 Open Journal of Genetics, 2012, 2, 136-154 OJGen http://dx.doi.org/10.4236/ojgen.2012.23019 Published Online September 2012 (http://www.SciRP.org/journal/ojgen/) Analysis of candidat e g enes of QTL and chromosomal regions for essential hypertension in the rat model Lishi Wang1,2, Jiaqian Zhu1,3, Yue Huang1, Qing Xiong4, Cong-Yi Wang5, Arnold Postlethwaite6, Yongjun Wang7, Weikuan Gu1* 1Departments of Orthopaedic Surgery-Campbell Clinic and Pathology, University of Tennessee Health Science Center (UTHSC), Memphis, USA 2Department of Basic Medicine, Inner Mongolia Medical University, Huhhot City, China 3Rust College, Holly Spring, USA 4Department of Biostatistics and Bioinformatics, Duke University, Durham, USA 5Center for Biotechnology and Genomic Medicine, Medical College of Georgia, Augusta, USA 6Department of Medicine, University of Tennessee Health Science Center (UTHSC), Memphis, USA 7Department of Neurology, Beijing Tiantan Hospital, Capital University of Medical Sciences, Beijing, China Email: Lwang37@uthsc.edu, Jzhu@rustcollege.edu, hyue@uthsc.edu, qxiong1@uthsc.edu, cwang@mail.mcg.edu, apostlet@uthsc.edu, yongjunwang111@yahoo.com.cn, *wgu@uthsc.edu Received 20 May 2012; revised 25 June 2012; accepted 10 July 2012 ABSTRACT This is an in silico analysis of quantitative trait loci (QTLs), genes, polymorphisms, and chromosomal regions regulating hypertension in the rat genome. Utilizing PGmapper, a program that matches phenol- types to genes, we identified 266 essential hyperten- sion-associated genes (HyperA), and 83 of these genes contain known hypertension-associated polymorphi- sms (HyperAP). The majority of HyperAP have been reported in recent years. Surprisingly, only a few of these HyperAP genes have been investigated for their candidacy as the QTL for hypertension. The fre- quency of candidate genes within peak regions of the QTL is higher than the rest of the QTL region. We also found that QTL located in both gene-rich regions and gene-rich chromosomes contained the most can- didate genes. However, the number of candidate genes within a peak region is not associated with the number of total genes in a QTL region. This data could not only facilitate a more rapid and compre- hensive identification for the causal genes underlying hypertension in rats, but also provides new insights into genomic structure in regulation of hypertension. Keywords: Chromosome; Hypertension; QTL; Gene; Polymorphism; Rat 1. INTRODUCTION Blood pressure and hypertension in humans are quantita- tive traits controlled by many genes [1-3]. Because of the difficulties in studying genetic variations relating to hy- pertension in humans, researchers chose the rat as an ideal model. In recently review articles, progress for the genetic dissection of essential hypertension in humans using a rat model have been summarized. Cowley [5] pointed out that the next daunting task is gene identifica- tion and validation. Currently, more than 300 Quantitative trait loci (QTLs) for blood pressure and hypertension are reported in the rat genome (see the Rat Genome Database web site at: http://rgd.mcw.edu/), and these QTLs are found in every rat chromosome [6]. More than 200 candidate genes relating to hypertension have been identified in rats and some of them, such as Kcnj1 and Drd2, are positioned within a QTL [7,8]. Most of these identified genes are relatable to human chromosomes. However, further ana- lysis on the relationship between these candidate genes and their likelihood for being genes involved in the QTLs for hypertension will enhance the identification of the real causal genes responsible for QTLs. In addition to just simply further analysis, more ques- tions need to be answered in the investigation for the candidate genes responsible for the hypertension QTLs. There are three main questions that need answering: Do these candidate genes include all the genes affecting blood pressure and hypertension? How many candidate genes are within the hypertension QTLs and how close are these genes to peak markers? What is a candidate gene’s relative potential candidacy for being a gene truly responsible for the QTL? The third question is the most important of the three and needs a definitive answer. Currently, due to the combination of rapidly developed genome sequence information and online literature, it is *Corresponding author. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 137 now possible to pinpoint accurately genes within a QTL and evaluate them much more efficiently. 2. MATERIALS AND METHODS 2.1. Software In this investigation, we used our recently developed software named PGMapper (http://www.genediscovery.org/pgmapper/index.jsp) to systematically examine the candidate genes for the hy- pertension QTLs across the whole rat genome [9]. PGMapper identified all the possible candidate genes for the hypertension QTL by combining the mapping infor- mation from Ensemble database, updated literature in- formation from PubMed, and the Online Medelian In- heritance in Man (OMIM) database. 2.2. Collection of QTL Loci We first selected blood pressure as a trait to pick up all of the possible hypertension QTLs from the Rat Genome Database (RGD, http://rgd.mcw.edu/). We then selected all the QTL that have a logarithm of odds (LOD) score > 2.8. This selection criterion was based on accepted link- age criteria. A LOD score of >4.3 was considered sig- nificant while a LOD score between 2.8 and 4.3 was considered suggestive for linkage [10,11]. If two QTL were overlapped and connected, we analyzed them inde- pendently. However, if one QTL is located within an- other QTL, we just used the QTL with the larger genome size. 2.3. Examination of Candidate Genes We used flanking markers to search candidate genes for QTLs that are fine-mapped and well-defined. If a flank- ing marker has no sequence location on Ensemble data- base, we used either nearby markers or markers at the peak region of the QTL (according to the curve of the LOD score). If a marker in the peak region of the QTL was used, we searched candidate genes using genomic regions of 20,000,000 base pairs (bp) on each side of the peak marker. After we searched for candidate genes using PGMap- per, visual examination of these selected candidate genes was conducted to determine the potential of each indi- vidual gene involved. First, the genes full names were checked to ensure their abbreviations. Next, at least one abstract per gene from the PGMapper’s report detailing each selected gene was read to determine the candidacy of that gene. In the majority of cases, more than one ab- stract was read to confirm the importance of that gene. In order to discern whether any of these candidate genes had been investigated for their candidacy in the hypertension QTL, we conducted a separate literature search on PubMed using the key words “QTL + hyper- tension + ‘gene name’.” We then read the associated literature from this search to determine any connection between our preliminary candidate genes from PGMap- per and essential hypertension. A gene is considered an essential hypertension-associated gene if it is associated with essential hypertension in at least one of the follow- ing studies: 1) functional studies (i.e., knockouts, trans- genics, mutagenesis, RNA interference, etc.); 2) associa- tion studies; 3) clinical studies. In case any genes associated to essential hypertension by association studies resulted in an inconclusive result, we excluded these genes from our candidate gene list. Candidate genes were examined whether they have been studied in the human population previously and their polymorphisms have been linked with hypertension. The key words used in our search were “hypertension and polymorphism”. Thus, only when both hypertension and polymorphism appear in the same abstracts with a gene name will PGMapper collect it into our list of genes needing further review. 2.4. Analysis of Gene Chromosomal Position and Their Candidacy We also searched for candidate genes within 1.5 cM re- gions from the peak marker in a QTL by PGMapper. These results indicated the importance of each selected candidate genes in the QTL. Total genomic bps within a QTL region were deter- mined by subtracting the nucleotide base position of the right flanking marker from the position of left flanking marker. Bps per gene of a QTL region were calculated by dividing the total bps of a QTL region by the total genes according to data from Ensembl (bps per gene = total bps/total genes). Bps per candidate gene of a QTL region were calculated by dividing the total bps of a QTL region by the number of candidate genes determined from our aforementioned methods (bps per candidate gene = total bps/total candidate genes). Gene-rich and gene-poor regions on chromosomes were determined by comparison of the bps per gene in a QTL region or chromosome. P values were determined using student’s t test. R values of correlation analysis were obtained with Excel. 3. RESULTS 3.1. QTL and Their Candidate Genes The 62 hypertension QTLs cover 2,015,062,129 bp ge- nomic sequences, which is roughly 73% of the total rat genome. Every autosomal and the X chromosome, except chromosomes 19 and Y (Table 1), contain at least one Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 Copyright © 2012 SciRes. 138 OPEN ACCESS Table 1. QTL and candidate genes identified from rat model. Chromosomes QTLs Total genes Length (bp) Hypertension candidate genesEssential hypertension associated genes Hypertension- associated polymorphisms Candidates within 1.5 cM X 2 454 48,888,222 5 4 0 1 1 8 3419 247,322,280 75 63 22 1 2 5 1852 256,217,383 44 12 4 1 3 4 1706 178,038,027 25 16 2 3 4 4 1006 129,644,537 22 16 3 3 5 3 1562 151,887,861 20 17 7 0 6 2 1087 121,776,584 18 14 8 0 7 4 1037 109,297,243 22 16 2 2 8 3 1111 12,479,341 15 13 1 0 9 2 568 71,306,988 13 10 2 1 10 3 1502 89,111,128 26 22 6 0 11 2 569 71,856,931 5 3 1 0 12 2 714 72,770,456 12 11 3 1 13 4 745 87,298,215 15 15 10 0 14 2 459 56,035,440 9 7 5 1 15 3 192 54,324,114 1 0 0 1 16 1 400 41,833,193 6 6 1 2 17 3 757 87,545,639 6 6 1 1 18 3 657 81,554,492 10 10 3 1 20 2 469 45,874,055 6 5 2 2 Total 62 20,266 2,015,062,129 355 266 83 21 hypertension QTL. The genomic size of these QTL ranges from 5,873,857 bps to 167,769,667 bps. Within the 2,015,062,129 bp genomic sequences, a total of 20,266 genes have been located. The number of genes within a QTL region ranges from 79 to 1398. The aver- age gene density throughout the whole rat genome (ex- cluding ChrY) is about one gene per 99,918 bp. Within the total 2,015,062,129 bp genomic sequences represent- ing the hypertension QTL, there are roughly one gene per 99,430 bp, which is similar to the average gene per bp ratio found in the rat genome. From the total 20,266 genes related to hypertension, 266 were selected as can- didate genes for essential hypertension (Table 1). Basi- cally, about one candidate gene was chosen per 76 se- lected genes. These 266 candidate genes are located throughout all autosomal and X chromosomes excluding chromosome 15 (which contains no obvious recognizable candidate genes), chromosome 19, and chromosome Y. The number of candidate genes within a QTL region can be as low as zero to as high as 32. The number of QTL on each individual chromosome varies from one to five. There is a positive correlationship between the number of QTL on a chromosome to the number of can- didate genes on a chromosome (R = 0.787). 3.2. Essential Hypertension Associated (HyperA) Candidate Genes and Essential Hypertension Associated Gene Polymorphisms (HyperAP) While the majority of candidate genes have been reported or listed in rat genome databases, we identified many new candidate genes during our search. Particularly, we have identified 83 polymorphic genes with publicized linkage to human hypertension (HyperAP). While HyperA genes are found on every chromosome containing one or more QTL regions, HyperAP genes are found only on auto- somal chromosomes containing QTL regions. In general, the number of HyperA genes is positively correlated to the number of HyperAP genes (Figure 1). Chromosome 1 has the largest number of HyperA genes and, consequentially, the largest number of HyperAP genes too. For instance, Chromosome X has a small number of HyperA genes and, therefore, does not have any HyperAP genes. Surprisingly, in spite of demonstrated linkage between the polymorphic HyperAP genes and hypertension, the candidacy for the majority of these genes for hyperten- sion QTL has not been investigated. Examples of the few  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 139 HyperAP genes investigated, with positive or negative results, are monocyte chemotactic protein-1 (CCL2), nitric oxide synthase 2A (NOS2), and natriuretic peptide precursor B (NPPB). A study identifying candidate genes for the QTL influencing blood pressure within Chr10 reported that the expression levels of CCL2 mRNA showed no difference between the kidneys of Dahl salt-sensitive (DS) and Lewis (LEW) rats fed a normal diet. However, CCL2 mRNA expression levels in DS were 10-fold higher than those in LEW with a high-salt diet [12]. Yet another study reported that NOS2 is not supported as a candidate for the QTL capable of causing a blood pressure difference between the S and MNS rats [13]. Nevertheless, it was suggested that the nitric oxide system appears to be secondarily involved in the regula- tion of blood pressure in the S rat, as evidenced by physiological data. In a study using a congenic approach with a rat model, the relationship between natriuretic peptide precursor A (NPPA) and NPPB genes and hy- pertension was examined, but an association between the NPPB gene and blood pressure was not found [14]. 3.3. Candidate Genes for the QTL on Gene- Rich and/or Gene-Poor Chromosomes We found that gene density for QTL on different chro- mosomes varies significantly. QTLs on two chromo- somes are located in gene-rich regions. These two chro- mosomes are chromosome 1 (one gene per 72,337 bp) and chromosome 10 (one gene per 59,328 bp). However, QTLs on chromosome 15 reside in extremely gene-poor regions (one gene per 282,938 bp). The other QTLs in gene-poor regions are on chromosome 11 (one gene per 30,000 bp) (Figure 2). 1 0 10 20 30 40 50 60 70 80 90 100 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 20, X Ch romosome Number of Gene s HyperA HyperAP Figure 1. Number of essential hypertension associated genes (HyperA) and genes of polymorphic associated to essential hypertension in human population (HyperAP) in QTL regions on chromosomes. 0 2000000 4000000 6000000 8000000 10000000 12000000 14000000 16000000 Chr1 Chr10 Chr11 Chr15 Bps per Candidate Figure 2. Number of candidate genes in QTL of gene rich chromosomes and gene poor chromosomes. Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 140 3.3.1. Chromosome 1 There are 6 QTLs that overlap and cover a large propor- tion of this chromosome. These QTL are Bp95, Bp96, Bp173, Bp196, Bp 255, and Bp259 (Supplementary Table 1). These QTL extend from 90,448,902 to 247,322,280 with a length of 156,873,379 bps. All of these QTL are located in gene-dense regions. Gene den- sities in four of these loci are relatively high. The first QTL, Bp95, is in the proximal region of this chromosome, and is located in the interval from 0 bp to 22,869,052 bp. This region contains 248 genes and four candidate genes (Table 1). Among them, transforming growth factor beta 1 (TGF-beta 1) is a fibrogenic cyto- kine implicated in hypertension in African-Americans. Suthanthiran et al. [15] demonstrated that African- Americans with end-stage renal disease (ESRD) have higher circulating levels of TGF-beta 1 protein compared to Caucasians with ESRD. They have also found that hyperexpression of TGF-beta 1 is more frequent in Afri- can-Americans with hypertension than in Caucasians. Accordingly, they proposed that TGF-beta 1 hyperex- pression may be an important mediator of hypertension and hypertensive nephrosclerosis. More recently, and importantly, polymorphisms in TGF-beta 1 have been associated with cardiovascular and renal damage. This data strongly suggests that TGF-beta 1 is the causal gene for this QTL. The other noteworthy gene in this QTL is osteoglycin (OGN). In a quantitative trait transcript (QTT) analysis study of the cardiac transcriptome in the rat, Petretto et al. [16] showed that OGN is a key regula- tor of left ventricular mass (LVM) in rats, mice and hu- mans, and suggested that OGN modifies the hypertrophic response to extrinsic factors such as hypertension and aortic stenosis. QTL Bp96 overlaps and is connected to Bp95. Bp96 covers a region from 22,842,962 to 102,532,417 bps and contains more than 1000 genes. Twenty genes were re- garded as candidate genes. Among these 20 genes, polymorphisms in at least five of them have been linked to hypertension in human populations. TGF-beta 1 is also among these candidates. The other important gene in this group is Apolipoprotein E (APOE) [17]. Associations between polymorphisms of APOE and hypertension have been extensively studied in several countries and differ- ent populations. However, the results are sometimes controversial and vary from population to population. Kcnj11 is another gene shown to be associated with hy- pertension in humans [18]. Specifically, there is signifi- cant evidence linking polymorphisms of Kcnj11 to blood pressure variations in Korean [18] and Japanese [19] populations. In addition, SNPs in the renal glomeru- lus-specific cell adhesion receptor (Nphs1) have known associations with hypertension in the Japanese popula- tion [20]. Glucocorticoid-regulated kinase (SGK1) also has a demonstrated relationship with hypertension in populations specifically those in Germany [21] and Swi- tzerland [22]. The Organic Cation Transporter 2, OCT2 (SLC22A2), implicated in both renal dopamine handling as well as the inactivation of circulating catecholamines, is also thought to be involved in blood pressure regula- tion [23]. Two QTL, Bp173 and Bp196, largely overlap each other and occupy the genome regions of 115,946,375 bp to 189,900,838 bp and 119,780,561 bp to 204,280,279 bp, respectively. From the more than 1200 known genes within these two QTL, a total of 33 candidate genes were found (Supplementary Table 1). Ten of these 33 genes have been linked to hypertension in humans by polymer- phic analysis. Those genes are the following: 1) Neuro- trophic tyrosine kinase, receptor, type 3 (Ntrk3), a gene studied in the Japanese population [20]; 2) Prolylcar- boxypeptidase (Prcp) D allele, which is coupled to chronic hypertension and associated with a significant increased risk of preeclampsia in both African-American and non-African-American women [24]; 3) Sodium chan- nel gamma subunit gene (SCNN1G), multiple poly- morphisms in human epithelial SCNN1G are associated with essential hypertension in several populations [25-27]; 4) Calpain-5 (Capn5), Capn5 variants are associated with diastolic blood pressure and cholesterol levels in the Spanish population [28]; 5) Solute Carrier family 9 mem- ber 2 (SLC9A2), Uromodulin (UMOD), and Elastin (ELN), five polymorphisms in these 3 genes were associ- ated with hypertension status [29]; 6) Uncoupling protein 2 (Ucp2), a common polymorphism of this gene is asso- ciated with hypertension in the Japanese population [30,31]; 7) Inositol polyphosphate phosphatase-like 1 (INPPL1 and SHIP2), variants of these genes affect es- sential hypertension [32]; 8) Purinergic receptor p2y, g protein-coupled, 2 (P2ry2), P2ry2 is among 61 non-syn- onymous polymorphisms of 41 hypertension candidate genes with blood pressure variation in the Japanese popu- lation [19]; 9) SCNN1B, this gene has shown a mod- est-sized but highly significant effect on common genetic variation in SCNN1B on plasma potassium [33]. Interac- tions between the rs889299 SNP and functional SNPs in other genes influencing aldosterone-responsive distal tubular electrolyte transport may be important in the eti- ology of essential hypertension; 10) Spontaneously hy- pertensive rat-clone A-hypertension-associated gene (SAH), SAH has been extensively investigated and its gene vari- ants are associated with obesity-related hypertension in Caucasians [34,35]. Among these 33 candidates, only two genes have been studied for their role as the QTL for hypertension. ApoE was investigated by Yuan et al. [36], and Okuda et al. [37] found that the expression of SAH has been increased in genetic hypertensive rats via microarray analysis. Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 141 The next pair of QTL are Bp255 and Bp259. These two QTL overlap in the region from 195,927,908 bp to 247,322,280 bp. Within these two QTL, 20 candidate genes were identified. However, polymorphisms of only two of these genes (CPT1A and RGS20/GNA14) have been linked to hypertension. Carnitine palmitoyltrans- ferase 1A (CPT1A) is among four genes that have their polymorphisms associated with left ventricular hyper- trophy (LVH) in essential hypertension [38]. In “The Millennium Genome Project for Hypertension”, Kohara et al. [39] found the genes RGS20 and GNA14. Domi- nant models for these minor alleles had significant asso- ciation with hypertension in the Japanese population. 3.3.2. Chromosome 10 There are 3 overlapping QTLs (Bp57, Bp168, and Bp186) that cover a large proportion of this chromosome (Sup- plementary Table 1). All of these QTL are located in gene-dense regions. Bp57 covers the genome region from 21,607,720 to 84,443,858 bps. The genome region contains a total of 1026 genes and has an average of one gene per 61,243 bps. A total of 11 candidate genes were identified from this QTL, which is basically one candidate gene per 93 selected genes. From these 11 candidate genes, the five genes that were of special interest were NOS2, CCL2, 12(s)-HETE and ALOX12, and CIAS1. Studying the ratio of circulating nitric oxide to endo- thelin-1 in patients with both systemic sclerosis (SSc) and pulmonary arterial hypertension (PAH), Kawaguchi et al. linked polymorphisms of the nitric oxide synthase 2 (NOS2) gene to susceptibility of both PAH and SSc in the Japanese population [40]. Chemokine, cc motif, ligand 2 (CCL2) has shown a significant and independent association between the -2518G/A polymorphism of the MCP-1 gene (presence of G allele) and hypertension in the Tunisian population [41]. The arachidonic acid-derived metabolite 12-(S) hy- droxyeicosatetraenoic acid (12(S)-HETE), catalyzed by 12-lipoxygenase (12-LOX, ALOX12), exhibits a variety of biological activities with implications in cardiovascular disease. In a study of 200 patients with essential hyper- tension (aged 56 1 years, mean s.e.m., including 97 males) and 166 matched controls (aged 54 1 years, 91 males), Quintana et al., [42] found that a non-synonymous polymorphism in ALOX12 is associated with both essen- tial hypertension and urinary levels of 12(S)-HETE. An intronic variable number of tandem repeat poly- morphisms in the cold-induced autoinflammatory syn- drome 1 (CIAS1) gene modifies gene expression, and is associated with essential hypertension in the Japanese population [43]. Timasheva et al. [44] found that carriers of the IL12B 1159 *A/*A genotype have a lower risk of stroke during a study to reveal the association of inter- leukin-6, interleukin-12, and interleukin-10 gene poly- morphisms with essential hypertension, and its clinical complications in a Tatar ethnic group from Bashkor- tostan, Russia. Bp168, overlaps Bp57 and covers a region from 27,184,742 bp to 102,587,587 bp and has a total of 1398 genes, which is roughly one gene per 53,936 bps. From these genes, 20 were found to be candidate genes for hy- pertension, and eight of these 20 genes, including NOS2, CCL2, ALOX12, are also found in Bp57. On average, every 3,770,142 bp within Bp168 has one candidate gene. Also, from these 20 genes Angiotensin Converting En- zyme (ACE) is a promising candidate gene for essential hypertension (EH) as it plays a key role in blood pressure regulation. ACE has been extensively studied in a variety of human populations with both positive [45] and nega- tive [46] results. Another promising candidate from Bp168 is Solute Carrier family 4, anion exchanger, member 1 (SLC4A1). SLC4A1 is a polymorphism of one of our candidate genes and has been associated with both blood pressure varia- tion and hypertension [47]. Bp186 is another overlapped QTL on this chromosome. It covers a region from 95,066,219 bp to 110,718,848 bp and contains 263 genes, which is roughly one gene per 59,515 bps. A total of five candidate genes were selected from these 263 genes, including ACE and Gh1, which are both overlapped by genes in Bp168. Both NOS2 and ACE have been disqualified as QTLs in the Dahl salt-sensitive (DSS) and Lewis (LEW) rat comparison [48]. However, we did not find any report on the investigation of candidacy for either ALOX12, CIAS1, or SLC4A1 as the QTL for hypertension. 3.3.3. Chromosome 11 Chromosome 11 includes two QTL, Bp187 and Bp- QTLcluster 10. Bp187 is located in a 41,381,787 bp re- gion. It contains as many as 296 genes with an average of 139,803 bps per gene. However, it contains only two candidate genes, which means there is only one candi- date gene per 20,690,893 bps. No polymorphisms of these two genes, adp-ribosylation factor-like 6 and (ARl6) superoxide dismutase 1 (SOD1), have been linked to human hypertension. BpQTLcluster 10 covers a 45,000,000 bp region and has a total of 406 genes, with one gene per every 110,837 bps. It contains four candidate genes. This means there is a candidate gene from every 101 genes or every 11,250,000 bps has one candidate gene. Among these four genes, SNP variations in calcium-sensing receptor (CASR) has been recently linked to human hypertension [49,50]. However, it has never been investigated as a candidate gene in this QTL. Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 Copyright © 2012 SciRes. 142 3.3.4. Chromosome 15 The three QTL found in Chromosome 15 are Bp191, Bp190, and Bp126. Bp191 is located on the proximal region covering a region of 14,323,976 bps. It contains as many as 118 genes; however, none of them were se- lected as candidate genes by our bioinformatic searching. Since Bp190 is located within Bp126 they are considered the same QTL. Bp126 is found within a region of 2,000,000 bps on each side of the peak marker, Ednrb, and contains as many as 158 genes. However, similar to Bp191, there is no obvious candidate gene for hyperten- sion from these 158 genes. OPEN ACCESS 3.4. Candidate Genes in Different QTL on the Same Chromosome Are Correlated t o the Density of Genes in Each QTL Region In general, the number of candidate genes per QTL is directly correlated to the gene density in of the QTL re- gion. The phenomenon is not restricted to the chromo- somes in this study but holds true throughout every ge- nome. The correlation between the total number of genes and the number of nucleotides in the QTL region is 0.817 (Figure 3(a)). The correlation between the total number of genes and the number of candidate genes in the QTL region is 0.827 (Figure 3(b)). QTL from several typical chromosomes are described as below. 3.4.1. Chromosome 2 Chromosome 2 includes the five QTL Bp115, Bp205, Bp14, Bp175, and Bp203. Bp115 is located on the proximal region covering a large 33,349,984 bp region. It contains as many as 218 genes, with an average of 152,981 bps per gene. However, since it contains only five candidate genes there are an average of 669,966 bps per candidate gene. Of these five candidate genes, CAST, THBS4, and F2R are the most noteworthy. Calpastatin (CAST) is among 16 genes that have shown linkage to human hypertension in the Japanese population. Wessel et al. [51] suggest that the A387P variant of the THBS4 gene may be an important deter- minant for the development of myocardial infarction at any age. Gigante B et al. [52] reported that F2R genetic 0 200 400 600 800 1000 1200 1400 1600 020000000400000006000000080000000100000000 120000000140000000160000000 180000000 Bps of nucl eotides number of ge ne s (a) 0 5 10 15 20 25 30 35 05001000 1500 Gene number C andidate gene number (b) Figure 3. (a) Correlationship between number of base pairs of genome and number of genes in QTL regions; (b) Correlationship between number of genes and number of candidate genes for essential hypertension in QTL regions.  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 143 variants may influence the natural history of coronary heart disease (CHD) in patients at high-risk for cardio- vascular events. Bp205 cover a 16,765,253 bp region and has on aver- age one gene per 153,809 bps. However, it contains no candidate gene. Bp14 is located next to Bp205. It covers 167,769,667 bps and has a total of 1284 genes, with one gene per 130,661 bps. It contains 32 candidate genes or one can- didate gene per 53 selected genes. On average, every 5,242,802 bps has one candidate gene. Several important candidate genes include: NPR, GHR, CRH, GSTM, and ARNT. Allelic variants of natriuretic peptide receptor (NPR) genes are associated with a family history of hy- pertension and cardiovascular phenotypes [53], including Npr1 and Npr3. Another of the candidate genes, growth hormone receptor (GHR), has been linked to hyperten- sion in two independent studies [19,54]. In “The Québec Family Study”, a study of genome-wide linkage analysis for systolic and diastolic blood pressure, Rice et al. [55] found an association between polymorphisms of cortico- trophin releasing hormone gene (CRH) and hypertension. A more recent study suggests that knowledge of glu- tathione s-transferase M1 (GSTM1) variant statuses is a potentially useful method for predicting a possible hy- pertensive status after 80 years of age in the Italian popu- lation [56]. Aryl hydrocarbon receptor nuclear transloca- tor-like (ARNT) is associated with susceptibility to hy- pertension in a genetic association study designed to test the relevance of these findings in 1304 individuals from 424 families [57]. Glutathione s-transferase M3 (GSTM3) -63A/C polymorphisms were associated with essential hypertension in Chinese Han populations. It is thought that the C allele might be a risk factor for EH in the Chi- nese Han ethnic group [58]. In a study by Delles et al., [59], it was found that a single nucleotide polymorphism in the 3' region of GSTM5 (rs11807) was associated with hypertension (P = 0.01), and with the T-allele being over-transmitted to hypertensive offspring. The one exception that gene density is not associated with the number of candidate genes on chromosome 2 is the Bp203 QTL, which is overlapped with Bp14, that covers a 30,053,784 bp region. It contains as many as 267 genes, which is an average of 112,560 bps per gene. It has nine candidate genes, which is one candidate gene per 3,339,309 bps. An interesting fact is that out of these nine candidate genes six of them are also found in Bp14. Bp175 covers a 33,278,084 bp region. It contains as many as 220 genes (151,264 bp per gene). It contains 4 candidate genes, which is one candidate gene per 8,319,521 bps. However, none of these four genes have been linked to hypertension in humans. 3.4.2. Chromosome 4 Chromosome 4 includes the four QTL Bp179, Bp86, Bp124, and Bp209, located proximal to distal, respect- tively. Bp179 locates on the proximal region covers a 40,000,132 bp region. It contains as many as 239 genes, with an average of 167,364 bps per gene. However, it contains only 10 candidate genes, which is an average of 4,000,013 bps per candidate gene. Key candidate genes from this QTL include HGF and LEP. Hepatocyte growth factor (HGF) is a growth factor that contributes to protection and/or repair of vascular endothelial cells. Two studies showed that C/A polymorphisms in intron 13 of the HGF gene are associated with susceptibility to essential hypertension in Japanese female subjects [60, 61]. Leptin (LEP) has been widely investigated for its role in hypertension. A recent study indicates that com- mon LEP polymorphisms are associated with blood pressure in the Brazilian Tunisian population [62]. Bp86 has a little overlap with Bp179 and covers a 35,960,924 bp region. It contains total of 1284 genes, with one gene per 92,207 bps. It has 11 candidate genes, or one candidate gene per 35 selected genes. On average, every 3,269,174 bp has at least one candidate gene. The most probable candidate gene from Bp86 is Lep, which is overlapped from Bp179, since none of other genes are linked to human hypertension. Bp124 is largely overlapped with Bp86 and covers a 40,000,190 bp region. It contains as many as 456 genes, and has an average of 87,719 bps per gene. It has 10 candidate genes, which is one candidate gene per 4,000,019 bps. Eight of these 10 candidate genes are overlapped with Bp86. The other two genes have no known association with hypertension. Bp209 covers a 40,000,115 bp region. It contains as many as 251 genes, with an average of 159,363 bps per gene. It has only three candidate genes, or one candidate gene per 13,333,371 bps. Among those three genes, only the CC genotype in oxidized LDL receptor gene (OLR-1) is an independent risk factor for hypertension in the Chinese population [63]. 3.4.3. Chromosome 8 On chromosome 8 the three QTL, are Bp184, Bp262, and Bp263. Bp262 is located in a 75,560,673 bp region close to proximal end. It has one gene per 88,999 bps and one candidate gene from every 85 genes. One average, there is one candidate gene per 7,556,067 bps. Three of the ten selected candidate genes showed association with human hypertension. Recently, it was reported that five polymorphisms in the KCNJ1 gene coding for the potas- sium channel, ROMK, showed associations with mean 24-hour systolic or diastolic blood pressure [25,46]. An- other candidate gene, Cytochrome P450, family 1, sub- family A, polypeptide 1 (CYP1A1) has been studied in several populations, and, particularly, the T6325C poly- morphism is thought to modulate essential hyperten- Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 144 sion-associated stroke risk [64]. Bp 184 is partially overlapped with Bp262 and has one gene per 101,726 bps with one candidate gene per 80 genes. One average, there is one candidate gene per 8,188,965 bps. Bp 263 has one gene per 82,646 bps. Six genes were selected as candidate genes, with one candidate gene per 71 genes. One average, one candidate gene is found per 5,900,984 bps. Only one candidate gene is associated with hypertension in human populations. In a study of the 206 M polymorphic variant of the SLC26A6 gene encoding a Cl(-)-oxalate transporter in patients with pri- mary hyperparathyroidism, Corbetta et al. [65] found that the SLC26A6 206M alleles were significantly re- lated to the presence of hypertension. Bp263 is essentially the same as Bp184 and SLC26A6 is also included as a candidate gene for this locus. 3.4.4. Chromosome 9 Chromosome 9 includes the two QTL Bp34 and Bp185. Bp34 is located on a region covering 37,780,765 bps. It contains as many as 311 genes, with an average of 121,482 bps per gene. However, there is an average of 5,397,252 bps per candidate gene. Of the candidate genes, Cytotoxic T-lymphocyte-associated protein 4 (CTLA4) is among the polymorphisms from 16 genes that are sig- nificantly associated with blood pressure variations (29) in the Japanese population. Bp185 cover a 40,000,158 bp region and has a total of 343 genes, with 116,618 bps per gene. It also has one candidate gene per 43 genes. On average, every 6,666,669 bps contains one candidate gene. Wen et al. [66] reported evidence for the association between a regulatory polymorphism in Secretogranin II (SCG2) and hypertension in African-American subjects. 3.4.5. Chromosome 14 Chromosome 14 includes the two QTL Bp189 and Bp59. Bp189 is located on a region covering 40,799,268 bps. It contains as many as 245 genes, with an average of 166,527 bps per gene. However, it only contains three candidate genes, which is an average of 13,599,756 bps per candidate gene. Dries et al. [67] showed that the Corin gene minor allele defined by 2 missense mutations is common in African-Americans and is associated with high blood pressure and hypertension. Their study was also confirmed in later study comprising the Chinese population [68]. Another of the candidate genes is ex- tracellular superoxide dismutase gene (SOD3). Based on the results from Naganuma et al. [69] who utilized a haplotype-based case-control study, a T-A haplotype of the SOD gene may be a genetic marker for essential hy- pertension. Bp59 covers a 15,236,276 bp region and has a total of 214 genes, with 71,197 bps per gene. It has one candi- date gene per 36 selected gene. On average, every 2,539,379 bps contains one candidate gene. Adducin (ADD1) is a candidate gene for salt-sensitive hyperten- sion. The association between ADD1 and essential hy- pertension has been well documented [70,71]. G pro- tein-coupled receptor kinases (GRKs) polymorphisms that lead to aberrant action of GRKs cause a number of conditions, which include hypertension and salt sensi- tive- ity. Polymorphisms in one particular member of this family, GRK4, have been shown to cause hyperphos- phorylation, desensitization, and internalization [72]. Yamada et al. suggested that the genotypes for ITGA2, GCK, and PTGIS may prove reliable for the assessment of the genetic component of hypertension. There claim was based on their study of a population comprised of 4853 unrelated Japanese individuals, including 2818 subjects with hypertension [73]. 3.4.6. Chromosome 18 Chromosome 18 includes the three QTL Bp2, Bp233, and Bp48. Bp2 is located on the proximal region of the chromo- some and consists of 33,239,845 bps. It contains as many as 282 genes, and has on average 117,871 bps per gene. However, there is an average of 16,619,922 bps per can- didate gene. There are only two candidate genes in this QTL, and only one (ROCK) with any linkage to EH in the human population. A Rho kinase (ROCK) polymer- phism influences blood pressure and systemic vascular resistance in human twins [74]. Bp233 covers a 40,000,179 bp region and has a total of 319 genes, with 125,392 bps per gene. It has one can- didate gene per 40 selected genes. On average, every 5,000,022 bps contains one candidate gene. Bp48 covers a 19,324,774 bp region and has a total of 128 genes, with 150,974 bps per gene. There is one can- didate gene per 43 selected genes. On average, every 6,441,591 bps contains one candidate gene. 3.5. Candidate Genes in the Peak Region of the QTL Compared to Candidate Genes in the QTL Region As indicated in our method, we selected QTL with LOD scores that are ≥2.8. However, QTL usually produce a curved or bell-shaped region. Thus, the LOD score at the top of the bell, the peak point, has the highest value. Theoretically, the higher a LOD score is the higher the probability there is a QTL. We then ask the question, if the LOD score in the peak region is high is the probabil- ity of a QTL gene in or near the peak point also high? Therefore, based on our assumptions we examined the number of candidate genes within 1.5 mbps on each side Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 145 of the peak region. As expected, there was a much high probability of candidate genes in those 1.5 mbp regions. In the whole rat genome, we obtained one candidate gene from every 12,594,138 bps. Of all the QTL, we have information on the peak positions of 45 QTL. From these 45 peak re- gions, we obtained 21 candidate genes (Table 1). On average, there are one candidate gene per 7.105 mbps in the peak region of a QTL. However, these 21 candidate genes are from only 13 QTL peak regions. The peak re- gions for the rest of 26 QTL do not contain obvious can- didate genes (Supplementary Table 1). Among these 13 peak regions, one region has three candidate genes while in each of other four regions there are 2 candidate genes (Table 1). There is no correlation between the number of candidate genes in a peak region and the number of total genes in a QTL region (R = –0.084). However, there is a weak negative correlation between the bps per candidate gene and the number of candidate genes in a peak region (R = –0.325). Overall, the number of candidate genes in a peak region appears independent from other factors in this study. Among all the QTL we examined, only Bp124 on chromosome 4 contained three candidate genes in its peak region. This QTL is located on a gene rich region with a LOD score of 3. It also contains a total of 456 genes with an average of 87,719 bps per gene. The three candidate genes from this QTL are from a pool of 10 candidates. The three candidates are Phosphodiesterase 1c (PDE1c), Aquaporin 1 (AQP1) and Corticotropin- Releasing Hormone Receptor 2 (CRHR2). Recent re- ports indicate that members in the calcium/calmodulin- dependent phosphodiesterase 1 (PDE1) family may play a major role in vascular smooth muscle cell proliferation. Particularly, PDE1c has been known to contribute to decreased cAMP production and increased proliferation of pulmonary areterial smooth muscle cells (PASMC) in patients with pulmonary hypertension (PHT) [75]. AQP1 is abundant in renal proximal tubular epithelium, the thin descending limb of the loop of Henle, and the descend- ing vasa recta of the kidney. In 2006, Lee et al. [76] re- ported that the expression of AQP1-3 channels is in- creased in the kidney in association with enhanced activ- ity of the AVP/cAMP pathway in spontaneously hyper- tensive rats. CRHR2 is known for its important role in coordinating the endocrine, autonomic, and behavioral responses to stress and immune challenges [77]. In a study of an Antalarmin blockade of corticotropin releasing hormone- induced hypertension in rats, Briscoe and colleagues [78] reported that the hypertension produced by central CRH administration is mediated through central CRHR1 re- ceptors, whereas, the hypertension produced by par- enteral CRH administration is mediated through periph- eral CRHR2 receptors. It would be very interesting to know whether all, or any of these three genes, are the causal genes for hypertension’s QTL. 4. CONCLUSIONS Our investigation raises an important issue in the selec- tion of candidate genes for specific QTLs, the considera- tion of the position of a gene in the QTL region. Our data suggests that a gene in the peak region of a QTL is of more relative importance than a gene further from the peak position. Theoretically, this agrees with the defi- nition of the LOD score. From experience, many QTL genes have been identified from positions that are close to the peak region of the QTL. The other issue our investigation discovered is whether candidate genes can be easily identified from a QTL lo- cated in either gene-rich or gene-poor regions. In our study, we found that QTL located in both gene-rich re- gions and gene-rich chromosomes contained the most candidate genes. However, the number of candidate genes within a peak region is not associated with the number of total genes in a QTL region. Also, more candidate genes does not necessary indicate that the real, causal gene is among these genes. Similarly, less candidate genes in the QTL of gene-poor regions or gene-poor chromosomes does not necessary mean that the analysis is easy due to the smaller number of candidate genes. The fact from our study is that there are an extremely large number of genome regions and genes covered by the hypertension QTLs. Even though our search for can- didate genes was limited to just essential hypertension, we still obtained 266 candidate genes. Considering the large number of QTL and genetic and environmental factors, identification of the QTL for hypertension with- out defined environmental conditions and a unified ge- nomic background is extremely difficult. 5. ACKNOWLEDGEMENTS Support for this work is from Center of Genomics and Bioinformatics (WG) and Center in Connective Tissue Research (WG), at University of Tennessee Health Science Center; Veterans Administration (WG); National Institute on Alcohol Abuse and Alcoholism (NIAAA), Na- tional Institutes of Health (1R01AA016342 to WG) and the Center of Integrating Genomics and Bioinformatics for International Studying of Strokes, the University of Tennessee Health Science Center, and the Veterans Administration Medical Center, Memphis, TN. Funding for YJW is from the “Major Project of Chinese National Programs for Fundamental Research and Development (No.2009CB521905).” REFERENCES [1] Chauvet, C., Ménard, A., Xiao, C., Aguila, B., Blain, M., Roy, J. and Deng, A.Y. (2012) Novel genes as primary Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 146 triggers for polygenic hypertension. Journal of Hyperten- sion, 30, 81-86. doi:10.1097/HJH.0b013e32834dddb1 [2] Simino, J., Shi, G., Kume, R., Schwander, K., Province, M.A., Gu, C.C., Kardia, S., Chakravarti, A., Ehret, G., Olshen, R.A., Turner, S.T., Ho, L.T., Zhu, X., Jaquish, C., Paltoo, D., Cooper, R.S., Weder, A., Curb, J.D., Boerwin- kle, E., Hunt, S.C. and Rao, D.C. (2011) Five blood pressure loci identified by an updated genome-wide link- age scan: Meta-analysis of the Family Blood Pressure Program. American Journal of Hypertension, 24, 347- 354. doi:10.1038/ajh.2010.238 [3] Johnson, M.D., He, L., Herman, D., Wakimoto, H., Wal- lace, C.A., Zidek, V., Mlejnek, P., Musilova, A., Si- makova, M., Vorlicek, J., Kren, V., Viklicky, O., Qi, N.R., Wang, J., Seidman, C.E., Seidman, J., Kurtz, T.W., Aitman, T.J. and Pravenec, M. (2009) Dissection of chromosome 18 blood pressure and salt-sensitivity quan- titative trait loci in the spontaneously hypertensive rat. Hypertension, 54, 639-645. doi:10.1161/HYPERTENSIONAHA.108.126664 [4] Deng, A.Y. (2007) Genetic basis of polygenic hyperten- sion. Human Molecular Genetics, 16, R195-R202. doi:10.1093/hmg/ddm126 [5] Cowley Jr., A.W. (2006) The genetic dissection of essen- tial hypertension. Nature Reviews Genetics, 7, 829-840. doi:10.1038/nrg1967 [6] Twigger, S.N., Shimoyama, M., Bromberg, S., Kwitek, A.E., Jacob, H.J. and RGD Team (2007) The Rat Gen- ome Database, update 2007—Easing the path from dis- ease to data and back again. Nucleic Acids Research, 35, D658-D662. doi:10.1093/nar/gkl988 [7] Tobin, M.D., Tomaszewski, M., Braund, P.S., Hajat, C., Raleigh, S.M., Palmer, T.M., Caulfield, M., Burton, P.R. and Samani, N.J. (2008) Common variants in genes un- derlying monogenic hypertension and hypotension and blood pressure in the general population. Hypertension, 51, 1658-1664. doi:10.1161/HYPERTENSIONAHA.108.112664 [8] Thomas, G.N., Critchley, J.A., Tomlinson, B., Cockram, C.S. and Chan, J.C. (2001) Relationships between the taqI polymorphism of the dopamine D2 receptor and blood pressure in hyperglycaemic and normoglycaemic Chinese subjects. Clinical Endocrinology, 55, 605-611. [9] Xiong, Q., Qiu, Y. and Gu, W. (2008) PGMapper: A web-based tool linking phenotype to genes. Bioinformat- ics, 24, 1011-1013. doi:10.1093/bioinformatics/btn002 [10] Lander, E. and Kruglyak, L. (1995) Genetic dissection of complex traits: Guidelines for interpreting and reporting linkage results. Nature Genetics, 11, 241-247. doi:10.1038/ng1195-241 [11] Van Ooijen, J.W. (1992) Accuracy of mapping quan- titative trait loci in autogamous species. TAG Theoretical and Applied Genetics, 84, 9. [12] Yasui, N., Kajimoto, K., Sumiya, T., Okuda, T. and Iwai, N. (2007) The monocyte chemotactic protein-1 gene may contribute to hypertension in Dahl salt-sensitive rats. Hy- pertension Research, 30, 185-193. doi:10.1291/hypres.30.185 [13] Denag, A.Y. (1998) Is the nitric oxide system involved in genetic hypertension in Dahl rats? Kidney International, 53, 1501-1511. doi:10.1046/j.1523-1755.1998.00933.x [14] Ye, P. and West, M.J. (2003) Cosegregation analysis of natriuretic peptide genes and blood pressure in the spon- taneously hypertensive rat. Clinical and Experimental Pharmacology and Physiology, 30, 930-936. doi:10.1111/j.1440-1681.2003.03937.x [15] Suthanthiran, M., Li, B., Song, J.O., Ding, R., Sharma, V.K., Schwartz, J.E. and August, P. (2000) Transforming growth factor-beta-1 hyperexpression in African-Ameri- can hypertensives: A novel mediator of hypertension and/or target organ damage. Proceedings of the National Academy of Sciences, 97, 3479-3484. [16] Petretto, E., Sarwar, R., Grieve, I., Lu, H., Kumaran, M.K., Muckett, P.J., Mangion, J., Schroen, B., Benson, M., Punjabi, P.P., Prasad, S.K., Pennell, D.J., Kiesewetter, C., Tasheva, E.S., Corpuz, L.M., Webb, M.D., Conrad, G.W., Kurtz, T.W., Kren, V., Fischer, J., Hubner, N., Pinto, Y.M., Pravenec, M., Aitman, T.J. and Cook, S.A. (2008) Integrated genomic approaches implicate osteo- glycin (Ogn) in the regulation of left ventricular mass. Nature Genetics, 40, 546-552. doi:10.1038/ng.134 [17] Frikke-Schmidt, R., Sing, C.F., Nordestgaard, B.G., Stef- fensen, R. and Tybjaerg-Hansen, A. (2007) Subsets of SNPs define rare genotype classes that predict ischemic heart disease. Human Genetics, 120, 865-877. doi:10.1007/s00439-006-0233-y [18] Koo, B.K., Cho, Y.M., Park, B.L., Cheong, H.S., Shin, H.D., Jang, H.C., Kim, S.Y., Lee, H.K. and Park, K.S. (2007) Polymorphisms of KCNJ11 (Kir6.2 gene) are as- sociated with Type 2 diabetes and hypertension in the Korean population. Diabetic Medicine, 24, 178-186. doi:10.1111/j.1464-5491.2006.02050.x [19] Kokubo, Y., Tomoike, H., Tanaka, C., Banno, M., Okuda, T., Inamoto, N., Kamide, K., Kawano, Y. and Miyata, T. (2006) Association of sixty-one non-synonymous poly- merphisms in forty-one hypertension candidate genes with blood pressure variation and hypertension. Hyper- tension Research, 29, 611-619. doi:10.1291/hypres.29.611 [20] Iwai, N., Tago, N., Yasui, N., Kokubo, Y., Inamoto, N., Tomoike, H. and Shioji, K. (2004) Genetic analysis of 22 candidate genes for hypertension in the Japanese popu- lation. Journal of Hypertension, 22, 1119-1126. doi:10.1097/00004872-200406000-00012 [21] Busjahn, A., Aydin, A., Uhlmann, R., Krasko, C., Bäh- ring, S., Szelestei, T., Feng, Y., Dahm, S., Sharma, A.M., Luft, F.C. and Lang, F. (2002) Serum- and glucocorti- coid-regulated kinase (SGK1) gene and blood pressure. Hypertension, 40, 256-260. doi:10.1161/01.HYP.0000030153.19366.26 [22] Trochen, N., Ganapathipillai, S., Ferrari, P., Frey, B.M. and Frey, F.J. (2004) Low prevalence of nonconservative mutations of serum and glucocorticoid-regulated kinase (SGK1) gene in hypertensive and renal patients. Ne- phrology Dialysis Transplantation, 19, 2499-2504. doi:10.1093/ndt/gfh417 [23] Lazar, A., Zimmermann, T., Koch, W., Gründemann, D., Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 147 Schömig, A., Kastrati, A. and Schömig, E. (2006) Lower prevalence of the OCT2 Ser270 allele in patients with essential hypertension. Clinical and Experimental Hy- pertension, 28, 645-653. doi:10.1080/10641960600946411 [24] Wang, L., Feng, Y., Zhang, Y., Zhou, H., Jiang, S., Niu, T., Wei, L.J., Xu, X., Xu, X. and Wang, X. (2006) Proly- lcarboxypeptidase gene, chronic hypertension, and risk of preeclampsia. American Journal of Obstetrics & Gyne- cology, 195, 162-171. doi:10.1016/j.ajog.2006.01.079 [25] Tobin, M.D., Tomaszewski, M., Braund PS, Hajat C, Raleigh SM, Palmer TM, Caulfield, M., Burton, P.R. and Samani, N.J. (2008) Common variants in genes under- lying monogenic hypertension and hypotension and blood pressure in the general population. Hypertension, 51, 1658-1664. doi:10.1161/HYPERTENSIONAHA.108.112664 [26] Büsst, C.J., Scurrah, K.J., Ellis, J.A. and Harrap, S.B. (2007) Selective genotyping reveals association between the epithelial sodium channel gamma-subunit and systolic blood pressure. Hypertension, 50, 672-678. doi:10.1161/HYPERTENSIONAHA.107.089128 [27] Iwai, N., Baba, S., Mannami, T., Katsuya, T., Higaki, J., Ogihara, T. and Ogata, J. (2001) Association of sodium channel gamma-subunit promoter variant with blood pressure. Hypertension, 38, 86-89. doi:10.1161/01.HYP.38.1.86 [28] Sáez, M.E., Martínez-Larrad, M.T., Ramírez-Lorca, R., González-Sánchez, J.L., Zabena, C., Martinez-Calatrava, M.J., González, A., Morón, F.J., Ruiz, A. and Serrano- Ríos, M. (2007) Calpain-5 gene variants are associated with diastolic blood pressure and cholesterol levels. BMC Medical Genetics, 8, 1. [29] Iwai, N., Kajimoto, K., Kokubo, Y. and Tomoike, H. (2006) Extensive genetic analysis of 10 candidate genes for hypertension in Japanese. Hypertension, 48, 901-907. doi:10.1161/01.HYP.0000242485.23148.bb [30] Ji, Q., Ikegami, H., Fujisawa, T., Kawabata, Y., Ono, M., Nishino, M., Ohishi, M., Katsuya, T., Rakugi, H. and Ogihara, T. (2004) A common polymorphism of uncou- pling protein 2 gene is associated with hypertension. Journal of Hypertension, 22, 97-102. doi:10.1097/00004872-200401000-00018 [31] Matsunaga, T., Gu, N., Yamazaki, H., Tsuda, M., Adachi, T., Yasuda, K., Moritani, T., Tsuda, K., Nonaka, M. and Nishiyama, T. (2009) Association of UCP2 and UCP3 polymorphisms with heart rate variability in Japanese men. Journal of Hypertension, 27, 305-313. doi:10.1097/HJH.0b013e32831ac967 [32] Marçano, A.C., Burke, B., Gungadoo, J., Wallace, C., Kaisaki, P.J., Woon, P.Y., Farrall, M., Clayton, D., Brown, M., Dominiczak, A., Connell, J.M., Webster, J., Lathrop, M., Caulfield, M., Samani, N., Gauguier, D., Munroe, P.B. (2007) Genetic association analysis of inosi- tol polyphosphate phosphatase-like 1 (INPPL1, SHIP2) variants with essential hypertension. Journal of Medical Genetics, 44, 603-605. doi:10.1136/jmg.2007.049718 [33] Gaukrodger, N., Avery, P.J. and Keavney, B. (2008) Plasma potassium level is associated with common ge- netic variation in the beta-subunit of the epithelial sodium channel. American Journal of Physiology—Regulatory, Integrative and Comparative Physiology, 294, R1068- R1072. doi:10.1152/ajpregu.00732.2007 [34] Jin, Y., Kuznetsova, T., Tikhonoff, V., Thijs, L., Hasenk- amp, S., Bäumer, V., Stolarz-Skrzypek, K., Ryabikov, A., Richart, T., Malyutina, S., Nikitin, Y., Casiglia, E., Kawecka-Jaszcz, K., Olszanecka, A., Brand-Herrmann, S.M., Brand, E., Fagard, R., Staessen, J.A. and European Project on Genes in Hypertension (EPOGH) Investigators (2009) Left ventricular structure in relation to the human SAH gene in the European Project on Genes in Hyper- tension. Hypertension Research, 32, 145-151. doi:10.1038/hr.2008.30 [35] Tikhonoff, V., Staessen, J.A., Kuznetsova, T., Thijs, L., Hasenkamp, S., Bäumer, V., Stolarz, K., Seidlerová, J., Filipovský, J., Nikitin, Y., Peleska, J., Kawecka-Jaszcz, K., Casiglia, E., Brand-Herrmann, S.M., Brand, E. and European Project on Genes in Hypertension (EPOGH) Investigators (2008) SAH gene variants revisited in the European Project On Genes in Hypertension. Journal of Hypertension, 26, 244-250. doi:10.1097/HJH.0b013e3282f2d36f [36] Yuan, Z., Su, Z., Miyoshi, T., Rowlan, J.S. and Shi, W. (2008) Quantitative trait locus analysis of circulating ad- hesion molecules in hyperlipidemic apolipoprotein E-de- ficient mice. Molecular Genetics and Genomics, 280, 375-383. doi:10.1007/s00438-008-0371-0 [37] Okuda, T., Sumiya, T., Mizutani, K., Tago, N., Miyata, T., Tanabe, T., Kato, H., Katsuya, T., Higaki, J., Ogihara, T., Tsujita, Y. and Iwai, N. (2002) Analyses of differential gene expression in genetic hypertensive rats by microa- rray. Hypertension Research, 25, 249-255. doi:10.1291/hypres.25.249 [38] Tripodi, G., Modica, R., Stella, A., Bigatti, G., Bianchi, G. and Stella, P. (2005) Haplotype analysis of carnitine transporters and left ventricular mass in human essential hypertension. Journal of Renal Nutrition, 15, 2-7. doi:10.1053/j.jrn.2004.09.035 [39] Kohara, K., Tabara, Y., Nakura, J., Imai, Y., Ohkubo, T., Hata, A., Soma, M., Nakayama, T., Umemura, S., Hirawa, N., Ueshima, H., Kita, Y., Ogihara, T., Katsuya, T., Taka- hashi, N., Tokunaga, K. and Miki, T. (2008) Identifica- tion of hypertension-susceptibility genes and pathways by a systemic multiple candidate gene approach: The mil- lennium genome project for hypertension. Hypertension Research, 31, 203-212. doi:10.1291/hypres.31.203 [40] Kawaguchi, Y., Tochimoto, A., Hara, M., Kawamoto, M., Sugiura, T., Katsumata, Y., Okada, J., Kondo, H., Okubo, M. and Kamatani, N. (2006) NOS2 polymorphisms asso- ciated with the susceptibility to pulmonary arterial hy- pertension with systemic sclerosis: Contribution to the transcriptional activity. Arthritis Research & Therapy, 8, R104. doi:10.1186/ar1984 [41] Jemaa, R., Ben, A.S., Kallel, A., Omar, S., Feki, M., El- asmi, M., Haj-Taïeb, S., Sanhaji, H. and Kaabachi, N. (2009) Association between the -2518G/A polymorphism in the monocyte chemoattractant protein-1 (MCP-1) gene Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 148 and hypertension in Tunisian patients. Clinical Biochem- istry, 42, 34-37. doi:10.1016/j.clinbiochem.2008.09.118 [42] Quintana, L.F., Guzmán, B., Collado, S., Clària, J. and Poch, E. (2006) A coding polymorphism in the 12-lipo- xygenase gene is associated to essential hypertension and urinary 12(S)-HETE. Kidney International, 69, 526-530. doi:10.1038/sj.ki.5000147 [43] Omi, T., Kumada, M., Kamesaki, T., Okuda, H., Mun- khtulga, L., Yanagisawa, Y., Utsumi, N., Gotoh, T., Hata, A., Soma, M., Umemura, S., Ogihara, T., Takahashi, N., Tabara, Y., Shimada, K., Mano, H., Kajii, E., Miki, T. and Iwamoto, S. (2006) An intronic variable number of tandem repeat polymorphisms of the cold-induced auto- inflammatory syndrome 1 (CIAS1) gene modifies gene expression and is associated with essential hypertension. European Journal of Human Genetics, 14, 1295-1305. doi:10.1038/sj.ejhg.5201698 [44] Timasheva, Y.R., Nasibullin, T.R., Zakirova, A.N. and Mustafina, O.E. (2008) Association of interleukin-6, in- terleukin-12, and interleukin-10 gene polymorphisms with essential hypertension in Tatars from Russia. Biochemical Genetics, 46, 64-74. doi:10.1007/s10528-007-9130-x [45] Sink, K.M., Leng, X., Williamson, J., Kritchevsky, S.B., Yaffe, K., Kuller, L., Yasar, S., Atkinson, H., Robbins, M., Psaty, B. and Goff Jr., D.C. (2009) Angiotensin- converting enzyme inhibitors and cognitive decline in older adults with hypertension: Results from the Cardio- vascular Health Study. Archives of Internal Medicine, 169, 1195-1202. doi:10.1001/archinternmed.2009.175 [46] Woo, S.W., Bang, S., Chung, M.W., Jin, S.K., Kim, Y.S. and Lee, S.H. (2009) Lack of association between ACE and bradykinin B2 receptor gene polymorphisms and ACE inhibitor-induced coughing in hypertensive Koreans. Journal of Clinical Pharmacy and Therapeutics, 34, 561-567. doi:10.1111/j.1365-2710.2009.01028.x [47] Kokubo, Y., Tomoike, H., Tanaka, C., Banno, M., Okuda, T., Inamoto, N., Kamide, K., Kawano, Y. and Miyata, T. (2006) Association of sixty-one non-synonymous poly- morphisms in forty-one hypertension candidate genes with blood pressure variation and hypertension. Hyper- tension Research, 29, 611-619. doi:10.1291/hypres.29.611 [48] Palijan, A., Lambert, R., Dutil, J., Sivo, Z. and Deng, A.Y. (2003) Comprehensive congenic coverage revealing mul- tiple blood pressure quantitative trait loci on Dahl rat chromosome 10. Hypertension, 42, 515-522. doi:10.1161/01.HYP.0000090096.88509.15 [49] Jung, J., Foroud, T.M., Eckert, G.J., Flury-Wetherill, L., Edenberg, H.J., Xuei, X., Zaidi, S.A. and Pratt, J.H. (2009) Association of the calcium-sensing receptor gene with blood pressure and urinary calcium in African- Americans. Journal of Clinical Endocrinology & Me- tabolism, 94, 1042-1048. doi:10.1210/jc.2008-1861 [50] Tobin, M.D., Tomaszewski, M., Braund, P.S., Hajat, C., Raleigh, S.M., Palmer, T.M., Caulfield, M., Burton, P.R. and Samani, N.J. (2008) Common variants in genes un- derlying monogenic hypertension and hypotension and blood pressure in the general population. Hypertension, 51, 1658-1664. doi:10.1161/HYPERTENSIONAHA.108.112664 [51] Wessel, J., Topol, E.J., Ji, M., Meyer, J. and McCarthy, J.J. (2004) Replication of the association between the thrombospondin-4 A387P polymorphism and myocardial infarction. American Heart Journal, 147, 905-909. doi:10.1016/j.ahj.2003.12.013 [52] Gigante, B., Bellis, A., Visconti, R., Marino, M., Morisco, C., Trimarco, V., Galasso, G., Piscione, F., De Luca, N., Prince, J.A., de Faire, U. and Trimarco, B. (2007) Retro- spective analysis of coagulation factor II receptor (F2R) sequence variation and coronary heart disease in hyper- tensive patients. Arteriosclerosis, Thrombosis, and Vas- cular Biology, 27, 1213-1219. doi:10.1161/ATVBAHA.107.140541 [53] Pitzalis, M.V., Sarzani, R., Dessì-Fulgheri, P., Iacoviello, M., Forleo, C., Lucarelli, K., Pietrucci, F., Salvi, F., Sor- rentino, S., Romito, R., Guida, P., Rappelli, A. and Riz- zon, P. (2003) Allelic variants of natriuretic peptide re- ceptor genes are associated with family history of hyper- tension and cardiovascular phenotype. Journal of Hyper- tension, 21, 1491-1496. doi:10.1097/00004872-200308000-00012 [54] Horan, M., Newsway, V., Yasmin, Lewis, M.D., Easter, T.E., Rees, D.A., Mahto, A., Millar, D.S., Procter, A.M., Scanlon, M.F., Wilkinson, I.B., Hall, I.P., Wheatley, A., Blakey, J., Bath, P.M., Cockcroft, J.R., Krawczak, M. and Cooper, D.N. (2006) Genetic variation at the growth hormone (GH1) and growth hormone receptor (GHR) loci as a risk factor for hypertension and stroke. Human Genetics, 119, 527-540. doi:10.1007/s00439-006-0166-5 [55] Rice, T., Rankinen, T., Province, M.A., Chagnon, Y.C., Pérusse, L., Borecki, I.B., Bouchard, C. and Rao, D.C. (2000) Genome-wide linkage analysis of systolic and diastolic blood pressure: The Québec Family Study. Cir- culation, 102, 1956-1963. doi:10.1161/01.CIR.102.16.1956 [56] Capoluongo, E., Onder, G., Concolino, P., Russo, A., Santonocito, C., Bernabei, R., Zuppi, C., Ameglio, F. and Landi, F. (2009) GSTM1-null polymorphism as possible risk marker for hypertension: results from the aging and longevity study in the Sirente Geographic Area (ilSI- RENTE study). Clinica Chimica Acta, 399, 92-96. doi:10.1016/j.cca.2008.09.017 [57] Woon, P.Y., Kaisaki, P.J., Bragança, J., Bihoreau, M.T., Levy, J.C., Farrall, M. and Gauguier, D. (2007) Aryl hy- drocarbon receptor nuclear translocator-like (BMAL1) is associated with susceptibility to hypertension and type 2 diabetes. Proceedings of the National Academy of Sci- ences, 104, 14412-14417. doi:10.1073/pnas.0703247104 [58] Liu, J., Li, M., Qi, M.L., Liu, X.L., Li, Y.H., Fu, L.Y. and Zhao, Y.Y. (2007) Association of -63A/C polymorphism of glutathione S-transferase M3 gene with essential hy- pertension in Chinese population. Chinese Journal of Medical Genetics, 24, 582-585. [59] Delles, C., Padmanabhan, S., Lee, W.K., Miller, W.H., Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 149 McBride, M.W., McClure, J.D., Brain, N.J., Wallace, C., Marçano, A.C., Schmieder, R.E., Brown, M.J., Caulfield, M.J., Munroe, P.B., Farrall, M., Webster, J., Connell, J.M. and Dominiczak, A.F. (2008) Glutathione S-transferase variants and hypertension. Journal of Hypertension, 26, 1343-1352. doi:10.1097/HJH.0b013e3282fe1d67 [60] Motone, M., Katsuya, T., Ishikawa, K., Iwashima, Y., Sugimoto, K., Yamamoto, K., Fu, Y., Matsuo, A., Ohishi, M., Rakugi, H. and Ogihara, T. (2004) Association be- tween hepatocyte growth factor gene polymorphism and essential hypertension. Hypertension Research, 27, 247- 251. doi:10.1291/hypres.27.247 [61] Takiuchi, S., Mannami, T., Miyata, T., Kamide, K., Ta- naka, C., Kokubo, Y., Koyama, Y., Inamoto, N., Katsuya, T., Iwai, N., Kawano, Y., Ogihara, T. and Tomoike, H. (2004) Identification of 21 single nucleotide polymer- phisms in human hepatocyte growth factor gene and as- sociation with blood pressure and carotid atherosclerosis in the Japanese population. Atherosclerosis, 173, 301-307. doi:10.1016/j.atherosclerosis.2003.12.020 [62] Genelhu, V.A., Celoria, B.M., Pimentel, M.M., Duarte, S.F., Cabello, P.H. and Francischetti, E.A. (2009) Asso- ciation of a common variant of the leptin gene with blood pressure in an obese Brazilian population. American Journal of Hypertension, 22, 577-580. doi:10.1038/ajh.2009.7 [63] Hou, X.W., Wang, L.F., Wang, N., Pang, D., Hui, B., Zhou, Y.L. and He, X. (2008) The G501C polymorphism of oxidized LDL receptor gene [OLR-1] is associated with susceptibility and serum C-reactive protein concen- tration in Chinese essential hypertensives. Clinica Chi- mica Acta, 388, 200-203. doi:10.1016/j.cca.2007.11.016 [64] Lança, V., Alcântara, P., Braz-Nogueira, J. and Bicho, M.P. (2004) Cytochrome P450 1A1 (CYP1A1) T6325C polymorphism might modulate essential hypertension- associated stroke risk. Revista Portuguesa de Cardiologia, 23, 343-355. [65] Corbetta, S., Eller-Vainicher, C., Frigerio, M., Valaperta, R., Costa, E., Vicentini, L., Baccarelli, A., Beck-Peccoz, P. and Spada, A. (2009) Analysis of the 206 M polymer- phic variant of the SLC26A6 gene encoding a Cloxalate transporter in patients with primary hyperparathyroidism. European Journal of Endocrinology, 160, 283-288. doi:10.1530/EJE-08-0623 [66] Wen, G., Wessel, J., Zhou, W., Ehret, G.B., Rao, F., Stridsberg, M., Mahata, S.K., Gent, P.M., Das, M., Coo- per, R.S., Chakravarti, A., Zhou, H., Schork, N.J., O’Connor, D.T. and Hamilton, B.A. (2007) An ancestral variant of Secretogranin II confers regulation by PHOX2 transcription factors and association with hypertension. Human Molecular Genetics, 16, 1752-1764. doi:10.1093/hmg/ddm123 [67] Dries, D.L., Victor, R.G., Rame, J.E., Cooper, R.S., Wu, X., Zhu, X., Leonard, D., Ho, S.I., Wu, Q., Post, W. and Drazner, M.H. (2005) Corin gene minor allele defined by 2 missense mutations is common in blacks and associated with high blood pressure and hypertension. Circulation, 112, 2403-2410. doi:10.1161/CIRCULATIONAHA.105.568881 [68] Wang, W., Liao, X., Fukuda, K., Knappe, S., Wu, F., Dries, D.L., Qin, J. and Wu, Q. (2008) Corin variant as- sociated with hypertension and cardiac hypertrophy ex- hibits impaired zymogen activation and natriuretic pep- tide processing activity. Circulation Research, 103, 502- 508. doi:10.1161/CIRCRESAHA.108.177352 [69] Naganuma, T., Nakayama, T., Sato, N., Fu, Z., Soma, M., Aoi, N. and Usami, R. (2008) A haplotype-based case- control study examining human extracellular superoxide dismutase gene and essential hypertension. Hypertension Research, 31, 1533-1540. doi:10.1291/hypres.31.1533 [70] Manunta, P. and Bianchi, G. (2006) Pharmacogenomics and pharmacogenetics of hypertension: Update and per- spectives—The adducin paradigm. Journal of the Ameri- can Society of Nephrology, 17, S30-S35. doi:10.1681/ASN.2005121346 [71] Mandò, C., Antonazzo, P., Tabano, S., Zanutto, S., Pileri, P., Somigliana, E., Colleoni, F., Martinelli, A., Zolin, A., Benedetto, C., Marozio, L., Neri, I., Facchinetti, F., Miozzo, M. and Cetin, I. (2009) Angiotensin-converting enzyme and adducin-1 polymorphisms in women with preeclampsia and gestational hypertension. Reproductive Sciences, 16, 819-826. doi:10.1177/1933719109336612 [72] Cohn, H.I., Xi, Y., Pesant, S., Harris, D.M., Hyslop, T., Falkner, B. and Eckhart, A.D. (2009) G protein-coupled receptor kinase 2 expression and activity are associated with blood pressure in black Americans. Hypertension, 54, 71-76. doi:10.1161/HYPERTENSIONAHA.108.125955 [73] Yamada, Y., Matsuo, H., Segawa, T., Watanabe, S., Kato, K., Hibino, T., Yokoi, K., Ichihara, S., Metoki, N., Yo- shida, H., Satoh, K. and Nozawa, Y. (2006) Assessment of the genetic component of hypertension. American Journal of Hypertension, 19, 1158-1165. doi:10.1016/j.amjhyper.2006.04.010 [74] Seasholtz, T.M., Wessel, J., Rao, F., Rana, B.K., Khan- drika, S., Kennedy, B.P., Lillie, E.O., Ziegler, M.G., Smith, D.W., Schork, N.J., Brown, J.H. and O’Connor, D.T. (2006) Rho kinase polymorphism influences blood pressure and systemic vascular resistance in human twins: role of heredity. Hypertension, 47, 937-947. doi:10.1161/01.HYP.0000217364.45622.f0 [75] Murray, F., Patel, H.H., Suda, R.Y., Zhang, S., This- tlethwaite, P.A., Yuan, J.X. and Insel, P.A. (2007) Ex- pression and activity of cAMP phosphodiesterase iso- forms in pulmonary artery smooth muscle cells from pa- tients with pulmonary hypertension: Role for PDE1. American Journal of Physiology—Lung Cellular and Molecular Physiology, 292, L294-L303. doi:10.1152/ajplung.00190.2006 [76] Lee, J., Kim, S., Kim, J., Jeong, M.H., Oh, Y. and Choi, K.C. (2006) Increased expression of renal aquaporin wa- ter channels in spontaneously hypertensive rats. Kidney and Blood Pressure Research, 29, 18-23. doi:10.1159/000092483 [77] Liaw, C.W., Lovenberg, T.W., Barry, G., Oltersdorf, T., Grigoriadis, D.E. and De Souza, E.B. (1996) Cloning and characterization of the human corticotropin-releasing Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 Copyright © 2012 SciRes. 150 OPEN ACCESS factor-2 receptor complementary deoxyribonucleic acid. Endocrinology, 137, 72-77. doi:10.1210/en.137.1.72 [78] Briscoe, R.J., Cabrera, C.L., Baird, T.J., Rice, K.C. and Woods, J.H. (2000) Antalarmin blockade of corticotropin releasing hormone-induced hypertension in rats. Brain Research, 881, 204-207. doi:10.1016/S0006-8993(00)02742-6  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 151 Supplementary Ta ble 1. Candidate Genes of hypertension in QTL on chromosomes. QTL Markers Search region (bp) Total genes Candidate genes (Essential and secondary) Candidate genes within 1.5 M/ polymorphic genes Bp95 D1UIA8, D1Rat18 0 - 22,869,052 248 total 194 known 38 novel 16 predicted Ogn, Gprk6, Tgfb1, Lama2(2nd) No/2 Bp96 D1Mgh2, D1Mit11 22,842,962 - 102,532,417 1211 total 946 known 138 novel Tgfb1, Abcc8, Apoe, Kcnj11, Abcc6(2nd), Nphs1(2nd), Akt2(2nd), Ptgir(2nd), Vip, Sgk1, Trpm4, Sod2, Tph1, Axl, Slc22a2, Zfp260, Myadm Tctex 1, Boat1, Slc9a3(2nd), Sdha(2nd) No/5+1 Bp173 D1Rat33 D1Rat130 115,946,375 - 189,900,838 954 total 772known, 120 novel 62 predicted 29. Iqgap1(2nd), Prc1, Fah, Plin, Ntrk3, Nr2f2, Prcp, Nox4, Scnn1g, Nmb Capn5, Anpep, Serpinh1, Pak1, Igf1r. Umod, Ucp2, Ucp3, Pde3b(2nd,), spon1, Inppl1, Phox2a, P2ry2, Scnn1b, Fgfr2, Sah, Xylt1, Hbb, Acsm3 No/10 Bp196 D1Mgh10, D1Mgh1 119,780,561 - 204,280,279 1146 946 known, 128 novel, 172 predicted Nox, Nmb, Capn5, Anpep, Serpinh1, Pak1, Fah, Iqgap1(2nd), Prc1, Plin, Nr2f2, Igf1r Scnn1b, Scnn1g, Fgfr2, Umod, Xylt1, Hbb, Prcp, Dhcr7, Th, Acsm3, Adm , No./4 Bp255 D1Rat208, D1Rat307 195,927,908 - 240,927,908 915 total 749 known 66 novel 80 predicted Dhcr7, Th, Cyp17a1, Bbs1(2nd,) Jak2, Gnaq, Adrbk1, Ccnd1, Cyp2c23, Pax2(2nd), Cyp2e1, Kcnq1, Cpt1a, Gldc(2nd) Peak marker is NA. Bp259 D1Rat71, D1Mgh12 216,663,010 - 247,322,280 262 total 195 known 40 novel 27 predicted Gldc(2nd), Pten, Rbp4(2nd), Mbl2, Kcnv2, Gna14, Gnaq, Jak2 Rbp4: 0.29 M./1 Bp115 D2Uia17 (peak) 0 - 33,349,984 218 total 138 known 49 novel 31 predicted Cast, Arts1, Thbs4, F2r, Pik3r1(2nd), No./3 Bp205 D2Rat73, D2Mgh14 26,101,089 - 42,866,342 109 total 80 known 19 novel 10 predicted No. NA Bp14 D2Mgh12, D2Mgh14 42,866,342 - 210,636,009 1284 total 939 known 216 novel 129 predicted Lmna, Fga, ATP1a, Npr1, Gucy1a3, Gucy1b3, Fgf2, Bbs7, Npr3, Lifr(2nd), Ghr, Ca2, Cp, Crh, Itga2, Gstm1, Arnt, Gstm2, Gstm3, Hsd3b1, Dear, Bche, PLD, Gdnf, Gstm4, Gstm5,, Kcna2, Gucy1a3, Kcnab1, Terc, Ccl28, Trpc3(2nd), Peak marker is not available/9 Bp203 D2Mit14, D2Mgh29 197,256,224 - 227,310,008 267 Total 210 known 34 novel 23 predicted Edg1, Kcna10, Vcam1, F3, Gstm1, Gstm2, Gstm3, Gstm4, Gstm5,, Kcna2, Edg1 (1.22 M)/0 + 1 Bp175 222,939,299 - 256,217,383 220 total 155 known 35 novel 30 predicted Cyr61, Edg7, Mttp, Ddah1 No. Bp15 D2Mgh12, D2Mgh14 0 - 26,373,454 402 total 333 known 42 novel 27 predicted Dbh, Pax8, Kynu, Ptgs1 or COX1, Vav2, Il1r(2nd), No/1 Bp264 D3Rat54, D3Rat17 10,267,753 - 121,619,110 1274 total 1039 known 146 novel 88 predicted Ptgs1 or COX1, Kynu, Avp, Nphp1(2nd), Cat, Bbs5, Pcna(2nd), Scn7a, Dpp4, Itgav Bbs5 (1.27 M)/2+1 Copyright © 2012 SciRes. OPEN ACCESS  L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 152 Continued Bp207 D3Mit49 (Peak marker) 111,188,559 - 151,188,749 495 total 391 known 67 novel 37 predicted Jag1(2nd), MKKS(2nd), Avp, Nphp1, Src, Il1b, Peak marker is not available/0+1 Bp81 D3Mgh2 (peak marker) 138,037,867 - 178,038,027 510 total 404 known 71 novel 35 predicted Src(2nd) Ptpn1, Hnf4a, Ada(2nd), Ahcy(2nd), Hrh3(2nd), Mmp9(2nd), Myh7b_predicted(2nd) Ptpn1 ptgds (1.03M) Bp179 D4Rat12, D4Mgh2 6,178,308 - 46,178,440 239 total 159 known 61 novel 19 predicted Nos3, Hgf, Cav(2nd), Pon-1, Cftr(2nd), Pon-2, ABCB1,, Colna2, Sema3a, Lep No/2 Bp86 D4Mit2, D4Mit11 55,369,932 - 91,330,856 390 total 325 known 43 novel 22 predicted 10+1. Lep, Npy, Pde1c(2nd), Trpv5, Aqp1, Crhr2, Hoxa9, Hoxa5(2nd), Casp2, Mtpn, Tbxas1(2nd), Peak marker is not available/0+1 Bp124 D4Rat34 (peak marker) 65,014,032 - 105,014,222 456 total 294 known 124 novel 38 predicted Npy, Pde1c, Trpv5, Aqp1, Crhr2, Hoxa9, Hoxa5, Casp2, Cntnap2, Fabp1, Pde1c(.077 M), Aqp1(.92 M) crhr2(1.17 M) Bp209 D4Mgh12 (peak marker) 163,954,705 - 203,954,820 251 total Lrp6, Pded3a(2nd), Olr1, No/1 Bp119 D5Mgh2 (peak) 0 - 37,517,235 214 total 192 known 40 novel 22 predicted Slc26a7, No Bp254 D5Rat9, D5Rat108 61,080,143 - 134,872,086 506 total 371 known 90 novel 45predicted Abca1, Tek(2nd), Cyp2j4 No/1 Bp147 D5Rat41(pe ak marker) 135,450,641 - 175,450,769 744 total 606 known 89 novel 49 predicted Ece1, Nppa, Tnfrsf1b, Cda(2nd), Slc9a1, Nppb, Uts2, Alpl, Cyp4a1, Slc2a5, Edn2(2nd), Tnfrsf4, Fabp3, Guca2b, Ptpru, Clcnkb No/6 BpQTL cluster7 D6Mit3, D6Mit12( D6Rat212 flanking marker) 14,873,099 - 75,049,528 372 total 255 known 69 novel 48 predicted 9. Id2(2nd), Apob, Ahr, Rock2, Slc26a4, Lpin1, Ucn(2nd), Hpcal1, Emilin1 Peak marker is not available/4 Bp211 D6Mit3, D6Mit12 75,049,528 - 136,649,683 715 total 443 known 102 novel 170 predicted Chga, Esr2, Yy1. Hif1a(2nd), Bdkrb1, Arg2, Dio2, Serpina1(2nd), Bdk rb2, Peak marker is not available/4 Bp182 D7Rat27, D7Rat139 32,004,505 - 89,945,095 351 total 2501 known 63 novel 38 predicted Nts, Cdk4,(2nd), Admr (2nd), Lrp1, No/1 Bp181 D7Rat73, D7Rat133 57,326,197 - 109,727,159 338 total 240 known 56 known 42 predicted Cdk4(2nd), Admr (2nd), Lrp1, Myc, Ndrg1. Tr hr, Angpt1, Gpr(2nd), No/1+1 Bp214 D7Rat135 (Peak) 88,021,524 - 128,021,684 490 total 399 known 47 novel 44 predicted Ndrg1, cyt11 b1, cyt11 b2, Ppara,, Pdgfb(2nd) Commd5, Mb, Adm2(2nd), Naga, Ptk2 No/ Bp183 D7Rat13, D7Rat80 124,315,013 - 141,301,748 248 total 206 known 27 novel 15 predicted Adm2(2nd), Acvrl1(2nd), Sp1, Vdr, Aqp2 Acvrl1 (1.43 M), Sp1 (0.14 M)/0 Copyright © 2012 SciRes. OPEN ACCESS 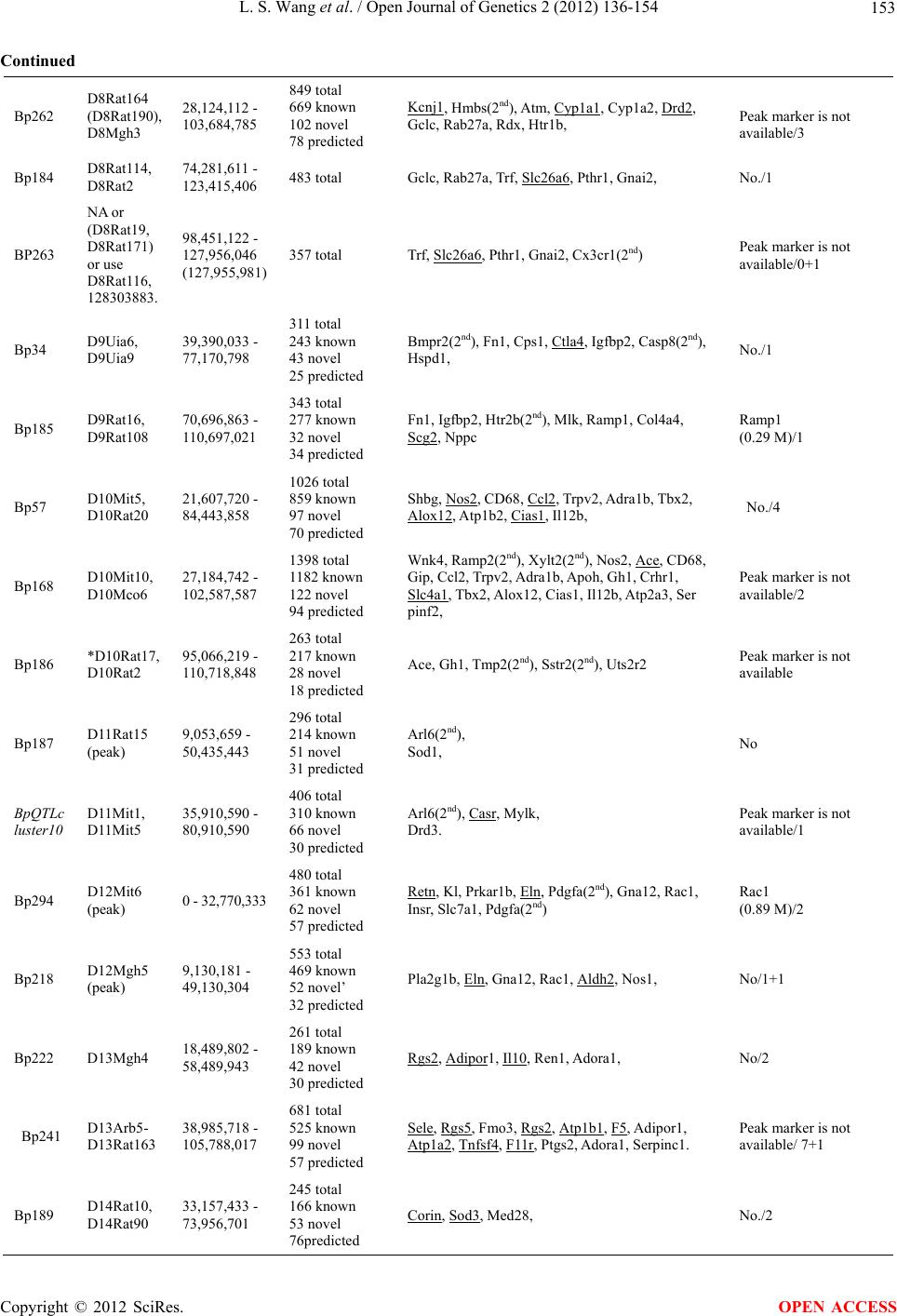 L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 153 Continued Bp262 D8Rat164 (D8Rat190), D8Mgh3 28,124,112 - 103,684,785 849 total 669 known 102 novel 78 predicted Kcnj1, Hmbs(2nd), Atm, Cyp1a1, Cyp1a2, Drd2, Gclc, Rab27a, Rdx, Htr1b, Peak marker is not available/3 Bp184 D8Rat114, D8Rat2 74,281,611 - 123,415,406 483 total Gclc, Rab27a, Trf, Slc26a6, Pthr1, Gnai2, No./1 BP263 NA or (D8Rat19, D8Rat171) or use D8Rat116, 128303883. 98,451,122 - 127,956,046 (127,955,981) 357 total Trf, Slc26a6, Pthr1, Gnai2, Cx3cr1(2nd) Peak marker is not available/0+1 Bp34 D9Uia6, D9Uia9 39,390,033 - 77,170,798 311 total 243 known 43 novel 25 predicted Bmpr2(2nd), Fn1, Cps1, Ctla4, Igfbp2, Casp8(2nd), Hspd1, No./1 Bp185 D9Rat16, D9Rat108 70,696,863 - 110,697,021 343 total 277 known 32 novel 34 predicted Fn1, Igfbp2, Htr2b(2nd), Mlk, Ramp1, Col4a4, Scg2, Nppc Ramp1 (0.29 M)/1 Bp57 D10Mit5, D10Rat20 21,607,720 - 84,443,858 1026 total 859 known 97 novel 70 predicted Shbg, Nos2, CD68, Ccl2, Trpv2, Adra1b, Tbx2, Alox12, Atp1b2, Cias1, Il12b, No./4 Bp168 D10Mit10, D10Mco6 27,184,742 - 102,587,587 1398 total 1182 known 122 novel 94 predicted Wnk4, Ramp2(2nd), Xylt2(2nd), Nos2, Ace, CD68, Gip, Ccl2, Trpv2, Adra1b, Apoh, Gh1, Crhr1, Slc4a1, Tbx2, Alox12, Cias1, Il12b, Atp2a3, Ser pinf2, Peak marker is not available/2 Bp186 *D10Rat17, D10Rat2 95,066,219 - 110,718,848 263 total 217 known 28 novel 18 predicted Ace, Gh1, Tmp2(2nd), Sstr2(2nd), Uts2r2 Peak marker is not available Bp187 D11Rat15 (peak) 9,053,659 - 50,435,443 296 total 214 known 51 novel 31 predicted Arl6(2nd), Sod1, No BpQTLc luster10 D11Mit1, D11Mit5 35,910,590 - 80,910,590 406 total 310 known 66 novel 30 predicted Arl6(2nd), Casr, Mylk, Drd3. Peak marker is not available/1 Bp294 D12Mit6 (peak) 0 - 32,770,333 480 total 361 known 62 novel 57 predicted Retn, Kl, Prkar1b, Eln, Pdgfa(2nd), Gna12, Rac1, Insr, Slc7a1, Pdgfa(2nd) Rac1 (0.89 M)/2 Bp218 D12Mgh5 (peak) 9,130,181 - 49,130,304 553 total 469 known 52 novel’ 32 predicted Pla2g1b, Eln, Gna12, Rac1, Aldh2, Nos1, No/1+1 Bp222 D13Mgh4 18,489,802 - 58,489,943 261 total 189 known 42 novel 30 predicted Rgs2, Adipor1, Il10, Ren1, Adora1, No/2 Bp241 D13Arb5- D13Rat163 38,985,718 - 105,788,017 681 total 525 known 99 novel 57 predicted Sele, Rgs5, Fmo3, Rgs2, Atp1b1, F5, Adipor1, Atp1a2, Tnfsf4, F11r, Ptgs2, Adora1, Serpinc1. Peak marker is not available/ 7+1 Bp189 D14Rat10, D14Rat90 33,157,433 - 73,956,701 245 total 166 known 53 novel 76predicted Corin, Sod3, Med28, No./2 Copyright © 2012 SciRes. OPEN ACCESS 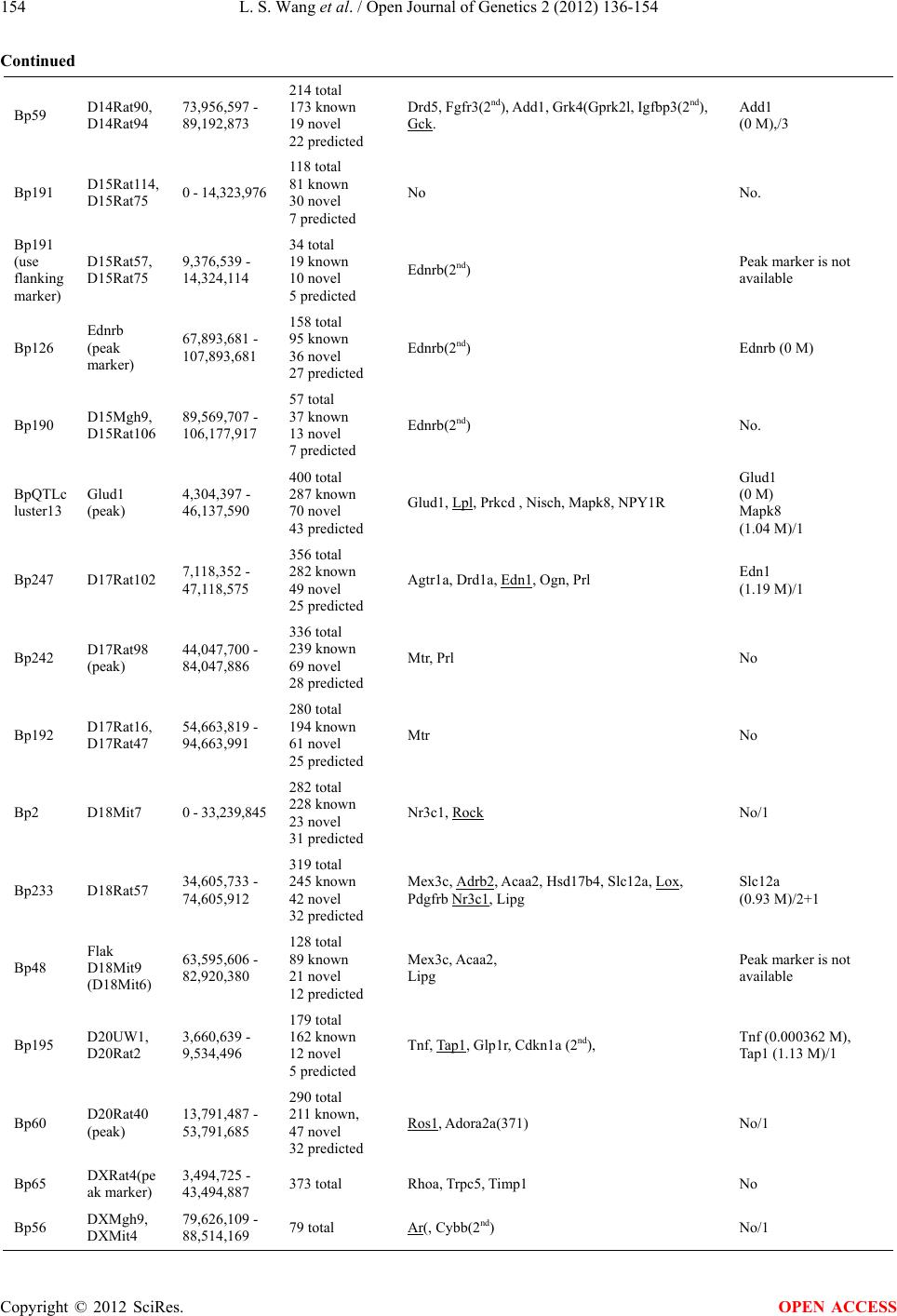 L. S. Wang et al. / Open Journal of Genetics 2 (2012) 136-154 Copyright © 2012 SciRes. 154 OPEN ACCESS Continued Bp59 D14Rat90, D14Rat94 73,956,597 - 89,192,873 214 total 173 known 19 novel 22 predicted Drd5, Fgfr3(2nd), Add1, Grk4(Gprk2l, Igfbp3(2nd), Gck. Add1 (0 M),/3 Bp191 D15Rat114, D15Rat75 0 - 14,323,976 118 total 81 known 30 novel 7 predicted No No. Bp191 (use flanking marker) D15Rat57, D15Rat75 9,376,539 - 14,324,114 34 total 19 known 10 novel 5 predicted Ednrb(2nd) Peak marker is not available Bp126 Ednrb (peak marker) 67,893,681 - 107,893,681 158 total 95 known 36 novel 27 predicted Ednrb(2nd) Ednrb (0 M) Bp190 D15Mgh9, D15Rat106 89,569,707 - 106,177,917 57 total 37 known 13 novel 7 predicted Ednrb(2nd) No. BpQTLc luster13 Glud1 (peak) 4,304,397 - 46,137,590 400 total 287 known 70 novel 43 predicted Glud1, Lpl, Prkcd , Nisch, Mapk8, NPY1R Glud1 (0 M) Mapk8 (1.04 M)/1 Bp247 D17Rat102 7,118,352 - 47,118,575 356 total 282 known 49 novel 25 predicted Agtr1a, Drd1a, Edn1, Ogn, Prl Edn1 (1.19 M)/1 Bp242 D17Rat98 (peak) 44,047,700 - 84,047,886 336 total 239 known 69 novel 28 predicted Mtr, Prl No Bp192 D17Rat16, D17Rat47 54,663,819 - 94,663,991 280 total 194 known 61 novel 25 predicted Mtr No Bp2 D18Mit7 0 - 33,239,845 282 total 228 known 23 novel 31 predicted Nr3c1, Rock No/1 Bp233 D18Rat57 34,605,733 - 74,605,912 319 total 245 known 42 novel 32 predicted Mex3c, Adrb2, Acaa2, Hsd17b4, Slc12a, Lox, Pdgfrb Nr3c1, Lipg Slc12a (0.93 M)/2+1 Bp48 Flak D18Mit9 (D18Mit6) 63,595,606 - 82,920,380 128 total 89 known 21 novel 12 predicted Mex3c, Acaa2, Lipg Peak marker is not available Bp195 D20UW1, D20Rat2 3,660,639 - 9,534,496 179 total 162 known 12 novel 5 predicted Tnf, Tap 1, Glp1r, Cdkn1a (2nd), Tnf (0.000362 M), Tap1 (1.13 M)/1 Bp60 D20Rat40 (peak) 13,791,487 - 53,791,685 290 total 211 known, 47 novel 32 predicted Ros1, Adora2a(371) No/1 Bp65 DXRat4(pe ak marker) 3,494,725 - 43,494,887 373 total Rhoa, Trpc5, Timp1 No Bp56 DXMgh9, DXMit4 79,626,109 - 88,514,169 79 total Ar(, Cybb(2nd) No/1
|