.1999.9431
[17] Maire, C.L., Wegener, A., Kerinon, C. and Nait Oumes-
mar, B. (2010) Gain-of-function of Olig transcription fac-
tors enhances oligodendrogenesis and myelination. Stem
Cells, 28, 1611-1622. doi:10.1002/stem.480
Copyright © 2012 SciRes. OPEN ACCESS