Human Embryo Neuronal Culture in Vitro: A Model to Study Cellular Physiology, Receptors, Power and
Toxicity of Cytostatic Drugs for Human Use 325
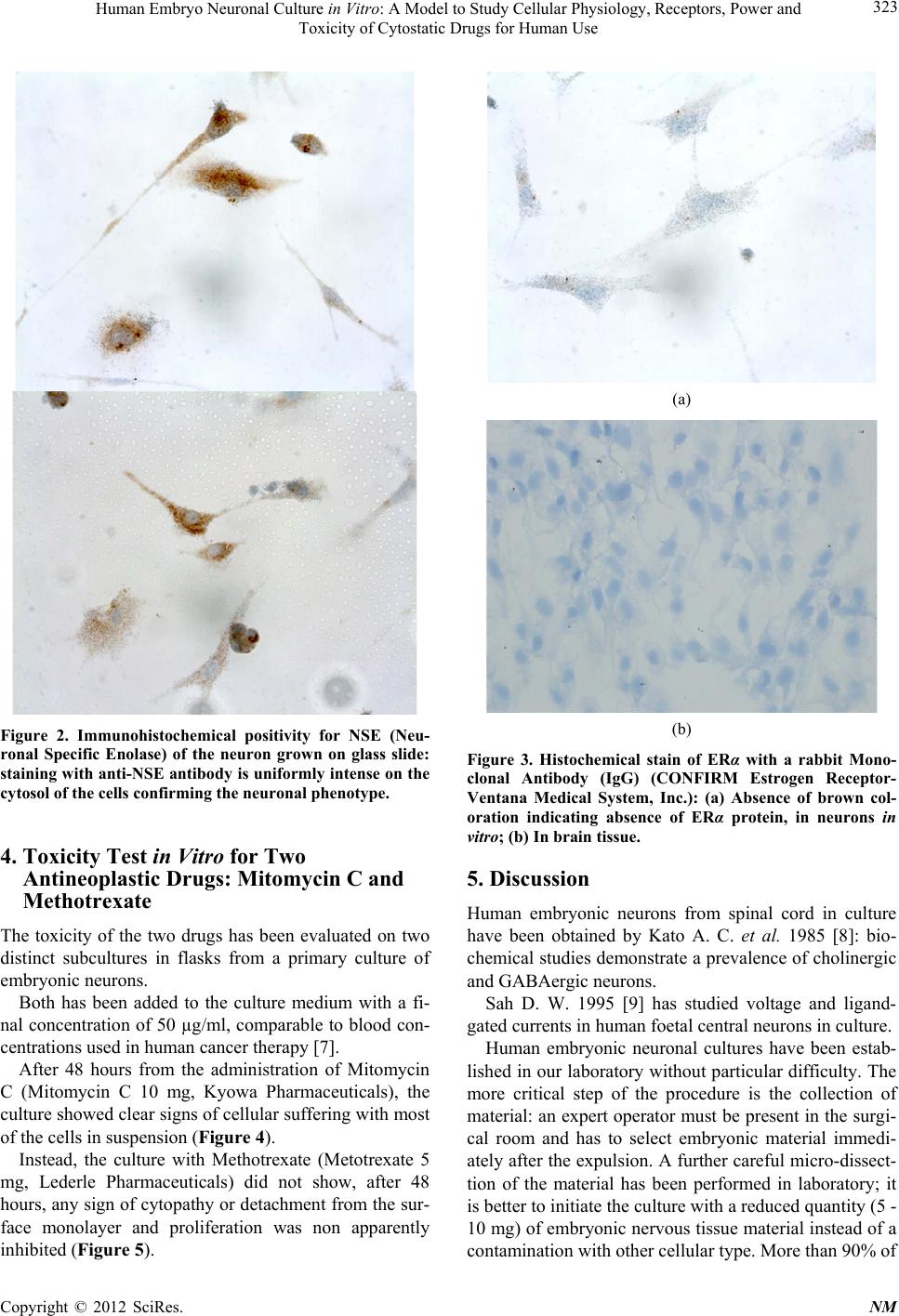
The cytogenetic investigation has easily been per-
formed on our human embryonic neurons (Figure 6); the
analysis of chromosomal fragility and SCE (Sister
Chromatide Exchange) is another powerful method to
test the clastogenic activity of many substances for hu-
man therapy [21,22].
REFERENCES
[1] H. W. Li, H. Liu, C. E. Corrales, J. R. Risner, J. Forrester,
J. R. Holt, S. Heller and A. S. B. Edge, “Differentiation of
neurons from Neural Precursors Generated in Floating
Spheres from Embryonic Stem Cells,” BMC Neurosci-
ence, Vol. 10, 2009, p. 122.
doi:10.1186/1471-2202-10-122
[2] M. Schuldiner, O. Yanuka, J. Itskovitz-Eldor, D. A. Mel-
ton and N. Benvenisty, “Effects of Eight Growth Factors
on the Differentiation of Cells Derived from Human Em-
bryonic Stem Cells,” Proceedings of the National Acad-
emy of Sciences of USA, Vol. 97, No. 21, 2000, pp. 11307-
11312. doi:10.1073/pnas.97.21.11307
[3] U. S. Sohur, J. G. Emsley, B. D. Mitchell and J. D. Mack-
lis, “Adult Neurogenesis and Cellular Brain Repair with
Neural Progenitors, Precursors and Stem Cells,” Phi-
losophical Transactions of the Royal Society B: Biologi-
cal Sciences, Vol. 361, No. 1473, 2006, pp. 1477-1497.
doi:10.1098/rstb.2006.1887
[4] C. E. Corrales, L. Pan, H. W. Li, M. C. Liberman, S.
Heller and A. S. B. Edge, “Engraftment and Differentia-
tion of Embryonic Stem Cell-Derived Neural Progenitor
Cells in the Cochlear Nerve Trunk: Growth of Processes
into the Organ of Corti,” Journal of Neurobiology, Vol.
66, No. 13, 2006, pp. 1489-1500. doi:10.1002/neu.20310
[5] M. Emgard, L. Holmberg, E. B. Samuelsson, B. A. Bahr,
S. Falci, A. Seiger and E. Sundström, “Human Neural
Precursor Cells Continue to Proliferate and Exhibit Low
Cell Death after Transplantation to the Injured Rat Spinal
Cord,” Brain Research, Vol. 1278, 2009, pp. 15-26.
doi:10.1016/j.brainres.2009.04.012
[6] Y.-K. Chang, M.-H. Chen, Y.-H. Chiang, Y.-F. Chen,
W.-H. Ma, C.-Y. Tseng, B.-W. Soong, J. H. Ho and O. K.
Lee, “Mesenchymal Stem Cell Transplantation Amelio-
ratesmotor Function Deterioration of Spinocerebellara-
taxia by Rescuing Cerebellar Purkinje Cells,” Journal of
Biomedical Science, Vol. 18, 2011, p. 54.
doi:10.1186/1423-0127-18-54
[7] E. Chu and A. Sartorelli, “Antineoplastic Chemotherapy,”
In: Katzung Basic & Clinical Pharmacology, 9th Edition,
The McGraw-Hill Companies, Inc., 2004, pp. 898-932.
[8] A. C. Kato, G. Touzeau, D. Bertrand and C. R. Bader,
“Human Spinal Cord Neurons in Dissociated Monolayer
Cultures: Morphological, Biochemical, and Electro- phy-
siological Properties,” The Journal of Neuroscience Co-
pyright Society for Neuroscience, Vol. 5, No. 10, 1985,
pp. 2750-2761.
[9] D. W. Sah, “Human Fetal Central Neurons in Culture:
Voltage- and Ligand-Gated Currents,” Journal of Neuro-
physiology, Vol. 74, No. 5, 1995, pp. 1889-1899.
[10] L. Q. Zhao and R. D. Brinton, “Select Estrogens within
the Complex Formulation of Conjugated Equine Estro-
gens (Premarin®) Are Protective against Neurodegenera-
tive Insults: Implications for a Composition of Estrogen
Therapy to Promote Neuronal Function and Prevent Alz-
heimer’s Disease,” BMC Neuroscience, Vol. 7, 2006, p.
24. doi:10.1186/1471-2202-7-24
[11] G. E. Gillies and S. McArthur, “Estrogen Actions in the
Brain and the Basis for Differential Action in Men and
Women: A Case for Sex-Specific Medicines,” Pharma-
cological Reviews, Vol. 62, No. 2, 2010, pp. 155-198.
doi:10.1124/pr.109.002071
[12] J. Matthews and J. A. Gustafsson, “Estrogen Signaling: A
Subtle Balance between ER Alpha and ER Beta,” Mo-
lecular Interventions, Vol. 3, No. 5, 2003, pp. 281-292.
[13] Z. Sheng, J. Kawano, A. Yanai, R. Fujinaga, M. Tanaka,
Y. Watanabe and K. Shinoda, “Expression of Estrogen
Receptors (Alpha, Beta) and Androgen Receptor in Sero-
tonin Neurons of the Rat and Mouse Dorsal Raphe Nuclei;
Sex and Species Differences,” Neuroscience Research,
Vol. 49, No. 2, 2004, pp. 185-196.
[14] R. J. Handa, S. Ogawa, J. M. Wang and A. E. Herbison,
“Roles for Estrogen Receptor Beta in Adult Brain Func-
tion,” Journal of Neurobiology, Vol. 24, 2011, pp. 160-
173.
[15] B. Greco, E. A. Allegretto, M. J. Tetel and J. D. Blaustein,
“Coexpression of ER_ with ER and Progestin Receptor
Proteins in the Female Rat Forebrain: Effects of Estradiol
Treatment,” Endocrinology, Vol. 142, 2001, pp. 5172-
5181. doi:10.1210/en.142.12.5172
[16] J. E. Donahue, E. G. Stopa, R. L. Chorsky, J. C. King, H.
M. Schipper, S. A. Tobet, J. D. Blaustein and S. Reichlin,
“Cells Containing Immunoreactive Estrogen Receptor-
Alpha in the Human Basal Forebrain,” Brain Researvh,
Vol. 856, No. 1-2, 2000, pp. 142-151.
doi:10.1016/S0006-8993(99)02413-0
[17] D. Montague, C. S. Weickert, E. Tomaskovic-Crook, D.
A. Rothmond, J. E. Kleinman and D. R. Rubinow, “Oes-
trogen Receptor Alpha Localisation in the Prefrontal
Cortex of Three Mammalian Species,”Journal of Neuro-
endocrinology, Vol. 20, No. 7, 2008, pp. 893-903.
doi:10.1111/j.1365-2826.2008.01743.x
[18] T. A. Ishunina, F. P. Kruijver, R. Balesar and D. F.
Swaab, “Differential Expression of Estrogen Receptor
Alpha and Beta Immunoreactivity in the Human Supra-
optic Nucleus in Relation to Sex and Aging,” Journal of
Clinical Endocrinology and Metabolism, Vol. 85, No. 9,
2000, pp. 3283-3291. doi:10.1210/jc.85.9.3283
[19] F. F. N. Rizzzuto, “Estrogen Receptors Localization in
the Spinal Trigeminal Nucleus: An Immunohistochemical
Study in Humans,” European Journal of Pain, Vol. 15,
No. 10, 2011, pp. 1002-1007.
[20] M. J. Gallego, P. Porayette, M. M. Kaltcheva, R. L. Bo-
wen, S. V. Meethal and C. S. Atwood, “The Pregnancy
Hormones Human Chorionic Gonadotropin and Proges-
terone Induce Human Embryonic Stem Cell Proliferation
and Differentiation into Neuroectodermal Rosettes,” Stem
Copyright © 2012 SciRes. NM