Journal of Cancer Therapy
Vol.2 No.5(2011), Article ID:16670,8 pages DOI:10.4236/jct.2011.25101
Antitumor Activity of Disintegrin-Like Components from the Venom of Montivipera raddei
![]()
Department of Human and Animal Physiology, Yerevan State University, Yerevan, Armenia.
E-mail: samiryan@fulbrightmail.org
Received August 30th, 2011; revised September 30th, 2011; accepted November 10th, 2011.
Keywords: Snake venom, Montivipera raddei (Armenian viper), disintegrins, antitumor activity
ABSTRACT
Our findings represent the first report of the antitumor activity of the disintegrin-like components from the venom of Armenian viper (M. raddei). The venom of M. raddei was separated by reverse phase high-performance liquid chromatography (RP HPLC), and individual fractions were analyzed for disintegrin activity. Disintegrin-like components from the venom of M. raddei, by blocking integrins on breast cancer cells (MDA-MB-435), not only interferes with adhesion of breast cancer cells to the extracellular matrix, but also inhibits cellular mobility which is essential for cancer invasion. These effects seriously curtail the metastatic capability of the MDA-MB-435 cells.
1. Introduction
Cancer is a class of diseases or disorders characterized by uncontrolled division of cells and the ability of these to spread, either by direct growth into adjacent tissue through invasion, or by implantation into distant sites by metastasis (where cancer cells are transported through the bloodstream or lymphatic system). Cell division or cell proliferation is a physiological process that occurs in almost all tissues and under many circumstances. Normally the balance between proliferation and programmed cell death is tightly regulated to ensure the integrity of organs and tissues. The uncontrolled and often rapid proliferation of cells can lead to an initially benign tumor but it may carry a risk of turning malignant (cancer). Benign tumors do not spread to other parts of the body or invade other tissues, and they are rarely a threat to life unless they extrinsically compress vital structures. Malignant tumors can invade other organs, spread to distant locations (metastasize) and become life-threatening [1].
Cancer cells can also switch the types of extracellular matrix receptors (integrins) they express, favoring ones that transmit progrowth signals [2]. These bifunctional, heterodimeric cell surface receptors physically link cells to extracellular superstructures known as the extracellular matrix (ECM). Successful binding to specific moieties of the ECM enables the integrin receptors to transduce signals into the cytoplasm that influence cell behavior, ranging from quiescence in normal tissue to motility, resistance to apoptosis, and entrance into the active cell cycle. Conversely, the failure of integrins to forge these extracellular links can impair cell motility, induce apoptosis, or cause cell cycle arrest [2].
Integrin subunits penetrate the plasma membrane, and in general have very short cytoplasm domains of about 40 - 70 amino acids. Outside the cell plasma membrane, the alpha and beta chains of integrins lie close together along a length of about 23 nm, the final 5 nm NH2-termini of each chain form a ligand-binding region for the ECM [3].
The importance of vitronectin receptors (integrins αvβ3 and αvβ5) in angiogenesis is well known [4]. Yet integrins are not only central to angiogenesis. Pathological examination indicates that the level of αv expression in breast cancer is directly related to migration or metastasis [5], with higher levels of integrin associated with a more metastatic phenotype.
The main functions of integrins are: attachment of the cell to the ECM, and signal transduction from the ECM to the cell. One of their most important functions is their role in cell migration. Three steps: cell adhesion, cell migration and cell invasion are critical to tumor metastasis and cancer progression. Identifying a molecule that will disrupt these functions is the beginning to a path that may lead to development of therapeutic agents for cancer.
Several classes of integrins recognize the RGD sequence present in ECM proteins [6]. Previous studies showed that peptides containing the RGD sequence bind competitively to integrins on the surface of tumor cells and inhibit binding of the cancer cells to the ECM [7]. Agents that disrupt interactions of these integrins should have significant anti-tumor activity [8].
Crude snake venoms are complex mixtures of components with diverse pharmacological activity and toxicity. The venoms of snakes belonging to one defined family contain a regularly recurrent set of major and minor components. However, it has been shown in numerous reports that the amount of individual components varies in venoms of different provenance. The term “disintegrin” was used to describe a group of viper venomderived, nonenzymatic small proteins that shared numerous structural and functional properties [9]. Disintegrins were discovered and isolated from the venom of snakes and were given their name because of their biological function of binding to integrins.
Disintegrins contain a characteristic tripeptide motif, e.g. Arg-Gly-Asp (RGD), which is critical for binding integrins. Disintegrins are among the most potent known natural inhibitors of integrin function and are active at nanomolar concentrations, whereas the activity of the short linear RGD peptides is observed at the micromolar level. Disintegrins have been found to inhibit platelet aggregation, angiogenesis, metastasis and tumor growth [10,11]. As such, disintegrins can be explored as therapeutic agents against a number of pathologies including Alzheimer’s disease, inflammation, autoimmune diseases, virus infection, asthma, osteoporosis, thrombosis and cancer [12-14]. All disintegrins contain an R/KGD (Arg/ Lys-Gly-Asp) sequence in the carboxyl-terminal half of the molecule that is essential to their ability to obstruct integrin interaction with the ECM [11,15].
Disintegrins from different snake species have been characterised: albolabrin, applagin, barbourin, batroxostatin, bitistatin, obtustatin [16], schistatin [17], echistatin [18], elegantin, eristicophin, flavoridin [19], halysin, kistrin, tergeminin, salmosin [20] and triflavin.
Snake venom RGD-disintegrins have been the target for investigation of αvβ3 integrin as a potential target for the suppression of cancer. These disintegrins are useful tools to decipher the mechanisms that occur during αvβ3- dependent angiogenesis. Monomeric RGD-disintegrins, accutin [21], triflavin [22], salmosin [23], rhodostomin [24], and the homodimeric RGD-disintegrin, contortrostatin [25], inhibit angiogenesis by binding to endothelial cells via αvβ3. It has been suggested that the binding of disintegrins to endothelial cells inhibits their motility and proliferation, possibly by induction of apoptosis in endothelial cells. Snake venom RGD-disintegrins showed direct interaction in several tumor cell lines. The blocking of αvβ3 integrin in tumor cells inhibited their adhesion to the extracellular matrix and significantly reduced their motility, subsequently inhibiting metastasis. This effect was noted for monomeric medium-sized disintegrins, such as salmosin [23] and triflavin [22] and the homodimeric disintegrin, contortrostatin [26]. These drugs may be considered as alternatives to the humanized monoclonal antibodies used in cancer therapy.
Armenian viper [Montivipera raddei (Boettger, 1890)] belongs to the family Viperidae and is widely distributed in Armenia, at an altitude of 1500 - 1600 m above the sea level (Arai-ler mountain). The venom of M. raddei was separated by reverse phase HPLC, and individual fractions were analyzed for disintegrin activity.
2. Materials and Methods
2.1. Materials
Venom of Armenian viper [Montivipera raddei (Boettger, 1890)] was received from the Institute of Zoology of Armenian National Academy of Sciencies. Purification was done by multi-step high-performance liquid chromatography (HPLC) according to an established protocol [7].
2.2. Methods
MDA-MB-435 cell line. Human metastatic mammary cancer cell line MDA-MB-435 was provided by Dr. Janet E. Price (Monroe Dunaway Anderson Cancer Center; Houston, Texas). Dulbecco’s Modified Eagle Medium (DMEM) with 10% fetal bovine serum was employed for culture of MDA-MB-435 cells. Cells were incubated at 37˚C with 5% CO2.
Isolation of proteins. We performed chromatographic division of Montivipera raddei (M. raddei) venom twice. In the first experiment we put the solution containing 50 mg of the venom through the column. The liquid eluted at 1 ml/min. The emergence of the first peak indicated that the venom has completely entered the column. Starting from the 30th minute, we picked 1 ml of eluate each time and accordingly marked the samples from 30 to 80.
Upon performance of SDS PAGE electrophoresis we determined that samples 42, 46, 50 and 53 most probably contained low-molecular weight fractions (up to 15 kDa).
In the subsequent experiment 200 mg of the same venom was added to the solution for chromatography through HPLC. Chromatograms received in this experiment are shown in figure 1. Since this chromatogram was identical with the first one we included only that one in the article. For the second experiment we selected eluates in a larger quantity (5 - 10 ml/min), so the numbering is already presented as samples 18 - 26. We selected samples 18 and 19 as the most probable ones to contain disintegrin fractions. For RP HPLC separations, M. raddei crude venom was dissolved in 750 μL of distill water. Proteins in the soluble material were separated using an ETTANTMLC HPLC system (Amersham Biosciences, Sweden) and a Lichrospher RP100 C18 column (250 × 4 mm, 5 mm particle size) eluted with a linear gradient of 0.1% triftoracetat in water (solution A) and acetonitril (solution B) using the following chromatographic conditions: isocratically (5% solution B) for 20 min, followed by 5% - 55% solution B for 68 min, and 55% - 70% solution B for 20 min. Protein detection was at 215 nm and peaks were collected manually and dried in a SpeedVac (Amersham Biosciences, Savant). The purity of the RP isolated proteins was initially checked by SDS-PAGE.
SDS-PAGE electrophoresis. Samples were mixed with SDS-loading buffer (15 μL/15 μL). Electrophoresis was carried out in 12.5% polyacrylamide gel at pH 6.4 in Phast System (Pharmacia, Uppsala, Sweden) according to manufacturer’s instructions. Following electrophoresis, the gel was stained with Coomassie Brilliant Blue R 250.
Western Blot. The proteins of the samples were separated using SDS-PAGE electrophoresis. Then proteins were transferred onto the membrane of polyvinylidene difluoride (PVDF) by electroblotting. Membrane was washed and placed into the blocking solution (0.5 g BSA in 50 ml TTBS) for one hour, washed again (with TTBS, three times by 15 min), and placed in the primary antibody solution (10 μl antibodies in 100 ml TTBS), and incubated overnight. After rinsing the membrane to remove unbound primary antibody, the membrane was exposed to secondary antibody (Goat anti-rabbit IgG-HRP, 5 μl antibodies in 50 ml TTBS), and placed on the sensitive sheet of photographic film, and exposed to the light to receive an image of the antibodies bound to the blot.
Adhesion Assay. 10 μL of vitronectin stock solution (0.5 μg/μL) was diluted in 6 ml PBS. Wells of 24-well flat bottom plates (Immulon II; Dynex Technologies Inc., Chantilly, VA, USA) were coated with vitronectin (150 μL/well) overnight at 37˚C. For the adhesion assay, MDA-MB-435 cells (5 × 104 cells per well) were seeded in the coated microtiter plate wells, and allowed to adhere to vitronectin overnight at 37˚C. For the inhibition assay, MDA-MB-435 cells were treated with samples 42, 46, 50, and 53 of the M. raddei venom (5 μg/300 μL per well) collected during HPLC and allowed to adhere to vitronectin overnight at 37˚C. For the assay, the wells were washed with PBS to remove unbound cells and cell debris.
Scheme of making pictures of the Invasion Assay
After incubation cells in each well were separately washed (by 200 μL PBS), trypsinised (100 μL), centrifuged (1000 g for 10 min), and resuspended by 200 μL of Media. Cell densities were determined with a Coulter counter. Assays at each sample were performed twice in triplicate.
Wound Assay. Tissue wounds undergo a complex and ordered series of events to repair tissue. These events may include infiltration of inflammatory immune cells as part of the process to remove and destroy necrotic tissue, increased vascularization by angiogenic factors, and increased cell proliferation and extracellular matrix deposition. Wound assay is carried out in cell or tissue culture for many years to estimate the migration and proliferation rates of different cells and culture conditions. This assay generally involves first growing a confluent cell monolayer. A small area is then disrupted and a group of cell destroyed or displaced by scratching a line through the layer. The open gap is then inspected microscopically over time as the cells move in and fill the damaged area.
Scheme of the Migration Assay
Twelve-well flat bottom plates were coated with matrigel (250 μL), and incubated overnight at 37˚C. MDAMB-435 cells (10 × 104 cells in 500 μL of media per well) were seeded on coated wells, and allowed to adhere to matrigel overnight at 37˚C. A line with a marker with bottom of the dishes was drawn.
Using a sterile 200 μL pipette tip three separate wounds trough the cells moving perpendicular to the line drawn in the step above were scratched. Base media with different samples of the venom of M. raddei collected during HPLC analysis was prepared. Cells were washed by PBS (300 μL) replaced with 500 μL media containing 0.25 μg/μl samples’ (each in duplicate) solutions. Pictures of control (after the first washing), 6 hours and 24 hours treatment using phase contrast and 10× were taken just above and just below each line. After each measurement the old media was replaced with the fresh-prepared media with the same samples.
MDA-MB-435 cell invasion assay. Modified Boyden chambers were employed for this assay [27]. 12-well Transwell chambers (Corning Costar, Cambridge, MA) with 8.0 μm pores were coated with 25 μl of 1:50 diluted Matrigel in serum free medium, and allowed to air dry in a sterile hood overnight and rehydrated with serum free medium for 2 h at room temperature prior to use. Media was removed from the inserts. 300 μL of the conditioned medium collected from fibrosarcoma cell (HT1080) culture was added to the lower chambers as chemoattractant. MDA-MB-435 cells were mixed with different samples of purified disintegrin-like components of the M. raddei venom in serum free medium. 300 μL of the cells (1.5 × 105) suspension was then applied to the upper chambers of the Transwell, and cells were allowed to invade across the Matrigel-coated membrane for 8 h. After noninvaded cells were removed with wet cotton swabs, invaded cells attached to the bottom of the membrane were fixed and stained with the Crystal Violet for 10 min and washed by PBS four times. Pictures of Transwell membranes with invaded cells from control (without treatment) and experimental chambers were done immediately after washing. Treatments were performed in duplicate, and the assay was repeated to verify results.
3. Results and Discussion
Before each experiment we determined the protein concentration through BCA-analysis and used several different concentrations that we were interested in for our experiment. Separate fractions received during the HPLC analysis (Figure 1) was initially analyzed by SDS-PAGE. SDS-PAGE electrophoresis revealed that the selected fractions contained two bands at 10 kDa, and 15 kDa corresponding to disintegrins molecular weights, and few bands at 25 - 30 kDa, plus several minor bands (Figures 2 and 3).
Samples 18 and 19 were tested with Western blot analysis. As primary and secondary antigenes we used polyclonal antibodies to the disintegrins from the blood of animals with cancer. Data received showed that fractions 18 and 19 form two binding sites with antibodies (Figure 4). This proved the presence of disintegrins in the researched fractions, which testify the immunoreactivity of disintegrins from the venom of M. raddei.

Figure 1. Chromatograms of the RP HPLC of the Montivipera raddei crude venom. Chromatographical conditions: isocratically (5% B) 20 min, 5% - 55% B - 68 min, 55% - 70% B - 20 min.
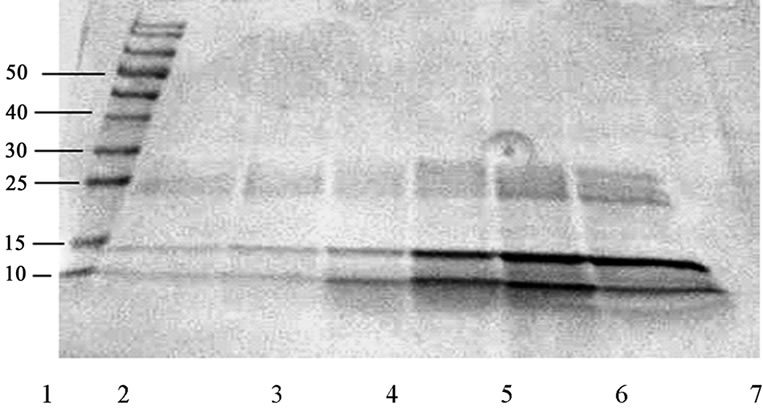
Figure 2. SDS-PAGE of the samples 36 (2), 39 (3), 42 (4), 46 (5), 50 (6), 53 (7) of the M. raddei venom.
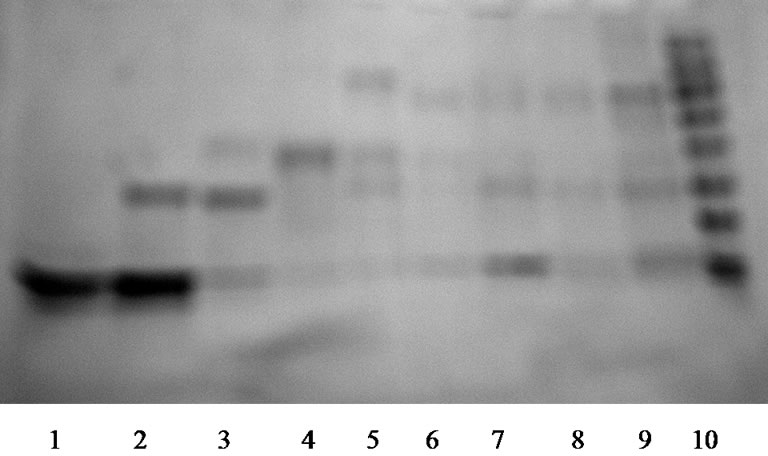
Figure 3. SDS-PAGE of the samples 18 (1), 19 (2), 20 (3), 21 (4), 22 (5), 23 (6), 24 (7), 25 (8), 26 (9) of the M. raddei venom.

Figure 4. Western blot analysis of samples 18 and 19 of the M. raddei venom. On the right—protein ladder.
Previously it was shown that human breast carcinoma (MDA-MB-435) cells adhered to various extracellular matrix proteins, including fibronectin, vitronectin, type I collagen, and laminin [26]. It was of interest to test whether adhision of these tumor cells to vitronectin may be inhibited by desintegrin-like fractions from the venom of V. raddei. The most inhibition of MDA-MB 435 cells adhision to vitronectin was revealed after pretreatment by the fraction 53 (98.65%), and 50 (89.11%) which molecular weights are less than 15 kDa (see Table 1).
It was shown that the migration of cells was inhibited by all fractions (including disintegrin contortrostatin from the venom of Agkistrodon contortrix contortrix, as a positive control) with known molecular weight 13,5 kDa) after 6 hours of pretreatment relatively to the cells without treatment (figures 5-6). And after 24 hours the most pronounced inhibition was revealed for the fraction 46, and also fractions 18 and 19 in different concentrations.

Table 1. Inhibition of the adhesion of MDA-MB 435 cells to vitronectin by proteins isolated from the venom of M. raddei.
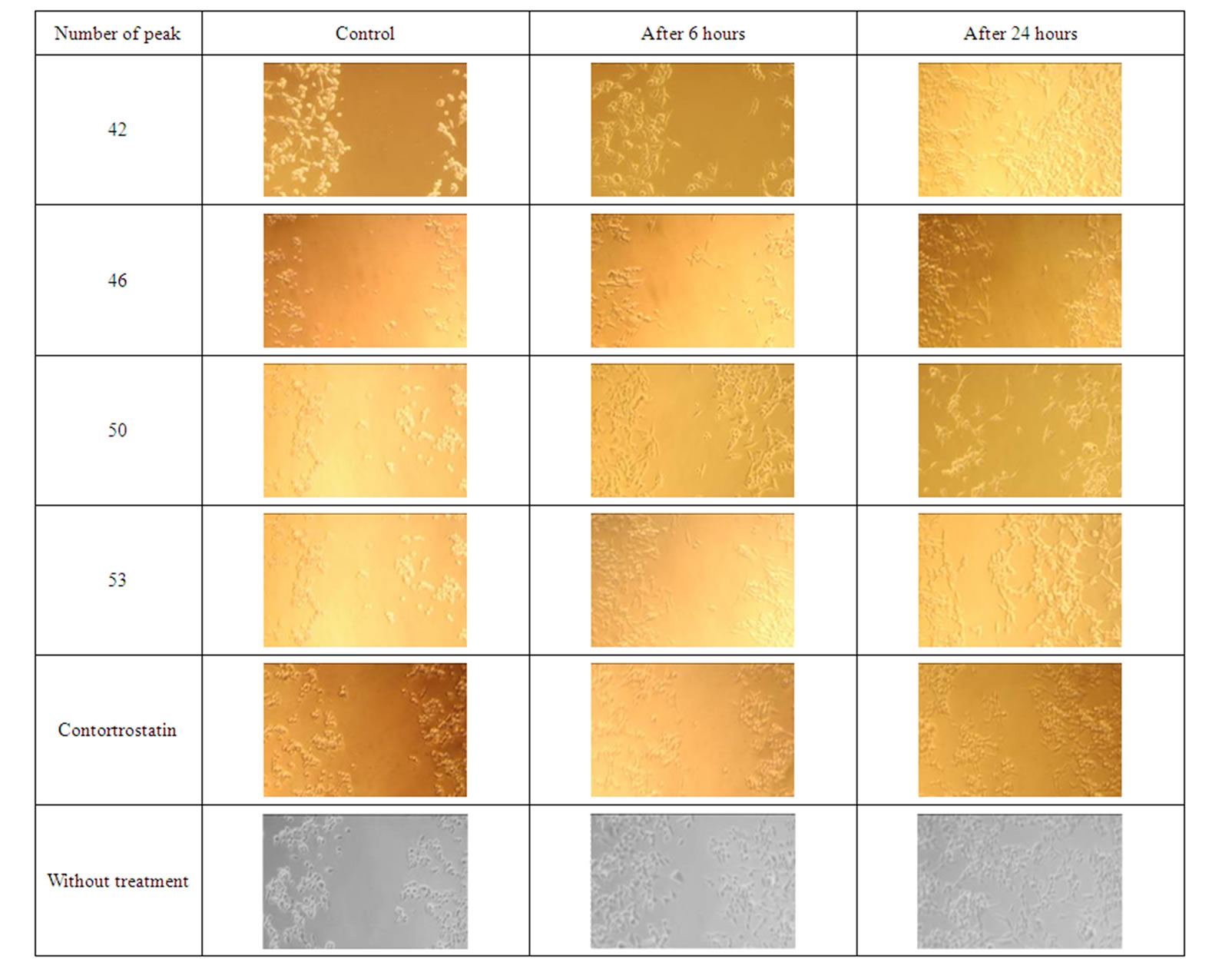
Figure 5. Inhibition of migration of MDA-MB 435 breast cancers cells through the matrigel by proteins isolated from the venom of M. raddei.

Figure 6. Inhibition of migration of MDA-MB 435 breast cancers cells through the matrigel by proteins isolated from the venom of M. raddei.
In order to test whether disintegrin-like fractions from the venom of M. raddei can inhibit invasion for MDAMB 435 cells across Matrigel™-coated Transwells, the cells were treated by different samples and incubated for 8 hours (37˚C). The pictures made after 8-hour incubation showed that the maximal inhibition was exhibited after the treatment by the samples 46 and 50 in compareson with cells without treatment and cells treated by contortrostatin (Figure 7).
Disintegrins are potent inhibitors of integrin-ligand interactions. The integrin inhibitory profile of disintegrins primarily depends on the sequence of a tripeptide RGD located at the apex of a mobile loop and constrained in its active conformation by the appropriate pairing of disulfide bonds.
At diagnosis, over 60% of breast cancer patients have metastatic disease. Dissemination of the cancer cells from the primary tumor to remote tissues or organs, and progressive growth of the metastatic cells at the distant site, is the most common cause of death in breast cancer patients.
Therefore, control of metastasis offers an important avenue for breast cancer treatment. Local invasion and penetration of the cancer cells into the lumen of a blood vessel (intravasation) will allow tumor cells to disseminate.
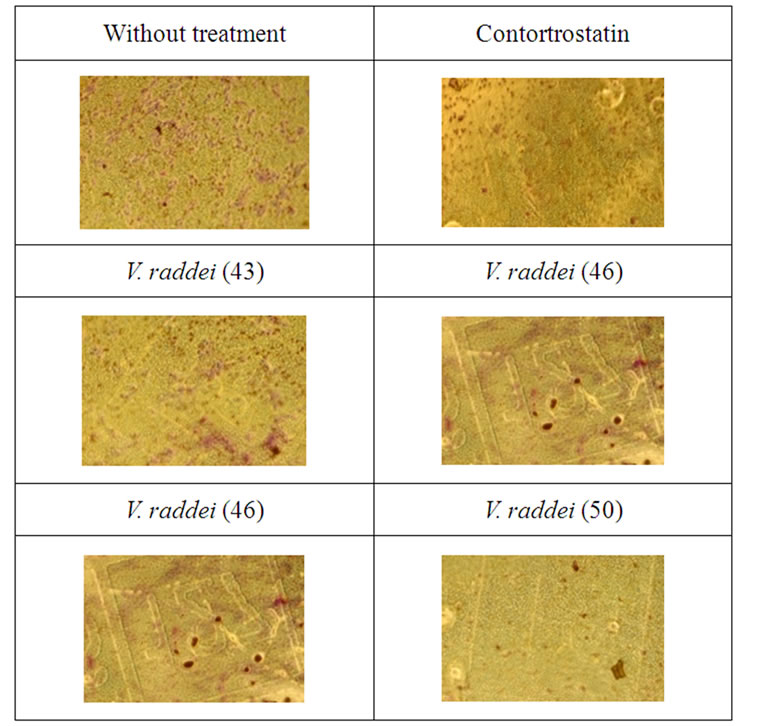
Figure 7. Invasion of MDA-MB 435 cells pretreated with fractions 43, 46, and 50 from the venom of M. raddei across Matrigel™-coated Transwells.
Other important events are the formation of tumor cell microthrombi, adherence, penetration through the endothelium and the adjacent basement membrane (extravasation), and ability to grow in a foreign environment and to stimulate angiogenesis. These processes must be successfully accomplished by the cancer cells to develop into metastatic tumors.
In view of the complexity of this process, a combination of factors may be required to effectively arrest metastasis. The importance of integrins in tumor metastasis is now becoming apparent [28]. Integrins mediate cell adhesion, migration, matrix degradation, proliferation, gene regulation, and angiogenesis.
Disintegrin-like component isolated from the venom of M. raddei has an effect on multiple steps crucial to tumor metastasis. Currently we are working on isolating the disintegrins from the venom of M. raddei in its pure form which will hopefully be given a corresponding name (like lebestatin from Vipera lebetina venom, contortrostatin from Agkistrodon contortrix contortrix venom, echistatin from Echis carinatus venom, etc).
Our data indicate that disintegrin-like component (sample 53) from the venom of M. raddei, with molecular weight less than 15 kDa has inhibitory effect on adhesion of MDA-MB-435 cells to vitronectin. We have also shown that this disintegrin-like component affects MDAMB-435 cell invasive capacity, as evidenced by its ability to reduce invasion (samples 46 and 50) through a Matrigel membrane.
We conclude that disintegrin-like components from the venom of M. raddei, by blocking integrins on breast cancer cells, not only interferes with adhesion of breast cancer cells to the extracellular matrix, but also inhibits cellular mobility which is essential for cancer invasion. These effects seriously curtail the metastatic capability of the MDA-MB-435 cells.
Our findings represent the first report of the antitumor activity of the disintegrin-like components from the venom of M. raddei. Further research on the molecular structure and mechanism of action of disintegrin-like components from the venom of M. raddei, in our opinion, should establish a useful therapeutic approach to impede breast cancer progression.
4. Acknowledgements
My sincere acknowledgements to
1) Francis Markland and Stephen Swenson for help and facilities to carry out experiments in the Cancer Research Laboratory (University of Southern California, Department of Biochemistry).
2) Aram Aghasyan (Institute of Zoology, Armenian National Academy of Sciences) for the Macrovipera raddei venom.
3) This work was supported by Fulbright Scholar Program Advanced Research and University Lecturing Awards in the United States.
REFERENCES
- D. Hanahan and R. A. Weinberg, “The Hallmarks of Cancer,” Cell, Vol. 100, No. 1, 2000, pp. 57-70. doi:10.1016/S0092-8674(00)81683-9
- F. G. Giancotti and E. Ruoslahti, “Integrin Signaling,” Science, Vol. 285, No. 5430, 1999, pp. 1028-1032. doi:10.1126/science.285.5430.1028
- H. Lodish, A. Berk, L. S. Zipursky, P. Matsudaira, D. Baltimore and J. Darnell, “Molecular Cell Biology,” 5th Edition, Freeman, New York, 2003, p. s973.
- P. C. Brooks, S. Stromblad, R. Klemke, D. Visscher, F. H. Sarkar and D. A. Cheresh, “Antiintegrin Alpha v Beta 3 Blocks Human Breast Cancer Growth and Angiogenesis in Human Skin,” Journal of Clinical Investigation, Vol. 96, No. 4, 1995, pp. 1815-1822. doi:10.1172/JCI118227
- B. Felding-Habermann, T. E. O’Toole, J. W. Smith, E. Fransvea, Z. M. Ruggeri, M. H. Ginsberg, P. E. Hughes, N. Pampori, S. J. Shattil, A. Saven and B. M. Mueller, “Integrin Activation Controls Metastasis in Human Breast Cancer,” Proceedings of the National Academy of Sciences of the USA, Vol. 98, No. 4, 2001, pp. 1853-1858. doi:10.1073/pnas.98.4.1853
- R. Hynes, “Integrins: Versatility Modulation, and Signaling in Cell Adhesion,” Cell, Vol. 69, No. 1, 1992, pp. 11- 25. doi:10.1016/0092-8674(92)90115-S
- M. Trikha, Y. A. De Clerk and F. S. Markland, “Contortrostatin, a Snake Venom Disintegrin, Inhibits β1 Integrin-Mediated Human Metastatic Melanoma Cell Adhesion, and Blocks Experimental Metastasis,” Cancer Research, Vol. 54, No. 18, 1994, pp. 4993-4998.
- S. Swenson, F. Costa, W. Ernst, G. Fujii and F. S. Markland, “Contortrostatin, a Snake Venom Disintegrin with Anti-Angiogenic and Anti-Tumor Activity,” Pathophysiol Haemost Thromb, Vol. 34, No. 4-5, 2005, pp. 169-176. doi:10.1159/000092418
- M. A. McLane, E. E. Sanchez, A. Wong, C. PaquetteStraub and J. C. Perez, “Disintegrins,” Current Drug Targets: Cardiovascular and Hematological Disorders, Vol. 4, No. 4, 2004, pp. 327-355. doi:10.2174/1568006043335880
- F. S. Markland, “Snake Venoms and the Hemostatic System,” Toxicon, Vol. 36, No. 12, 1998, pp. 1749-1800. doi:10.1016/S0041-0101(98)00126-3
- M. A. McLane, C. Marcinkiewicz, S. VijayKumar, I. Wierzbicka-Patynowski and S. Niewiarowski, “Viper Venom Disintegrins and Related Molecules,” Proceedings of the Society for Experimental Biology and Medicine, Vol. 219, No. 2, 1998, pp. 109-119.
- C. Marcinkiewicz, “Functional Characteristic of Snake Venom Disintegrins: Potential Therapeutic Implication,” Current Pharmaceutical Design, Vol. 11, No. 7, 2005, pp. 815-827. doi:10.2174/1381612053381765
- N. Moiseeva, R. Bau, S. D. Swenson, F. S. Markland, J.-Y. Choe, Z.-J. Liu and M. Allaire, “Structure of Acostatin, a Dimeric Disintegrin from Southern Copperhead (Agkistrodon cotortrix contortrix), at 1.7 Å Resolution,” Acta Crystallographica Section D: Biological Crystallography, Vol. 64, 2008, pp. 466-470. doi:10.1107/S0907444908002370
- C. S. Xu and S. Rahman, “Identification by Site-Directed Mutagenesis of Amino Acid Residues Flanking RGD Motifs of Snake Venom Disintegrins for Their Structure and Function,” Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao (Shanghai), Vol. 33, No. 2, 2001, pp. 153-157.
- R. J. Gould, M. A. Polokoff, P. A. Friedman, T-F. Huang, J. C. Holt, J. J. Cook and S. Niewiarowski, “Disintegrins: A Family of Integrin Inhibitory Proteins from Viper Venoms,” Proceedings of the Society for Experimental Biology and Medicine, Vol. 195, No. 2, 1990, pp. 168-171.
- M. P. Moreno-Murciano, D. Monleón, C. Marcinkiewicz, J. J. Calvete and B. Celda, “NMR Solution Structure of the Non-RGD Disintegrin Obtustatin,” Journal of Molecular Biology, Vol. 329, No. 1, 2003, pp. 135-145. doi:10.1016/S0022-2836(03)00371-1
- S. Bilgrami, S. Yadav, P. Kaur, S. Sharma, M. Perbandt, C. Betzel and T. P. Singh, “Crystal Structure of the Disintegrin Heterodimer from Saw-Scaled Viper (Echis carinatus) at 1.9 A Resolution,” Biochemistry, Vol. 44, No. 33, 2005, pp. 11058-11066. doi:10.1021/bi050849y
- D. Monleón, V. Esteve, H. Kovacs, J. J. Calvete and B. Celda, “Conformation and Concerted Dynamics of the Integrin-Binding Site and the C-terminal Region of Echistatin Revealed by Homonuclear NMR,” Biochemical Journal, Vol. 387, No. 1, 2005, pp. 57-66. doi:10.1042/BJ20041343
- Y. Fujii, D. Okuda, Z. Fujimoto, K. Horii, T. Morita and H. Mizuno, “Crystal Structure of Trimestatin, a Disintegrin Containing a Cell Adhesion Recognition Motif RGD,” Journal of Molecular Biology, Vol. 332, No. 5, 2003, pp. 1115-1122. doi:10.1016/S0022-2836(03)00991-4
- J. Shin, S. Y. Hong, K. Chung, I. Kang, Y. Jang, D. S. Kim and W. Lee, “Solution Structure of a Novel Disintegrin, Salmosin, from Agkistrondon halys Venom,” Biochemistry, Vol. 42, No. 49, 2003, pp. 14408-14415. doi:10.1021/bi0300276
- C. H. Yeh, H. C. Peng and T. F. Huang, “Accutin, a New Disintegrin Inhibits Angiogenesis in Vitro and in Vivo by Acting as Integrin Alphabeta3 Antagonist and Inducing Apoptosis,” Blood, Vol. 92, No. 9, 1998, pp. 3268-3276.
- J. R. Sheu, M. H. Yen, Y. C. Kan, W. C. Hung, P. T. Chang and H. N. Luk, “Inhibition of Angiogenesis in Vitro and in Vivo: Comparison of the Relative Activities of Triflavin, an Arg-Gly-Asp-Containing Peptide and AntiAlpha(v)beta3 Integrin Monoclonal Antibody,” Biochimica et Biophysica Acta, Vol. 1336, No. 3, 1997, pp. 445- 454. doi:10.1016/S0304-4165(97)00057-3
- S. I. Kim, K. S. Kim, H. S. Kim, M. M. Choi, D. S. Kim and K. H. Chung, “Inhibition of Angiogenesis by Salmosin Expressed in Vitro,” Oncology Research, Vol. 14, No. 4-5, 2004, pp. 227-233.
- C. H. Yeh, H. C. Peng, R. S. Yang and T. F. Huang, “Rhodostomin, a Snake Venom Disintegrin, Inhibits Angiogenesis Elicited by Basic Fibroblast Growth Factor and Suppresses Tumor Growth by a Selective Alpha(v)beta(3) Blockade of Endothelial Cells,” Molecular Pharmacology, Vol. 59, No. 5, 2001, pp. 1333-1342.
- F. S. Markland, K. Shieh, Q. Zhou, V. Golubkov, R. P. Sherwin and V. Richters, “A Novel Snake Venom Disintegrin that Inhibits Human Ovarian Cancer Dissemination and Angiogenesis in an Orthotopic Nude Mouse Model,” Haemostasis, Vol. 31, No. 3-6, 2001, pp. 183-191.
- Q. Zhou, R. P. Sherwin, C. Parrish, V. Richters, S. G. Groshen, D. Tsao-Wei and F. S. Markland, “Contortrostatin, a Dimeric Disintegrin from Agkistrodon contortrix, Inhibits Breast Cancer Progression,” Breast Cancer Research and Treatment, Vol. 61, No. 3, 2000, pp. 249-260. doi:10.1023/A:1006457903545
- L. A. Repesh, “A New in Vitro Assay for Quantitating Tumor Cell Invasion,” Invasion Metastasis, Vol. 9, No. 3, 1989, pp. 192-208.
- F. G. Giancotti and F. Mainiero, “Integrin-Mediated Adhesion and Signaling in Tumorgenesis,” Biochimica et Biophysica Acta, Vol. 1198, No. 1, 1994, pp. 47-64.