American Journal of Plant Sciences
Vol.5 No.5(2014), Article ID:43916,10 pages DOI:10.4236/ajps.2014.55087
Induced Accumulation of Polyphenolics and Flavonoids in Cyanobacteria under Salt Stress Protects Organisms through Enhanced Antioxidant Activity
Dhananjaya P. Singh1*, Ratna Prabha1, Kamlesh K. Meena2, Lalan Sharma1, Arun K. Sharma1
1National Bureau of Agriculturally Important Microorganisms, Indian Council of Agricultural Research, Kushmaur, Maunath Bhanjan, 275103, India
2National Institute of Abiotic Stress Management, Indian Council of Agricultural Research, Baramati, Pune, 413115, India
Email: *dpsfarm@rediffmail.com
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


Received 21 January 2014; revised 22 February 2014; accepted 8 March 2014
ABSTRACT
Induced accumulation of polyphenolics and flavonoids in cyanobacterial strains grown under different salt concentrations is correlated with their growth under stress conditions and enhanced antioxidant activity. Plectonema boryanum, Hapalosiphon intricatus, Anabaena doliolum and Oscillatoria acuta grown for 21 days under different salt concentrations (80, 160, 240, 320 and 400 mM) in BG11 medium showed differential growth responses in terms of biomass, total protein, chlorophyll content, total content of polyphenol, flavonoid and carotenoid, accumulation of phenolic acids (gallic, caffeic, chlorogenic, ferullic and vanillic) and flavonoids (rutin and quercetin). Cyanobacterial extracts showed prominent free radical scavenging antioxidant activity (AOA) in terms of % DPPH discoloration. Highly significant (p < 0.05) and strong correlation was found between TPC and AOA (r = 0.974). Other positive but non-significant (p < 0.05) correlations were observed between AOA and gallic acid (r = 0.893) and AOA and caffeic acid (r = 0.931). Significant and strong correlation was also observed between gallic and caffeic acid (r = 0.973). Positive but lesser magnitude correlations were recorded between TPC and caffeic acid (r = 0.905), TPC and gallic acid (r = 0.920), gallic and vanillic acid (r = 0.916) and caffeic and vanillic acid (r = 0.814). An integrated combination of growth parameters, salt-induced accumulation of phenylpropanoids and stress-derived subsequent antioxidant property of cyanobacterial extracts is thought to provide evidence that secondary metabolic changes can act as the possible alternative mechanism to overcome stress-induced damages in cells.
Keywords:Cyanobacteria; Phenylpropanoids; Flavonoids; Antioxidants; Stress Tolerance

1. Introduction
Cyanobacteria are the most ancient and ubiquitous prokaryotic life forms on the earth to exhibit adaptations to diverse extreme environments including high salinity, drought, high and low temperatures and contaminated soils [1] . Various levels of biochemical, molecular, physiological and genetic studies in cyanobacteria have contributed to uncovering the underlying adaptation mechanisms of salt-stress tolerance [2] . Salt stress is an important abiotic factor to affect survival of microbes in aquatic environment and growth of the organism is usually diminished due to high or fluctuating salt concentrations that require organisms to maintain water potential in their cytoplasm [3] . Cyanobacterial adaptations to salt stress generally include expulsion of inorganic ions from the cells to balance osmotic potential and accumulation of osmoprotectants (compatible solutes-sucrose, trehalose, glucosylglycerol [2-0-(a-D-glucopyranosy1)-glycerol; GG], glycine betaine, proline and/or glutamate) to prevent denaturation of macromolecules [4] .
Phenylpropanoids contribute to multi-functional cellular responses in organisms towards biotic and abiotic stimuli. They are the key indicators and mediators of the stress responses in organisms upon exposure to light and ultra-violet intensities [5] , unusual temperatures [6] , resistance to plant pathogens [7] , pests, diseases and various abiotic stresses [8] . Evidences show that these molecules play a critical role in plant growth and development, xylogenesis and flowering and can act as free radical quenchers of stress-induced reactive oxygen species to protect organisms from oxidative damage [9] . Accumulation of secondary metabolites in plants in response to the challenges posed by biotic and abiotic stress stimuli can lead to the adaptation to many environmental factors [10] [11] and phenolic acids and flavonoids are reported to have the potential to serve as antioxidants under stress conditions [12] .
Although positive correlations among the accumulation of polyphenolics, enhanced antioxidant activity and tolerance to stresses have been reported in plant species [13] , only scarce reports are available on phenylpropanoid-mediated salt-stress tolerance responses in cyanobacteria. We demonstrated induced accumulation of phenolic acids and flavonoids in different cyanobacterial species under salt-stress conditions and correlated with enhanced antioxidant activity as a possible alternative mechanism to overcome stress-induced damages in the cells.
2. Materials and Methods
2.1. Organisms and Growth Conditions
Cyanobacterial strains namely Plectonema boryanum (NAIMCC-C-00022), Hapalosiphon intricatus (NAIMCCC-00121), Anabaena doliolum (NAIMCC-C-00001) and Oscillatoria acuta (NAIMCC-C-00017) were obtained from the National Agriculturally Important Microorganisms Culture Collection (NAIMCC), Maunath Bhanjan, India. All the strains were revived in BG11 media (40 ml in 100 ml flasks) from their respective slants. Cultures were bubbled with air containing 1% (v/v) carbon dioxide and were kept under continuous illumination at 70 µ Em−2·S−2 from incandescent lamps with 12 h light–dark cycles at 25˚C ± 2˚C.
Exponentially growing cyanobacterial cultures were inoculated to 500 ml flasks containing 200 ml BG11 media (OD663nm ~ 0.67) supplemented with different concentrations of NaCl (control, 80, 160, 240, 320 and 400 mM). Cultures were finally grown in the culture room under growth conditions mentioned above. Cyanobacterial strains growing in salt-containing and non-salt supplemented media (control) were freshly harvested after 21 days of inoculation, centrifuged and washed thoroughly to remove sodium chloride and then subjected to growth measurement (biomass accumulation) and extracted with suitable solvents for biochemical analysis.
2.2. Extraction
Total phenolic content of cyanobacterial cells was extracted as described by Singh et al., 2011. Briefly, 500 mg cells of cyanobacterial cells were extracted with methanol (methanol/water; 80:20 v/v; 10 ml, thrice) for 4 - 8 h following ultrasonication for 5 min at room temperature. The clear greenish supernatant was collected and mixed with charcoal (1 g) to remove pigments. The solvent was evaporated under vacuum to dryness and finally re-dissolved in methanol by vortexing and stored at 4˚C for bioassay and chemical analysis.
2.3. Biochemical Tests
Chlorophyll and carotenoid was quantified by extracting pigments from 200 mg freshly harvested cyanobacterial cells using methanol: water (9:1, v/v, 4ml, thrice). After removal of the cell debris through centrifugation at 15,000 × g for 5 min, chlorophyll in the supernatant and carotenoid was quantified as per the method mentioned in the reference [14] .
Total protein content (TPC) was extracted thrice from freshly harvested cells (500 mg) with 1% tricholoroacetic acid (4 ml) through sonication and the precipitate was separated by centrifugation at 15,000 × g for 10 min at 4˚C. The cell pellet was resuspended in 1 N NaOH (4 ml), boiled for 30 min, cooled and again centrifuged at 15,000 × g for 5 min. The supernatant was quantified for total content of protein with taking bovine serum albumin (BSA) as standard.
Total polyphenol content (TPC) was estimated spectrophotometrically using the Prussian Blue method and expressed in terms of gallic acid (HiMedia, India) equivalents. Absorbance of the reaction mixture was recorded at 700 nm using UV-VIS spectrophotometer (Shimadzu Corporation, Japan). Analytical grade reagents were used throughout the experiments.
Total flavonoid content was measured by the aluminum chloride colorimetric method [15] . Freshly harvested cells (500 mg) were extracted thrice with methanol (4 ml) and mixed with 0.1 ml of 10% aluminum chloride hexahydrate, potassium acetate (0.1 ml of 1 M) and 2.8 ml of deionized water. After incubation for 40 min at room temperature, the absorbance of the reaction mixture was recorded at 415 nm. Total flavonoid content was expressed in terms of µg rutin equivalents/g fresh cell wt.
2.4. HPLC Analysis
Phenolic extracts of cyanobacterial strains dissolved in methanol were analyzed by High Performance Liquid Chromatograph (HPLC) on Waters model (Waters, USA) equipped with binary Waters 515 reciprocating pumps, variable 2996 photodiode array detector (PDA) and system controller equipped with Waters® Empower™ software for data integration and analysis. Phenolic acids and flavonoids were separated in isocratic conditions on a reverse phase C-18 column (250 × 4.6 mm i.d., 5μm particle size) at 25˚C ± 1˚C at a flow rate of 1 ml/min using methanol/0.4% acetic acid in water (76:24, v/v) as mobile phase. Phenolic acids (gallic, caffeic, ferulic, chlorogenic and vanillic) and flavonoids (rutin and quercetin) were monitored in the eluents at 254 and 280 nm and quantified and identified by comparing the retention times of the peaks in the samples with those in the phenolic standards and by co-injection. HPLC analysis was carried out in triplicate.
2.5. Determination of Antioxidant Activity
Antioxidant activity was measured in terms of DPPH scavenging capacity [16] . Methanolic extract of cyanobacterial strains equivalent to 2000 µg·g–1 gallic acid equivalents was thoroughly mixed with a free radical scavenger 2,2-diphenyl-l-picrylhydrazyl (DPPH) solution (6 × 10–5 mol·l–1; 2.4 mg DPPH dissolved in 100 ml methanol) and the discoloration in the absorbance (A) was recorded at 515 nm at 0 and 30 min. The reaction was carried out in triplicate and methanol was used as blank. Antioxidant activity was calculated as % of discoloration = (1 – [A sample t at 30 min/A control t at 0 min]) × 100.
2.6. Statistical Analysis
All the experiments and biochemical determinations were carried out in triplicate, mean centered and scaled-up to variance prior to Analysis of Variance test using SAS software (version 9.3). Mean differences were analyzed using Duncan’s Multiple Range Test (DMRT) for separations of means significantly differing at p < 0.05. Correlations between the assays were determined by Pearson correlation coefficient at the significance level of p < 0.05.
3. Results
Cyanobacterial strains grown for 21 days under different salt concentrations (80, 160, 240, 320 and 400 mM) in BG11 medium along with the control (no salt) showed differential growth responses in terms of biomass accumulation, total protein and chlorophyll content (Figures 1(a)-(c)). High salt concentrations (400 mM) resulted in slow growth of P. boryanum, H. intricatus, A. doliolum and O. acuta (8.27, 4.67, 1.10 and 5.33 g fresh wt/l, respectively at 400 mM NaCl) in comparison to control (18.27, 10.17, 6.23 and 21.30 g fresh wt/l, respectively). At the same salt concentrations, protein content varied form 2.64, 2.91, 1.43 and 2.83 mg/g fresh wt in comparison to control (3.02, 3.90, 2.35 and 4.84 mg/g fresh wt, respectively) in P. boryanum, H. intricatus, A. doliolum and O. acuta. Chlorophyll content also varied from 80.5, 69.67, 61.47 and 78.27 mg/g fresh wt at 400 mM salt concentration in P. boryanum, H. intricatus, A. doliolum and O. acuta to 116.63, 85.40, 80.47 and 125.03 mg/g fresh wt, respectively in control.
Total phenol, total flavonoids and carotenoid content varied significantly in the cyanobacterial strains treated with different salt concentrations. Increased accumulation of total phenol was observed in all the strains grown under salt conditions in comparison to control (Figure 2(a)). In P. boryanum total phenol accumulation increased with increasing NaCl concentrations (1274.33 to 2714.33 µg/g fresh wt, respectively at 80 and 400 mM NaCl as compared to 1157 µg/g fresh wt in control). In H. intricatus and A. doliolum, the accumulation increased to a maximum of 4300 and 1754 µg/g fresh wt, respectively at 80 mM NaCl but in O. acuta it remained
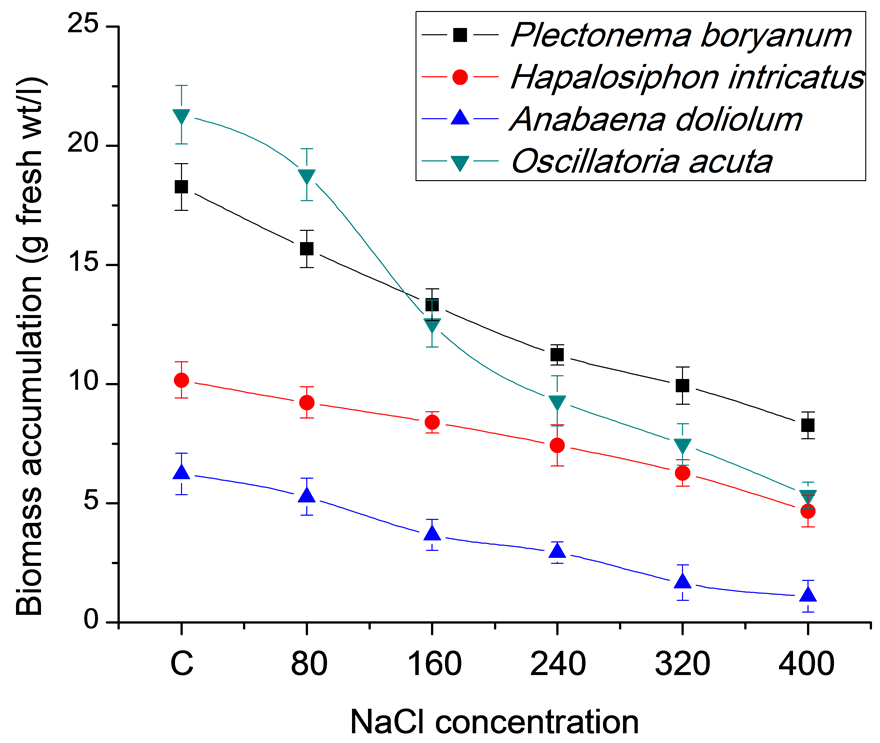 (a)
(a)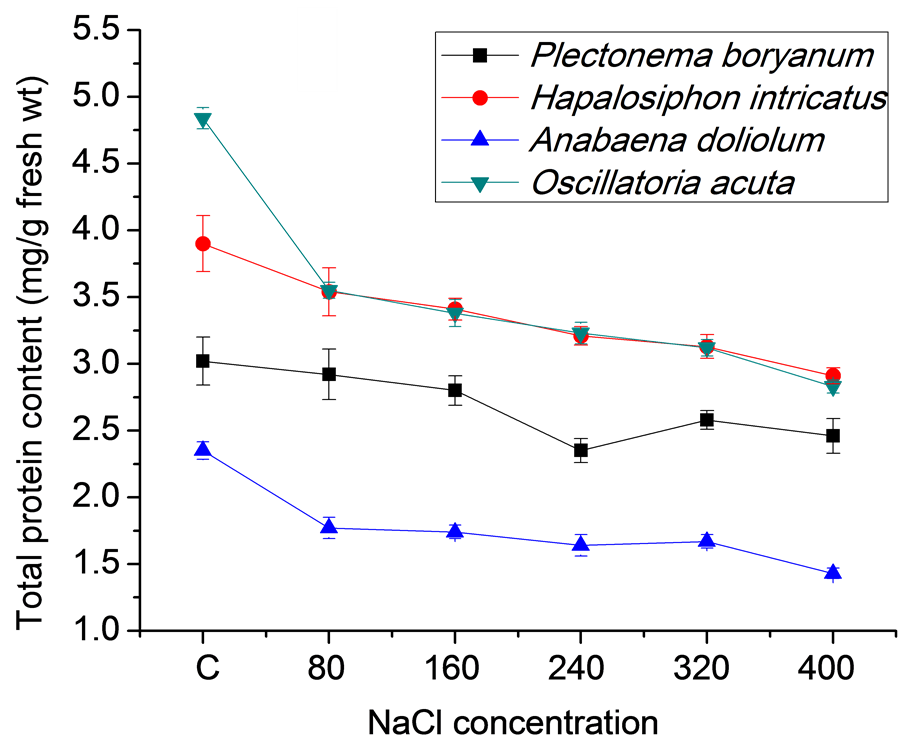 (b)
(b)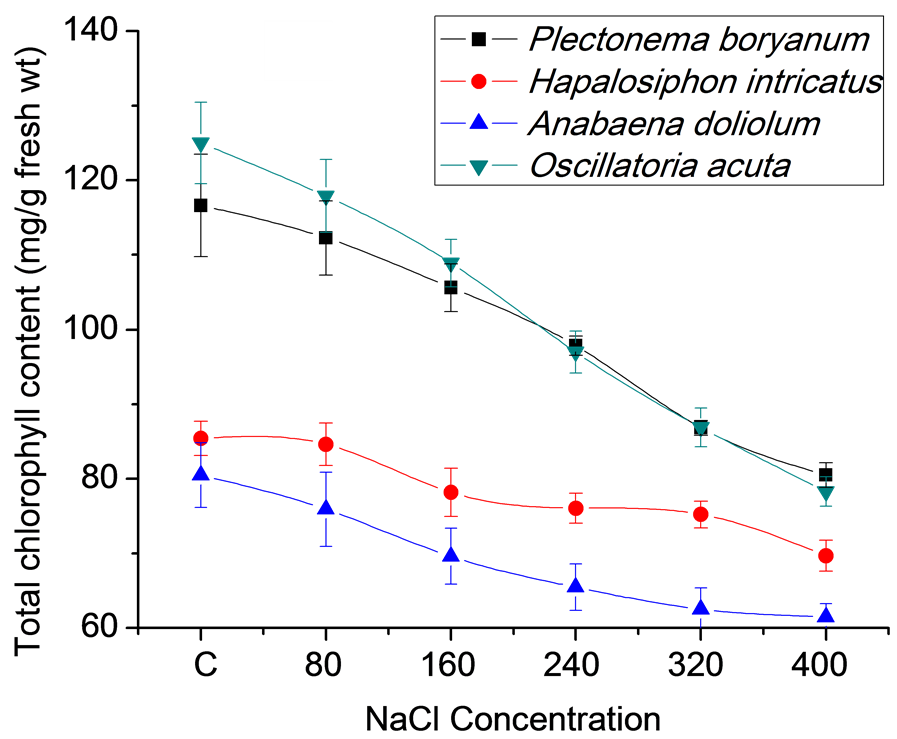 (c)
(c)
Figure 1. Growth characteristics of cyanobacterial strains after 21 days under salt stress conditions. (a) Biomass accumulation; (b) Total protein content and (c) Total chlorophyll content.
2763.00 and 2772.33 µg/g fresh wt at 80 and 160 mM, respectively. Total flavonoid content followed similar pattern as has been observed for total phenol content. Low salt conditions favored high accumulation of total flavonoid accumulation. In P. boryanum, H. intricatus, A. doliolum and O. acuta, the accumulation was 1041, 1129, 3214.33 and 2439.67 µg/g fresh wt respectively at 80 mM NaCl concentration and except in H. intricatus, other cyanobacterial strains showed fairly low content of flavonoid accumulation, even less than the control at 320 and 400 mM NaCl concentration (Figure 2(b)). The accumulation of total carotenoids (TC) in response to salt concentration was negatively affected by increasing the amount of NaCl. Among all the strains, TC content was maximum in H. intricatus (2868.33 µg/g fresh wt) followed by 1940, 1248 and 994 µg/g fresh wt in O. acuta, A. doliolum and P. boryanum, respectively in control while the lowest content varied from 2205 in H. intricatus to 1199.33, 891.33 and 863.33 µg/g fresh wt in O. acuta, P. boryanum and A. doliolum respectively at 400 mM NaCl concentration (Figure 2(c)).
Differential accumulation of individual phenolic acids was observed in cyanobacterial strains grown at different salt concentrations. Gallic acid was the most prominent among all the phenolic acids and its accumulation was higher in organisms grown at all the NaCl concentrations than the control. In H. intricatus and O. acuta maximum gallic acid (177.43 and 143.47 µg/g fresh wt, respectively) was observed at 400 mM NaCl but in P. boryanum and A. doliolum, it was maximum (132.73 and 32.07 µg/g fresh wt) at 320 and 160 mM, respectively (Table 1). Except in O. acuta where caffeic acid accumulation was maximum (16.00 µg/g fresh wt) at 400 mM NaCl, in P. boryanum, H. intricatus and A. doliolum its accumulation was maximum (14.72, 18.48 and
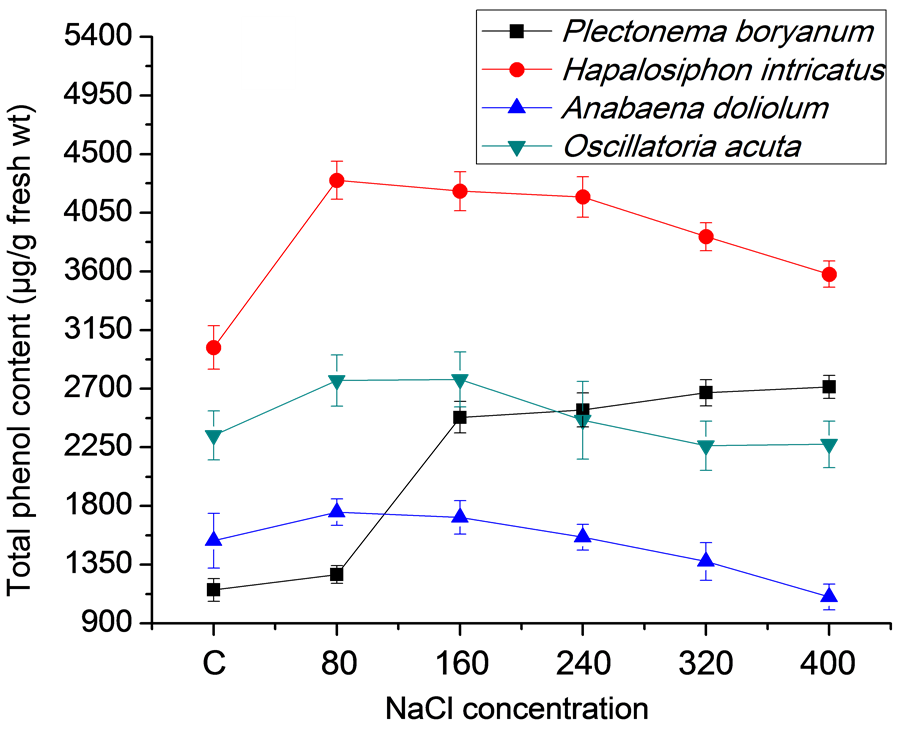 (a)
(a)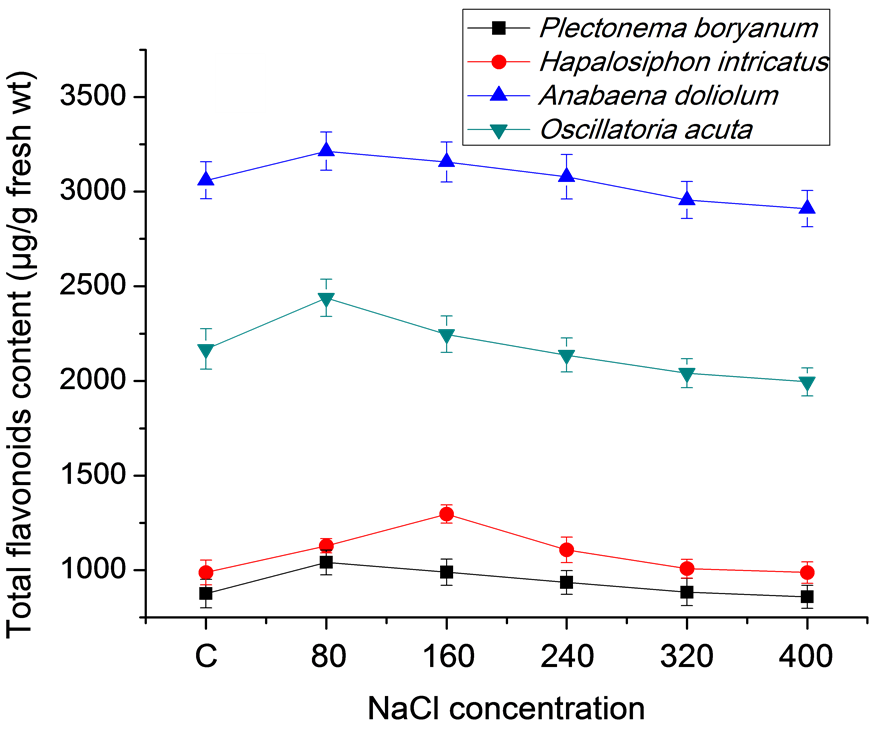 (b)
(b)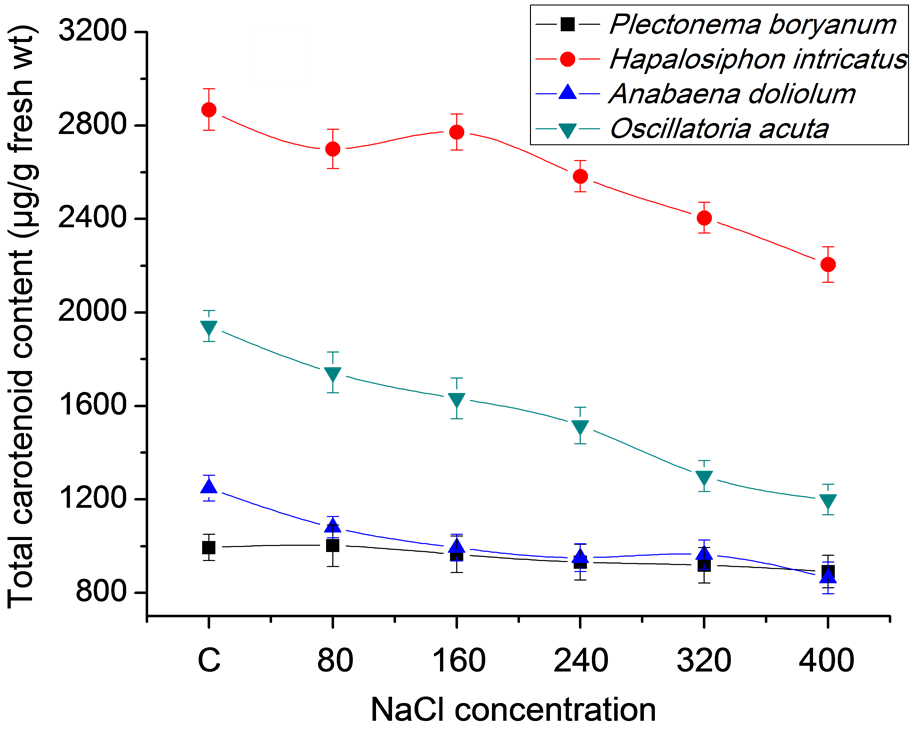 (c)
(c)
Figure 2. Total phenol, flavonoid and carotenoid content in cyanobacterial strains after 21 days under salt stress conditions: (a) Total phenol; (b) Total flavonoids; (c) Total carotenoids content.

Table 1. Phenolic acids in the extract of cyanobacterial strains grown under salt stress conditions after 21 days.
Rt-retention time; Means in the same column followed by different letters are significantly different (p < 0.05).
10.66 µg/g fresh wt, respectively) at 320 mM (Table 1). Differential accumulation of chlorogenic, ferulic and vanillic acids was also observed in cyanobacterial strains grown under salt stress conditions (Table 1). Content of rutin and quercetin (flavonoids) was also enhanced in the NaCl treated strains than in control (Table 2). Except in H. intricatus, rutin content was maximum in all the strains at 400 mM NaCl concentration in which it was maximum (11.13 µg/g fresh wt) at 320 mM. Quercetin, although low in content, was accumulated differentially and maximum accumulation was recorded in A. doliolum (1.82 µg/g fresh wt) at 400 mM (Table 2).
Table 2. Flavonoid content in cyanobacterial strains grown under different salt concentrations after 21 days.
*Retention time in min; Means in the same column followed by different letters are significantly different (p < 0.05).
The free radical scavenging antioxidant activity (AOA) of the cyanobacterial extracts measured in terms of % DPPH discoloration showed promising increase in the AOA with the extracts of strains grown under different NaCl concentrations. Except in P. boryanum extract in which AOA increased from 80 to 320 mM NaCl (9.88% and 21.16%, respectively) (and with the exception at 400 mM (AOA 20.79%), the AOA in all the strains remained almost stabilized (only small decrease is recorded) with increasing NaCl concentration up to 240 mM but decreased severely at 320 and 400 mM (Table 3). However, at 80 mM NaCl concentration, maximum AOA was recorded in H. intricatus (32.85%) while minimum value (9.88%) was found in P. boryanum.
Highly significant (p < 0.05) and strong correlation was found between TPC and AOA (r = 0.974). Other positive but non-significant (p < 0.05) correlations were observed between AOA and gallic acid (r =0.893) and AOA and caffeic acid (r = 0.931) (Table 4). Significant and strong correlation was also observed between gallic and caffeic acid (r = 0.973). Positive but lesser magnitude correlations were recorded between TPC and caffeic acid (r = 0.905), TPC and gallic acid (r = 0.920), gallic and vanillic acid (r = 916) and caffeic and vanillic acid (r = 0.814). However, negative correlations were also observed in certain cases (Table 4).
The magnitude of the variance components of organisms (O), the salt stress (S) and organism × salt stress (O × S) interactions indicates their relative importance for specific antioxidant activities and related parameters. Results summarized in Table 5 compares the proportion of mean squares to the total mean square of each variance component. The variance due to biomass accumulation, total protein content, chlorophyll content, TPC, total flavonoids were found to be highly significant (p = 0.05) with respect to the organism, the stress and O × S (Table 5). Interestingly, the O × S interaction becomes highly significant on all the parameters recorded for cyanobacterial strains following salt stress conditions. The highest impact of growing environment was reflected in terms of the greatest magnitude for O × S interaction for TPC which has high coefficient of variance followed by total flavonoids content (Table 5). The TPC and total flavonoids in all the cyanobacterial strains was least affected under salt stress conditions and followed high degree of accumulation indicating a relative dependence of the organisms on phenylpropanoids under stress conditions.
4. Discussion
Cyanobacteria are known to possess stress-protective enzymatic and non-enzymatic antioxidative mechanisms to alleviate the damages caused due to the reactive oxygen species (ROS) under physiological stresses [17] . Adaptation and tolerance to abiotic stresses, especially to the salt stress in cyanobacteria has widely been worked out in terms of the induction of distinct sets of genes whose products enable successful acclimation [18] , changes in physiological conditions like photosynthesis, protein, carbohydrate, lipids and fatty acid biosynthesis [19] [20] , biochemically active extrusion of toxic inorganic ions and accumulation of compatible solutes including sucrose, trehalose, glucosylglycerol and glycine betaine [21] . Recently, “omics” approaches helped to elucidate salt acclimation processes through differential expression of hundreds of genes and essentially revealed functions of many upregulated proteins in salt-stressed cells under specific salt-sensing mechanisms in salt-resistant cyanobacterial strains [21] . However, antioxidative property entwined with the alterations in phenylpropanoid secondary metabolites in cyanobacteria under salt-stress conditions and its overall impact on
Table 3. Antioxidant activity of cyanobacterial extracts in terms of % DPPH discoloration.
*% of remaining DPPH in the presence of cyanobacterial methanolic extract; means in the same column followed by different letters are significantly different (p < 0.05).
Table 4. Correlation coefficients among total phenolics, antioxidant activity and phenolic acid content in cyanobacterial strains.
*Correlation is significant at the 0.05 level.
Table 5. Variance components (type III sum of squares) for Organisms (O), salt stress(S), and O × S interaction effects for biomass accumulation.
*significant (p < 0.05). Significance is based on the mean square value of organisms × salt stress.
stress tolerance has not been specifically resolved.
Increase in the total polyphenols in macroalgae during high salinity and its possible role in protection to the oxidative stress-induced damage in plants is documented [22] . Induced accumulation of TPC under salt stress conditions followed by enhanced antioxidant activity in all the cyanobacterial strains validates early reports.
Similarly, phenolic acids especially gallic, chlorogenic, ferulic and vanillic acids and flavonoids rutin and quercetin are strong free radical scavengers and thereby, possess antioxidative property [23] . Carotenoids can quench and thus, minimize reactive oxygen species (ROS) formation but their specific mechanism under salt stress conditions in specific species is still uncertain. Interestingly, the content of carotenoids has been reported to decrease in response to salt stress and this is also reflected by our results. Carotenoids positively influence preventive antioxidative defense but more investigation is needed to verify this suggestion [24] .
We reported the impact of salt stress on the cyanobacterial phenylpropanoid accumulation that in turn enhances antioxidative property in the organisms leading to the tolerance to varied salt concentrations. This is evident from a positive correlation between accumulation of phenylpropanoids and antioxidant activity. Furthermore, the composition of polyphenolics such as gallic, caffeic, chlorogenic, ferulic and vanillic acids and flavonoids (rutin and quercetin) has shown to be differentially accumulated in a salt concentration-dependent manner except in certain circumstances, in comparison to the un-stressed control. Besides other known mechanisms of salt and osmotic stress tolerance and adaptation, accumulation of secondary metabolites, especially phenylpropanoids, which has scarcely been reported in cyanobacteria in stress-alleviating context, could be another important strategy for the cyanobacterial organisms to protect cellular damage.
Acknowledgements
DPS is grateful to the Indian Council of Agricultural Research, India for financial support in the form of Institute Project. Help rendered by Dr. Mahesh Yandigeri in statistical analysis is gratefully acknowledged.
References
- López-Rodas, V., Maneiro, E. and Costas, E. (2006) Adaptation of Cyanobacteria and Microalgae to Extreme Environmental Changes Derived from Anthropogenic Pollution. Limnetica, 25, 403-410.
- Pandhal, J., Wright, P.C. and Biggs, C.A. (2008) Proteomics with a Pinch of Salt: A Cyanobacterial Perspective. Saline Systems, 4, 1-18. http://dx.doi.org/10.1186/1746-1448-4-1
- Mikkat, S., Hagemann, M. and Schoor, A. (1996) Active Transport of Glucosylglycerol Is Involved in Salt Adaptation of the Cyanobacterium Synechocystis sp. Strain PCC 6803. Microbiology, 142, 1725-1732. http://dx.doi.org/10.1099/13500872-142-7-1725
- Allakhverdiev, S.I., Klimov, V.V. and Hagemann, M. (2005) Cellular Energization Protects the Photosynthetic Machinery against Salt-Induced Inactivation in Synechococcus. BBA-Bioenergetics, 1708, 201-208. http://dx.doi.org/10.1016/j.bbabio.2005.01.002
- La Camera, S., Gouzerh, G., Dhondt, S., Hoffman, L., Frittig, B., Legrand, M. and Heitz, T. (2004) Metabolic Reprogramming in Plant Innate Immunity: The Contributions of Phenylpropanoid and Oxylipin Pathways. Immunological Reviews, 198, 267-284. http://dx.doi.org/10.1111/j.0105-2896.2004.0129.x
- Janas, K.M., Cvikrova, M., Palagiewicz, A. and Eder, J. (2000) Alterations in Phenylpropanoid Content in Soybean Roots during Low Temperature Acclimation. Plant Physiology and Biochemistry, 38, 587-593. http://dx.doi.org/10.1016/S0981-9428(00)00778-6
- Singh, U.P., Sarma, B.K. and Singh, D.P. (2003) Effect of Plant Growth Promoting Rhizobacteria and Culture Filtrate of Sclerotium rolfsii on Phenolic and Salicylic Acid Contents in Chickpea (Cicer arietinum). Current Microbiology, 46, 131-140. http://dx.doi.org/10.1007/s00284-002-3834-2
- Vogt, T. (2010) Phenylpropanoid Biosynthesis. Molecular Plant, 3, 2-20. http://dx.doi.org/10.1093/mp/ssp106
- Oueslati, S., Karray-Bouraoui, N., Attia, H., Rabhi, M., Ksouri, R. and Lachaal, M. (2010) Physiological and Antioxidant Responses of Mentha pulegium (Pennyroyal) to Salt Stress. Acta Physiologiae Plantarum, 32, 289-296. http://dx.doi.org/10.1007/s11738-009-0406-0
- Edreva, A., Velikova, V., Tsonev, T., Dagnon, S., Gürel, A., Aktaş, L. and Gesheva, E. (2008) Stress-Protective Role of Secondary Metabolites: Diversity of Functions and Mechanisms. General and Applied Plant Physiology, XXXIV, 67-78.
- Ramakrishna, A. and Ravi Shankar, G.A. (2011) Influence of Abiotic Stress Signals on Secondary Metabolites in Plants. Plant Signal Behaviour, 6, 1720-1731. http://dx.doi.org/10.4161/psb.6.11.17613
- Agati, G., Biricolti, S., Guidi, L., Ferrini, F., Fini, A. and Tattini, M. (2011) The Biosynthesis of Flavonoids Is Enhanced Similarly by UV Radiation and Root Zone Salinity in L. vulgare Leaves. Journal of Plant Physiology, 168, 204-212. http://dx.doi.org/10.4161/psb.6.11.17613
- Korkina, L.G. (2007) Phenylpropanoids as Naturally Occurring Antioxidants from Plant Defence to Human Health. Cellular and Molecular Biology, 53, 15-25.
- Ferjani, A., Mustardy, L., Sulpice, R., Marin, K., Suzuki I., Hageman, M. and Murata, N. (2003) Glucosylglycerol, a Compatible Solute, Sustains Cell Division under Salt Stress. Plant Physiology, 131, 1628-1637. http://dx.doi.org/10.1104/pp.102.017277
- Kim, D.O., Jeong, S.W. and Lee, C.Y., (2003) Antioxidant Capacity of Phenolic Phytochemicals from Various Cultivars of Plums. Food Chemistry, 81, 321-326. http://dx.doi.org/10.1016/S0308-8146(02)00423-5
- Beta, T., Nam, S., Dexter, J.E. and Sapirstein, H.D. (2005) Phenolic Content and Antioxidant Activity of Pearled Wheat and Roller Milled Fractions. Cereal Chemistry, 82, 390-393. http://dx.doi.org/10.1094/CC-82-0390
- Kesheri, M., Richa and Sinha, R.P. (2011) Antioxidants as Natural Arsenal against Multiple Stresses in Cyanobacteria. International Journal of Pharma and Bio Sciences, 2, B169-B187.
- Kanesaki, Y., Suzuki, I., Allakhverdiev, S.I., Mikami, K. and Murata, N. (2002) Salt Stress and Hyperosmotic Stress Regulate the Expression of Different Sets of Genes in Synechocystis sp. PCC 6803. Biochemica Biophysica Research Communication, 290, 339-348. http://dx.doi.org/10.1006/bbrc.2001.6201
- Tang, D., Shi, S., Li, D., Hu, C. and Liu, Y. (2007) Physiological and Biochemical Responses of Scytonema javanicum (cyanobacterium) to Salt Stress. Journal of Arid Environments, 71, 312-320. http://dx.doi.org/10.1006/bbrc.2001.6201
- Allakhverdiev, S.I. and Murata, N. (2008) Salt Stress Inhibits Photosystems II and I in Cyanobacteria. Photosynthetic Research, 98, 529-39. http://dx.doi.org/10.1007/s11120-008-9334-x
- Hagemann, M. (2011) Molecular Biology of Cyanobacterial Salt Acclimation. FEMS Microbiology Review, 35, 87-123. http://dx.doi.org/10.1111/j.1574-6976.2010.00234.x
- Mpofu, A., Sapirstein, H.D. and Beta, T. (2006) Genotype and Environmental Variation in Phenolic Content, Phenolic Acid Composition, and Antioxidant Activity of Hard Spring Wheat. Journal of Agricultural and Food Chemistry, 54, 1265-1270.
- Karamac, M., Kosiñska, A. and Pegg, R.B. (2005) Comparison of Radical-Scavenging Activities for Selected Phenolic Acids. Polish Journal of Food Nutrition Science, 14, 165-170.
NOTES
*Corresponding author.