Open Journal of Genetics, 2012, 2, 163-166 OJGen
http://dx.doi.org/10.4236/ojgen.2012.23021 Published Online September 2012 (http://www.SciRP.org/journal/ojgen/)
Universal RNA editing in a human liver at the fetal st age
Dong Liu1, Cong Liu1, Xiyin Wang1, Sigurdur Ingvarsson2, Huiping Chen1
1Department of Medical Genetics, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, China
2Institute for Experimental Pathology and Faculty of Medicine, University of Iceland, Keldur, Iceland
Email: huiping@mail.hust.edu.cn
Received 14 May 2012; revised 17 June 2012; accepted 10 July 2012
ABSTRACT
It is known that RNA editing occurs in human cells,
which can change the information transmission from
DNA to RNA and proteins. Most previous studies
have focused on editing of the mRNAs. Here we re-
ported that several kinds of RNAs, including miRNA,
rRNA, mRNA, miscRNA and unknown RNA, exhib-
ited base editing in a human fetal liver. Several edit-
ing types are displayed. Our data reveals that RNA
editing may occur in different species of RNAs.
Keywords: RNA Editing; miRNA; rRNA; mRNA;
miscRNA; Fetal Liver
1. INTRODUCTION
The central dogma of molecular biology is that “DNA
makes RNA makes protein” or “RNA makes DNA makes
RNA makes protein”. The transmission of information is
from DNA to RNA to protein, or from RNA to DNA to
RNA to protein. The genetic information comes from the
DNA or RNA. However, the genetic information can be
changed during the transmission. Previous studies
showed that RNA can be modified or changed, known as
RNA editing [1,2]. In addition to mRNA editing, mi-
RNAs also exhibited editing [3-5]. The main editing type
is adenosine (A) to inosine (I) modification [6]. Because
inosine acts as guanosine during translation, A-to-I con-
version in coding sequences leads to amino acid changes
and often entails changes in protein function [3,6]. RNA
can also be edited by deaminating cytidine (C) to uridine
(U).
Although the editing of mRNA includes well defined
alteration of coding capacity, the editing of regulatory
RNA molecules is less understood, but might interfere
with their functionality. A recent understanding is that
there is a broader role of RNA molecules than previously
considered and that they are crucial contributors to cel-
lular processes such as gene expression on transcriptional
and post-transcriptional levels. Editing on those regula-
tory RNA molecules could modulate how effective they
are in cellular processes.
2. MATERIAL AND METHODS
We speculate that RNA modification or editing takes
place in all kinds of RNAs. To test this hypothesis, a
liver tissue was obtained from a female fetus of 27 weeks
delivered due to severe syndrome of high blood pressure
of the mother in Tongji Hospital, Wuhan, Hubei, China.
The fetus died shortly after delivery. The mother of the
fetus had no other diseases than high blood pressure.
Small RNA (≤200 nt) was isolated from about 50 mg of
the liver tissue using a mirVanaTM miRNA isolation kit
(Ambion, Austin, TX) following the manufacturer’s in-
structions.
About 1 μg of the isolated RNAs were polyadenylated
using poly(A) polymerase (New England Biolabs). Then
the Poly(A)-tailed small RNA was purified through
phenol/chloroform extraction and ethanol precipitation.
A 5’ linker (5’-GGA CAC UGA CAU GGA CUG AAG
GAG UAG AAA-3’) was ligated to the poly(A)-tailed
RNA using T4 RNA ligase (New England Biolabs) and
the ligation products were recovered by phenol/ chloro-
form extraction followed by ethanol precipitation. Re-
verse transcription was conducted using the entire
poly(A)-tailed RNA with primer (5’-CGC TAC GTA
ACG GCA TGA CAG TG (T)24-3’) and SuperScript III
reverse transcriptase (Invitrogen) according to the manu-
facturer’s instructions. The cDNA was PCR amplified
using primers 5’-GGA CAC TGA CAT GGA CTG AAG
GAG TA-3’ and 5’-CGC TAC GTA ACG GCA TGA
CAG TG-3’. After agarose gel electrophoresis and Gold
View staining slices with PCR products were excised
and purified, using EasyPure quick gel extraction kit
(TransGen Biotech). The DNA fragments were directly
cloned into pEASY-T1 vector (TransGen Biotech). Col-
ony PCR was performed and the clones with PCR prod-
ucts were sequenced bidirectionally to avoid sequen-
cing errors. The small RNA sequences were analyzed
using BLAST analysis against the human genome and
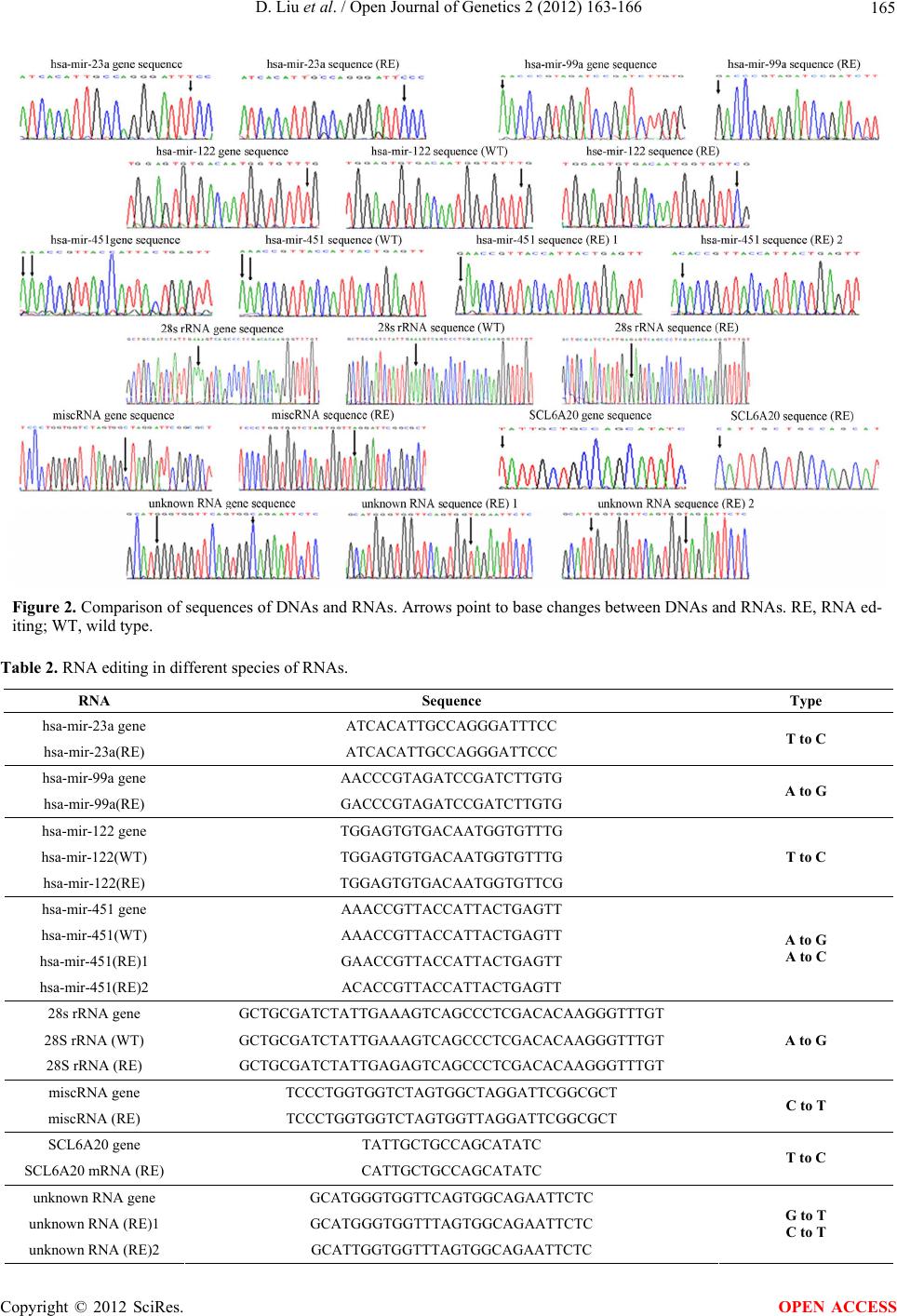
transcripts databases and miRBase (Figure 1).
OPEN ACCESS