Paper Menu >>
Journal Menu >>
 Advances in Anthropology 2012. Vol.2, No.3, 139-156 Published Online August 2012 in SciRes (http://www.SciRP.org/journal/aa) http://dx.doi.org/10.4236/aa.2012.23017 Copyright © 2012 SciRes. 139 Haplogroup R1a, Its Subclades and Branches in Europe during the Last 9000 Years Igor L. Rozhanskii, Anatole A. Klyosov* The Academy of DNA Genealogy, Newton, USA Email: *aklyosov@comcast.net Received May 5th, 2012; revised June 11th, 2012; accepted June 25th, 2012 This study identifies and describes 38 branches of the haplogroup R1a STR haplotypes which currently exist in Europe or which migrated from Europe to areas in the east, south, and southeast between 6000 and 4500 years before the present (ybp). The study is based on 2471 haplotypes which have been tested for either 67- or 111-markers; it essentially creates a unified robust system, which assembles dozens of R1a-SNPs and thousands of STRs and assigned haplotypes to branches, some of which do not have SNP assignments as yet. The assembled system consists of base (deduced ancestral) haplotypes, one for each STR branch and for each SNP-assigned subclade, each with its characteristic (ancestral) set of alleles, ar- ranged in the chronological space from ~9000 ybp to 1300 ybp. We found that the most ancient R1a sub- clades (R1a1-M198− and R1a1a-M198+/M417−) bearers of which currently live in Europe (the present day haplotypes are scattered between England and the Balkans) appeared in Europe at least 7300 ybp, and possibly 9000 ybp. R1a’s three principal downstream subclades, L664 (North-Western branch), Z93 (South-Eastern branch), and Z283 (Eurasian branch), split from their common European ancestor at about the same time, around 7000 - 6000 ybp. L664 apparently stayed in North-Western Europe; its lineage re- covered and began expanding ~4575 ybp. The Z93 subclade began to expand during the Aryan migra- tions, on the Aryan’s journey to India and the Middle East in the 3rd-2nd millennia BC. The Z283 sub- clade split ~5500 ybp into three branches. One of them, Z280 (the Central Eurasian branch) moved east to the Russian Plain in 4800 - 4600 ybp, and formed at least 16 sub-branches there and in the course of the later westward repopulation of Europe in the 1st millennium BC—1st millennium CE. Some of the older branches, like the Russian Plain branch, largely stayed in the present Russia-Ukraine-Belarus-Poland- Baltic countries region, and were described by early historians as the Scythians, Antes, Veneti, and a mul- titude of different proto-Slavic tribes (though many of them belonged to haplogroups other than R1a, primarily I1 and I2). Those R1a branches which are “older” than 3000 years, such as the Russian Plain branch (4600 ybp), the Western Eurasian (4300 ybp), and the Balto-Carpathian (4300 ybp), did not move en mass to Europe but stayed behind at the Russian Plain. In the middle of 1st millennium CE, the time of the collapse of the Roman Empire, multiple migrations of R1a were taking place eastward and westward; these migrations gradually formed the current landscape of R1a in Europe. All 38 branches and their dat- ings are listed in the Appendix of this paper; current distribution maps are shown in the body of the paper. Keywords: Y Chromosome; Mutations; Haplotypes; Haplogroups; TMRCA; STR; SNP; R1a1 Introduction The preceding article in this journal (Klyosov & Rozhanskii, 2012) provided data in support of the theory that haplogroup R1a arose in Central Asia around 20,000 years before the pre- sent (ybp), and described the prior history of the haplogroup, which is directly related to the appearance of Europeoids (Cau- casoids) ~58,000 years ago. At some point, apparently between 20 and 15 thousand ybp, the bearers of R1a began a migration to the west, through Tibet and over the Himalayas. They arrived in Hindustan no later than 12,000 ybp. They apparently continued their way across the Iranian Plateau, along East Anatolia and the rest of Asia Minor between 10,000 and 9000 ybp. By ~9000 ybp they ar- rived in the Balkans and spread westward over Europe and to the British Isles. At that point, R1a still had DYS392 = 13 in most haplotypes, as did its sister haplogroup R1b. This marker is very slow to change, and mutates on average once in 3500 conditional generations (that is once in 87,500 years in one DNA-lineage; the concept of conditional generations of 25 years in each is described in “Materials and Methods” section). More than 99% of the R1a haplotypes known today bear the mutation giving DYS392 = 11. Somewhere along this extended timescale, bearers of R1a or its upstream haplogroups devel- oped the Pre-proto-Indo-European (Pre-IE) language and car- ried it along during their journey from Central Asia to Europe. Linguists date the earliest signs of Pre-IE in Anatolia at 9600 ± 600 ybp (Gamkrelidze & Ivanov, 1995; Gray & Atkinson, 2004; Renfrew, 2000), which coincides with the data of DNA gene- alogy for the migration of R1a bearers across Asia Minor that is described in Klyosov and Rozhanskii (2012). Some known archaeological cultures in the Balkans and Cen- tral/Eastern Europe, dated 8000 - 7000 ybp (Bug-Dniester, Vinča, Starčevo, LBK, etc.), can be attributed, as least in part, to bearers of R1a. Yet they also can be attributed to other an- cient haplogroups, such as I, J, E, G. As the bearers of haplogroup R1b1a2 began to populate *Corresponding author.  I. L. ROZHANSKII, A. A. KLYOSOV Europe after 4800 ybp (the Bell Beakers and other R1b1 mi- gratory waves, including, perhaps the Kurgan people, though their identification and haplogroup assignment remain unclear, and likely represented both R1b and R1a bearers in different time periods), haplogroup R1a had moved from Central and North-Western Europe to the Russian Plain around 4800 - 4600 ybp (Klyosov, 2011). From there, R1a migrated to the south, east, and south-east as the historic Aryans (Klyosov & Roz- hanskii, 2012). We have to emphasize that the definition of the Aryans in DNA genealogy is broader than the definition of Aryans in linguistics and archaeology. According to linguists and archae- ologists, Aryans were the ancient people who spoke Indo- European Aryan or “steppe” languages; this early group split at some point into the Indo-Aryan and Iranian languages. When and where that split occurred is a matter of debate; linguists generally claim that the split occurred immediately before the Aryans moved to India and Iran, while some archaeologists believe that it happened a thousand years before that. In DNA genealogy, with its emphasis on genetic heritage, the Aryans, as the putative bearers of Indo-European dialects, also clearly carried haplogroup R1a, which apparently was brought to India and Iran coincident in time with the spread of Indo- European (IE) dialects in those same regions. That time is gene- rally recognized around 3500 ybp, since studies of Gordon Childe (Childe, 1926) and even earlier. In other words, R1a likely represented a substantial portion of the Aryan linguistic and cultural migration, the roots of which we can trace back to Europe and to earlier migrations, following R1a ancient move- ments. As soon as we accept such a definition, the time of the split of the Aryans into “Indo-Aryans” and “Iranians” becomes evident: migration is swift but language change is slow. The “language split” must have occurred before the Aryan entrance into Anatolia and northern Syria (Mitanni). This reasoning suggests that the split was around 4500 ybp—as the archaeolo- gists predicted. Today’s linguists find IE languages in Anatolia and Mitanni, the South Urals, Iran, India, beyond the Ural Mountains, and in South Siberia. All these languages have the same Aryan roots. These peoples developed common horse-breeding terminology and shared essentially the same vocabulary for household items, and for gods and religion, although sometimes, apparently, the shared words ended up with contrasting meaning, as in India and Iran, where some ancient Aryan good and evil gods some- times had opposite meanings. Currently, most of descendants of the ancient peoples who carry European R1a live in Eastern Europe, primarily in Russia (up to 63% of the population) and Poland, Ukraine, Belarus (up to 57% of the population in the last three countries) (Underhill et al., 2009; Balanovsky et al., 2008; Kayser et al., 2005; Khar- kov et al., 2004, 2005). Haplogroup R1a and its regional distribution and history were a subject of many studies, which have been reviewed in the preceding paper (Klyosov & Rozhanskii, 2012). R1a was identified in excavated DNA in Germany with the dating of 4600 ybp (Haak et al., 2008), in Siberia, with the dating of 3800 - 3400 ybp (Keyser et al., 2009), in Tarim Basin, Xinjiang, China, with the dating of ~4000 ybp (Li et al., 2010). Its distri- bution was described in various parts of Eurasia (e.g., Wells et al., 2001; Behar et al., 2003; Cinnioglu et al., 2004; Kharkov et al., 2004; Sengupta et al., 2006; Balanovsky et al., 2008; Sharma et al., 2009; Underhill et al., 2009; Abu-Amero et al., 2009). However, only in the last few years, thank mainly to efforts of Dr. Peter Underhill’s laboratory at Stanford Univer- sity, Dr. Thomas Krahn of Family Tree DNA’s Genomics Re- search Center, and researchers of 1000 Genomes Project, many new SNPs in haplogroup R1a have been discovered; as indi- cated by beginning letters M, L, and Z, respectively, in the SNPs below. Their base haplotypes, mutation patterns and his- tory have not been described in the literature as yet. Apparently, only one attempt was made regarding R1a-M458 subclade (Underhill et al., 2009), however, the authors employed the erroneous “evolutionary mutation rate” (see Klyosov & Roz- hanskii, 2012, and references therein) and obtained a grossly overestimated date for M458 appearance as 10,700 ybp in Eastern Europe (Poland), about 250% higher than the figure obtained in our studies. Commonly, 250% - 300% exaggeration in historical datings is a “feature” of the “evolutionary mutation rate” method (ibid.). This study focuses on DNA lineages, or branches, of hap- logroup R1a in Europe, from the Atlantic to the Ural Mountains, and on the history of their appearance and migrations across and from Europe in the last ~9000 years (Appendix 1). The methodology of the study, based largely on considerations of extended 67- and 111-marker haplotypes, and involving prin- cipal methods of DNA genealogy, is described in detail in the preceding papers in this journal (Rozhanskii & Klyosov, 2011; Klyosov & Rozhanskii, 2012) and in the Materials and Meth- ods section of this article. Available databases containing tens of thousands of 67-marker haplotypes are listed in Rozhanskii and Klyosov (2011) and in this paper (Appendix 2). The Old European Branch (≥9000 ybp) (R1a-SRY10831.2+, M417−) The Old European Branch of R1a in Europe has a distinct ancient DYS392 = 13, as does its sister haplogroup R1b (see the diagram below). However, while in R1b this allele is by far the predominant one, DYS392 = 13 occurs in R1a in only 0.4% of all haplotypes. All others have DYS392 = 11. Actually, only twelve DYS392 = 13 haplotypes have been identified thus far in the 67-marker haplotype format, and nine of them form the Old European branch (the haplotype tree of this branch is shown in Figure 1). The following diagram shows the upper part of the hap- logroup tree with respect to R1a and its most ancient subclades (R1b is shown for a comparison) (ISOGG-2012, a fragment, http://www.isogg.org/tree/ISOGG_HapgrpR.html) in its current classification: R M207 R1 M173 R1b M343 R1a L62/M513, L146/M420 R1a1 L120/M516, SRY10831.2 R1a1a M17, M198, M512 R1a1a1 L457, M417 The two haplotypes at the bottom of the tree in Figure 1 are from Belarus and the US, both have positive SNP SRY10831.2 and negative M198, which firmly places them into the R1a1 subclade on the haplotype tree above. They two have their combined haplotype (here and throughout the paper haplotypes are given in the FTDNA format): Copyright © 2012 SciRes. 140  I. L. ROZHANSKII, A. A. KLYOSOV Figure 1. The 67-marker 9 haplotype tree for the Old European Branch in Europe. This and all other haplotype trees in this paper are composed from haplotypes listed in the IRAKAZ database (Rozhanskii et al., 2012). The upper 7-haplotype branch has SNP M417–, M198+ (R1a1a), the lower 2-haplotype branch has M198–, SRY- 10831.2+ (R1a1). 13 25 15 10/11 12 12 12 13 12 13 13 31/33—16 9 9 11 11 24/25 16 20/21 29/34 12/13 16 16 16—10/11 11/12 19 23 15 15/16 17/20 17/18 35 38 11/12 11—11 8 15 17 8 11 10 8 11/12 10 12 22 22 17 10 12 12 15 8 12 22/23 22/23 13 12 11 13/14 10/11 11 12 12/13 The 25 mutations between them are translated to 25/.12 = 208 → 262 conditional generations (25 years each), or 6550 years. If they belong to the same DNA-lineage, their most re- cent common ancestor lived ~3300 ybp. If they belong to dif- ferent lineages, their common ancestor would have lived much earlier. The upper, rather tight series of haplotypes in Figure 1 all have positive M198, but negative M417, which places them in the R1a1a subclade. Their base haplotype is as follows: 13 25 15 11 13 14 12 12 10 14 13 31—16 9 10 10 11 25 14 19 31 12 15 15 15—10 11 19 23 16 16 17 19 36 38 11 11—11 8 17 17 8 12 10 8 12 10 12 22 22 15 10 12 12 13 8 13 23 22 12 12 11 13 11 11 12 12 All seven haplotypes have 78 mutations from the base hap- lotype, which gives 78/7/.12 = 93 → 103 conditional genera- tions, or 2575 ± 390 years to their common ancestor. However, this date actually points to a time when the population ex- panded. In reality, the base haplotype shown above differs from the two M198– haplotypes by 40 and 41 mutations respectively; if they belonged to the same subclade, that would sets them apart by at least 12,300 and 12,800 years. Since they belong to different subclades (R1a1 and R1a1a), their common ancestors lived more than 6400 years before the present, and not less than the age of the R1a1a-M17 subclade (at least 9000 ybp). We do not know exactly where their common ancestor lived, since members of this branch are scattered in Europe from the descendants of those R1a bearers who came to the Balkans Balkans to the British Isles (Map 1). It may be that they are e a particular su Principal European R1a Subclades As wahanskii (2 nstream subclade in Europe was R1a1a1-M417, w R1a1a1 M417 ~9000 ybp at the European end of their migration route from Central Asia (Klyosov & Rozhanskii, 2012). In this paper, unless we specifically describ bclade with its particular SNP, we will use R1a as a common overall designation of all downstream subclades of the hap- logroup. s described above and in Klyosov and Roz 012), R1a arose around 20,000 ybp in Central Asia, migrated (in part) to Europe over the following 11,000 years, and arrived in Europe, as R1a1 and R1a1a subclades, more than 7 thousand years or, more probably, ~9000 ybp. There are only a few de- scendants of these ancient subclades which are currently identi- fied in Europe. The next dow hich arose ≥7600 ybp (see below). Its principal downstream subclades are as follows: R1a1a1i L664– (the Tenths, NW branch) R1a1a1i L664+ R1a astern branch) 1a1h Z93 (South-E R1a1a1g Z283 (Eurasian branch) R1a1a1g1 M458 (European branch) R1a1a1g2 Z280 (Central Eurasian) R1a1a1g3 Z284 (Scandinavian) The Tenths, or the North-Western branch, is the oldest sub- cl lotype tree is shown in Fig- ur ain purpose of presenting the overall tree in Figure 2 is North-Western Branch (the Tenths), 4575 ybp s ade of R1a-M417, identified thus far. It arose at least 6300 ybp (see below) and has a distinct mutation DYS388 = 10 as distinct from the DYS388 = 12 in majority of R1a haplotypes. Subclade Z93 separated from the rest of R1a around 5700 ybp. It migrated from Europe or from the Russian Plain no later than 4800 ybp. The migration was to the south (to the Middle East) and south-east (Iran and India), where bearers of Z93 are the predominant R1a bearers today. Subclade Z283 arose ~5500 ybp and populated Eastern Europe, part of Central Europe, and further east to the Ural Mountains. The overall 111-marker R1a hap e 2. The purpose of the Figure is not in details of positions of individual haplogroups, but to show the overall shape, configu- ration of the tree, and relative positions of its subclades. A few of M420 and M417 haplotypes, available in the 111-marker format, sit at the very base/core of the tree, since they de- scended from the most ancient European R1a ancestors. The principal subclades of the Z283 Eurasian super-branch are lo- cated around the tree—Z280, Z284, M458. Some of them con- sist of two or several more sub-branches, which are described below. The m to show that the SNP (single nucleotide polymorphism) sub- clades and STR (short tandem repeat) branches form a robust mutually supporting system. They are two sides of the prover- bial coin; both are necessary for a justified, meaningful, and advanced analysis of haplotype datasets. This is one the most distinct of R1a branches. All haplotype Copyright © 2012 SciRes. 141  I. L. ROZHANSKII, A. A. KLYOSOV Copyright © 2012 SciRes. 142 Map 1. European Branch, referred to birthplaces of the furthermost ancestors recorded in available data- The Old bases. Red pins denote a branch with SNP M198−, SRY10831.2+. Blue pins mark a branch with M417−, M198+. Figure 2. rker haplotype tree of R1a haplogroup, composed of 251 f this branch have DYS388 = 10 (hence, the Tenths), and two- live in the USA (with roots mainly in England, Ireland, Scan- e is the old branch (with a common ances- to -branches as follows: 24 16 15 19 20 33 39 12 11—11 23 15 16 18 19 34 37 12 11—11 a common ancestor of 2125 ± 280 An 111-ma haplotypes. Most of the tree is taken by the Eurasian super-branch Z283, of which branches Z280 (Central Eurasian), Z284 (Scandinavian), and M458 (European) are shown. Another two principal branches are Z93 (South-Eastern) and L664 (North-Western branch, the Tenths). The L664 branch shown at 7 o’clock. o thirds of them are located in England, Ireland and Northern Germany (Map 2), and almost the same number of their bearers dinavia, Germany). The haplotype tree (Figure 3) splits into four major lineages, or sub-branches. On r of ~4300 ybp), which in turn splits to subclades L664- (4100 ybp; though, this SNP seems to be rather unstable) and L664+ (4300 ybp), then the young branch, with L664+ (2250 ± 250 ybp), and two DYS448 null-mutated branches, of 1500 ± 325 ybp and its downstream of 260 ± 60 ybp, the last marks the arrival of the Tenths to the USA (Klyosov, 2010a, 2010b, 2012). The base 111-marker haplotype of the old branch splits into two sub 13 25 16 10 11 14 12 10 10 13 11 30—15 9 10 11 11 24 14 19 31 12 15 15 16—11 11 19 8 17 17 8 12 10 8 11 10 12 22 22 16 10 12 12 14 8 14 23 22 12 13 11 13 11 11 12 13—31 15 9 16 12 26 27 18 13 12 1212 11 10 12 11 10 11 11 31 12 15 24 12 9 10 19 15 20 12 23 15 12 15 25 12 23 19 10 15 18 9 11 11 13 25 15 10 11 14 12 10 10 13 11 31—15 9 10 11 11 25 14 19 31 12 14 14 17—12 11 19 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 14 23 22 12 12 11 13 11 11 14 13—32 15 9 15 12 26 27 19 12 121212 10 9 12 11 10 11 11 31 12 14 24 13 9 10 19 15 20 12 24 15 12 16 24 12 24 19 10 15 19 9 11 11 which differ by as much as 33 mutations. One branch (shown first) is likely L664−, with ybp (.139 mutations per marker), another, likely L664+, is 2025 ± 260 years “old” (.133 mutations per marker). The distance between them sets these two lineages apart by 33/.198 = 167 →  I. L. ROZHANSKII, A. A. KLYOSOV Map 2. The map of the North-Western branch, depicting birthplaces of the furthermost ancestors recorded in available Yellow pins denote an old branch. Deep-blue and light-blue pins mark younger branches. databases. Figure 3. A haplotype tree of the Tenth’s (DYS388 = 10), composed of 285 o The dataset was provided by Martin Voorwin- 0 years. This placed their common ancestor by (2125 + 2025 + 5000)/2 = 4575 ybp. haplotype tree (ISOGG-2012) f 67-marker haplotypes. den and updated with the Prendergast family series in January 2012. The younger branch on the left all has SNP L664+, where determined; the branch on the 3 o’clock (the Prendergast family with a common ancestor ~1000 ybp) has both L664+ and L664−, which raises a ques- tion on technical (methodological) stability of the L664−. The old branch has both L664+ and L664−. An analysis of a more extended dataset of 67-marker haplo- 200 conditional generations, or 500 types of the Tenths gave 4300 years to their common ancestor, which is the same value within the margins of error, and showed that the divergence of L664+ and L664− branches oc- curred around 4100 ybp, or almost from the common ancestor of currently living Tenths (Klyosov, 2012). The robustness of the calculations despite the small number of extended haplotypes is clear. Twenty-one 67-marker haplo- types show 191 mutations, which give 191/21/.12 = 76 → 83 generations, or 2075 years to a common ancestor. The same haplotypes in the 111-marker format give 324 mutations, that is 324/21/.198 = 78 → 85 generations, or 2125 years to the com- mon ancestor. Obviously, the margins of error of 2075 ± 260 and 2125 ± 240 ybp may be somewhat excessive for the calcu- lations. The combined 67-marker base haplotype of the Tenths dif- fers from the Old European base haplotype (see above) by 29 mutations, which places their common ancestor at ≥7600 years before the present. Since they belong to different subclades, M417− and M417+, their common ancestor lived 7600 ybp only if the “depth” of the M417+ subclade is no more than 7600 years. In that case, the Tenths actually initiated the M417+ subclade. South-Eastern Branch (Z93), ~5700 ybp According to the Copyright © 2012 SciRes. 143  I. L. ROZHANSKII, A. A. KLYOSOV R1a1a1h Z93 R1a1a1h1 Z94 R1a1a1h1 L342.2 R1a1a1h1a L657 subclade Z93 has tades, Z94, L342.2 and 657. All four subcnly the Middle East, hree downstream subcl lades populate maiL India, and some locations in the eastern part of the Russian Plain, hence, it is called the South-Eastern branch (Map 3). It arose in Europe ~5700 ybp and migrated along with the future Aryans across the Russian Plain to the south, east, and south- east between 5000 and 3500 ybp. The haplotype tree of the R1a-Z93 clade is shown in Figure 4. Its Jewish R1a1 branch includes more than half of the whole tree (104 haplotypes out of 203 haplotypes); all the Jewish haplotypes are related to the downstream subclade L342.2. The Bashkir and the Kyrgyz branches also have the same L342.2. In fact, L342.2 is the predominant subclade in the Z93 clade, em- bracing 98% of its haplotypes. Only a few haplotypes on the tree and one English lineage of closely related individuals are L342.2 negative, which tells us the initial state of Z93. Bearers of Z93 are spread from England through Armenia, Oman, and Kuwait, all the way to Khakassia in South Siberia; the latter (or his ancestors) apparently was a migrant from the Russian Plain. Six currently known representatives have the following base haplotype 13 24 16 11 11 15 12 12 10 13 11 30—15 9 9/10 11 11 24 14 20 31 12 15 15 15—11/12 11 19 22/23 16 16 18 17 35 40 13 11 —11 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 13/14 23 21 12 12 11 13 10 11 12 13 All six have 111 mutations from the base haplotype which gives 111/6/.12 = 154 → 177 generations, or 4425 ± 630 years to their common ancestor. In fact, they belong to different mi- nor sub-branches, which split from the trunk at the very begin- ning of the Z93 subclade, and their actual TMRCA is expected to be 5000 years (or more) before the present. Most of the haplotype tree is L657 negative. An example of L657− base haplotype of an available 20-haplotype Asian branch in the 111 marker format is given below: 13 25 16 11 11 14 12 12 10 13 11 30—15 9 10 11 11 24 14 20 32 12 15 15 16—11 12 19 23 16 16 18 19 34 38 13 11—11 8 17 17 8 11 10 8 11 10 12 22 22 15 10 12 12 13 8 14 23 21 13 12 11 13 11 11 12 13—32 15 9 15 12 26 27 19 12 12 13 12 10 9 12 11 10 11 11 30 12 13 24 13 9 10 19 15 19 11 23 15 12 15 24 12 23 19 10 15 17 9 11 11 An example of L657+ base haplotype of an available Asian branch is as follows: 13 25 16 10 11 14 12 12 10 13 11 30—16 9 10 11 11 24 14 20 32 12 15 15 17—12 11 19 23 15 17 17 19 36 40 14 11—11 8 17 17 8 11 10 8 11 10 12 22 22 15 10 12 12 13 8 12 23 21 12 12 11 13 11 11 12 12 The first base haplotype is apparently ancestral to the branch, consisting of haplotypes from Turkey, Saudi Arabia, Qatar, the United Arab Emirates, Palestine, India, Kazakhstan, Iraq, Paki- stan, Poland, Germany, and one from Tatar Russia. 20 of 67- marker haplotypes in the branch contain 354 mutations, which place their common ancestor at 354/20/0.12 = 148 → 174 gene- rations, or 4350 ± 490 ybp. Fourteen 111-marker haplotypes in the branch contain 381 mutations, which place their common ancestor at 381/14/.198 = 139 → 162 generations, or 4050 ± 460 ybp. The second sub-branch is non-symmetrical, and contains six haplotypes from Saudi Arabia with a common ancestor of only 315 ± 110 ybp, and one very different haplotype from India, with an apparent common ancestor for all L657+ haplotypes of 2700 ybp. There are 16 mutations between the two L657+ and Map 3. The map of the South-Eastern branch, depicting birthplaces of the furthermost ancestors recorded i available databases. Magenta pins denote a parent Z93 branch, red and green pins correspond to Bash- rgyz branches, respectively. The rest of the branch (L342.2 positive) is marked by deep-blue n kir and Ky pins. The most represented Ashkenazy Jewish branch (not shown) covers densely an area of historical Polish-Lithuanian Commonwealth. Copyright © 2012 SciRes. 144  I. L. ROZHANSKII, A. A. KLYOSOV Figure 4. A haplotype tree of the South-Eastern branch (Z93), composed of 203 of 67-marker haplotypes. Practically all haplotypes (excep −) are assigned to L342.2+ subclade. There are two L- mIt ight relate to the appearance of the L342.2 subclade. 0 12 12 13 8 14 23 21 13 12 42.2) to 4800 years before the pre- se 4 with the 67 markers reported by FTDNA. Eight Kyrgyz ha xxx 10 12 12x 8 14 xx 12 12 11 xx 11 12 12 hich corre- sp 111-marker haplotype has the following alleles: 4 9 11 11 11 24 14 0 35 38 14 11—11 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 14 8 14 23 21 12 0 15 20 11 23 15 12 15 extended one among R1a1 haplotypes, both in num- ber of h of all Europea estern br t four L342.2 L657+ branches, of 7 and 17 haplotypes each. The tight, numerous young branch on the left is the Jewish R1a1 branch. The Bashkir (10 haplotypes) and Kyrgyz (7 haplotypes) branches are also shown. 657− branches in the 67-marker format, which places a com on ancestor of the two base haplotypes at 5300 years ago. m The R1a-L342.2 Bashkir branch of 10 haplotypes points to their common ancestor of about 1125 ± 190 ybp. Their 111- marker base haplotype is as follows: 13 24 16 11 11 15 12 12 12 13 11 31—15 9 10 11 11 24 14 20 31 12 15 15 15—11 11 19 23 16 15 18 19 35 39 14 11—11 8 17 17 8 12 10 8 11 10 10 22 22 15 1 11 13 11 11 12 13—33 15 9 15 11 26 28 19 11 12 1212 10 9 12 11 10 11 12 30 13 14 24 13 9 10 19 15 19 12 24 14 13 15 24 12 23 19 10 15 17 9 11 11 It has 28 mutations compared with the 111-marker predomi- nantly Middle Eastern L657− base haplotype, which places their common ancestor (L3 nt. The Kyrgyz branch is based mostly on the SMGF database (www.smgf.org), which employs another format, and shares only 3 plotypes are known in the 67- and 111-marker formats, but six of them belong to a rather recent lineage (650 ± 150 ybp), with possible origin in Altay. Since some alleles in the base haplotype of the entire branch are uncertain, they are marked by “x”: 13 25 16 1111 14 12 12 10 14 11 32—15 9 11 11 23 14 2131 12 1515 16—11 11 19 23 16xxxxx15 11—11 8 17 17 8 x 10 8 xx 12 A common ancestor of the Kyrgyz branch lived 2100 ± 250 ybp; this and the other branches of the L342.2 share a common ancestor within the range of 4400 ± 500 ybp, w onds to the ancestor of L342.2 itself. Finally, the large Jewish R1a-L342.2 branch with its base 12 11 13 10 11 12 13—32 15 9 17 12 27 27 19 12 12 12 12 10 9 12 11 10 11 11 30 12 12 25 13 9 10 2 25 13 25 16 10 11 14 12 12 10 13 11 30—1 20 30 12 12 15 15—11 11 19 23 14 16 19 2 12 23 19 10 15 17 9 11 11 This group descends from a common ancestor who lived only 1300 ± 150 ybp. It arose in Germany, but as part of a line- age with a common ancestor living 4475 ± 400 ybp, or what has been called the “Abraham times” (Klyosov & Rozhanskii, 2012). It is of interest to compare which base haplotype is closer to the Jewish base haplotype—the Middle Eastern or the Bashkir base haplotype. It would seem that the Middle Eastern haplo- type should be closer. Indeed, the mutational distances in the 111-marker format are 30 and 40, respectively. The common ancestor of the Jewish base and the Middle Eastern base, and of the Jewish base and the Bashkir base lived 4900 ± 500 and 4400 ± 500 ybp, respectively—or at the same time, within the margin of error. It fits well with the 4800 ybp obtained above for the L342.2 subclade. The common ancestor of all these L342.2 branches lived at the time of the Aryan migrations from Eastern Europe across the Russian Plain on their way to the east and south. Eurasian Branch (Z283), ~5500 ybp The Eurasian R1a branch Z283 includes haplotypes from Europe and from the Asian part of Russia which spreads to Pacific Ocean on the east. It is also found in low frequencies in Asia Minor and among ethnic Armenians. The Eurasian branch is the most aplotypes and in its territory. It embraces 86% n R1a1 haplotypes, while the Tenths (North-W anch) take only 12%. The rest are minor subclades, such as the Old Europeans. The Z93 subclade, with its mainly Asian haplotypes, encompasses 10% of the European R1a1 haplo- types, and it was not included in the above figures. Three coun- tries—Poland, Russia and Germany—embrace 55% of all the Eurasian branch, with the rest mainly in Ukraine, Belarus, the Czech Republic, Slovakia, Slovenia, Lithuania, Finland, Hun- gary, and Italy. R1a1a1g Z283 (Eurasian branch) R1a1a1g2 Z280 (Central Eurasian branch) R1a1a1g2a P278.2 (West Carpathian) R1a1a1g2b L365 (North European) R1a1a1g2c L366 (Central Eurasian-3) R1a1a1g2d Z92 (North Eurasian) R1a1a1g2d (North Carpathian) R1a1a1g2e L784 R1a1a1g3 Z284 (Scandinavian branch) R1a1a1g3a L448 (Young Scandinavian) R1a1a1g3a1 L176.1 (Scottish Highland) R1a1a1g3b Z287 (Old Scandinavian) R1a1a1g1 M458 (European branch) R1a1a1g1a Central European branch R1a1a1g1a1 L1029 R1a1a1g1b L260 (West Slavic branch) The aboveades of diagram shows the downstream subcl Copyright © 2012 SciRes. 145  I. L. ROZHANSKII, A. A. KLYOSOV Eurasian is the Central Eurasia which include al- most half followed first by the Scan and its subclades), with one-third 458 (and its ubclades)lo- trees are shown here not for a detailed analysis but for their overheir own rather distbranches which do not have ato Fig- ur Tpe bye timt pl 1111 12 13 The CPlain base hations separate the common3 → 34 generations, or 850 years, and their common ancestor lived (4600 + 3500 + branch Z283. The most populous one n branch (Z280) with its subclades, of all Eurasian (Z283) haplotypes. It is dinavian branch (Z284 of the Eurasian haplotypes; and then by M with 21% of the Eurasian haplotypes. A few haps types from Canada, Germany, Switzerland, England, Poland and Turkey have been proven to belong to the parent Z283 branch. Central Eurasian Z280 Subclade (4900 ± 500 ybp) Figure 5 shows the 67-marker 801-haplotype tree of the cen- tral Eurasian branch (Z280); Figure 6 presents the 111-marker tree of 96 haplotypes, currently available in this extended for- mat. The all shape. All four available SNPs have t inct branches on the trees, along with other n assigned SNP as yet (see legend es 5 and 6). Overall, there are 13 currently identified branches within subclade Z280. A common ancestor of Z280 lived 4900 ± 500 ybp, and had a base (ancestral) 111-marker haplotype as fol- lows: 13 25 16 11 11 14 12 12 10 13 11 30—15 9 10 11 11 24 14 20 32 12 15 15 16—11 12 19 23 16 16 18 19 35 38 14 11—11 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 14 23 21 12 12 11 13 11 11 12 13—32 15 9 15 12 26 27 19 12 12 12 12 10 9 12 11 10 11 11 30 12 13 24 13 9 10 19 15 20 11 23 15 12 15 24 12 23 19 10 15 17 9 11 11 Some of the Z280 subclade bearers can be assigned, at pre- sent, to a rather loose parent branch, the Central Eurasian group proper, descending directly from the above base haplotypes. Base haplotypes of the most populous branches are shown be- low, along with the timespans to their common ancestors. The Russian Plain (RP) Branch, or Central Eurasian-1 (CEA-1) Branch (4600 ± 500 ybp) 13 25 16 11 11 14 12 12 10 13 11 30—15 9 10 11 11 24 14 20 32 12 15 15 16—11 11 19 23 16 16 19 19 35 38 14 11—11 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 14 23 21 12 12 11 13 11 11 12 13 he RP base haplotype differs from the Z280 base haploty only two mutations (425 “lateral” years, that is an averag e required for these two mutations to occur), as marked; i aces their common ancestor at (4900 + 4600 + 425)/2 = 4960 ybp, which is Z280 itself (4900 ± 500 ybp) within the margin of error. Central Eurasian-2 (CEA-2) Branch (3500 ± 400 ybp) 13 25 16 11 11 14 12 12 10 13 11 30—15 9 10 11 11 24 14 20 31 12 15 15 16—11 1119 23 16 16 18 19 34 38 14 11—11 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 13 23 21 12 12 11 13 entral Eurasian-2 branch differs from the Russian plotype by 4 mutations (marked above). Four muta ancestors by 4/.12 = 3 Figure 5. A haplotype tree of the Central Eurasian branch (Z280), composed of 801 of 67-marker haplotypes. The North European (L365), Central Eurasian-3 (L366), West Carpathian (P278.2), and North Eurasian (Z92) branches have their identified SNPs. The tree contains other branches which do not have their SNPs identified as yet, and some of them are as follows, clock-wise: North Carpathian (NC), Balto-Carpathian-1 (BC- 1), West Eurasian-3 (WEA-3), Central Eurasian-1 (CEA-1), WEA-1, CEA-2, Eastern Carpathian (EC), BC-3, BC-2. Figure 6. A haplotype tree of the Central Eurasian branch (Z280), composed of 96 of 111-marker haplotypes. The North European (L365), West Car- pathian (P278.2), and North Eurasian (Z92) branches are indicated, as well as (clock-wise): Balto-Carpathian (BC), North Carpathian (NC), Eastern Carpathian (EC). 850)/2 = 4475 ybp. This is the time of the Russian Plain com mon ancestor (4600 ± 500 ybp) within the margin of error. - Copyright © 2012 SciRes. 146  I. L. ROZHANSKII, A. A. KLYOSOV Copyright © 2012 SciRes. 147 entral Eurasian-3 (CEA-3) Branch 12 22 22 15 10 12 12 12 8 13 23 21 12 (RP branch)stor at ~4825 th the Russian Plain (RP) an (Z280) common an 100 ± 500 ybp) type by three mutations (650 years), hence, their common ithin the margin of-branch (WEA-1) with a com0 ybp. It devi- at ). 19 23 16 16 17 20 36 39 12 11—11 8 17 17 8 12 12 11 13 The base haplotyp280 by 8 mutations, w 14 11—11 8 17 17 83 21 12 12 11 13 Twelve mutations base Z280 haplo- ty arpathian (BC) Branch (4300 ± 500 ybp) Eastern Carpathian (EC) Branch (2600 ± 300 ybp) 13 25 16 11 11 14 12 12 10 13 11 30—15 9 10 11 11 24 14 20 32 12 15 15 16—11 11 C (2500 ± 350 ybp) 13 25 16 11 11 14 12 12 10 13 11 30—15 9 10 11 11 24 14 20 31 13 15 15 16—11 12 19 22 16 16 18 19 33 39 13 11—11 17 17 8 12 10 8 11 10 12 10 8 10 10 12 22 22 15 10 12 12 13 8 14 23 21 1111 12 13 e differs from that of Z 8 12 11 13 11 11 12 13 The Central Eurasian-3 is a minor branch, characterized by SNP L366. It differs from the parent base haplotype hich corresponds to 1800 years, and translates to a common ancestor at 4650 ybp. Again, it is well within margin of error for TMRCA of the entire Central Eurasian branch. by 11 mutations. This places the common ance ybp, which is within the margin of error for bo and the Central EurasiWestern Carpathian (WC) Branch (2600 ± 300 ybp) 13 25 15 10 11 15 12 12 11 13 11 29—15 9 10 11 11 24 14 20 31 12 15 15 16—11 12 19 23 17 16 17 19 35 39 cestors. Western Eurasian (WEA) Branch (4 12 10 8 11 11 12 22 22 15 10 12 12 13 8 13 2 12 11 12 13 (2775 years) from the 13 25 16 11 11 14 12 12 11 13 11 30—15 9 10 11 11 24 14 20 33 12 1515 16—11 12 19 23 16 16 1819 34 38 14 11—11 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 14 23 21 12 12 11 13 1111 12 13 The Western Eurasian base haplotype differs from the Z280 base haplo pe place the common ancestor of the Western Carpathian branch at 5100 ybp. Its assignment is also confirmed by SNP P278.2. The above branches are spread almost uniformly over most of ancestor lived 4825 ybp. This is Z280 itself w error. The WEA branch has a distinct sub mon ancestor of 2300 ± 30 Central and Eastern Europe; however, they are virtually ab- sent in Scandinavia (Map 4). Balto-C es by 10 mutations from the parent WEA base haplotype, or by 2275 “lateral” years. This places their common ancestor with the parent branch at 4300 ybp, i.e., the WEA parent branch itself (4100 ± 500 ybp 13 25 16 10 11 14 12 12 10 13 11 30—15 9 10 11 11 24 14 20 32 13 15 15 16—11 12 19 23 16 16 18 18 34 37 13 11—11 Map 4. The map of the parent Central Eurasian, Western Eurasian, Eastern and Western Carpathian branches, de- picting birthplaces of the furthermost ancestors recorded in available databases. Yellow pins denote the par- ent branch, pins of other colors correspond to junior branches.  I. L. ROZHANSKII, A. A. KLYOSOV 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 14 23 21 12 12 11 13 11 11 12 13 The Balto-Carpathian branch deviates by 6 mutations (1325 “lateral” years) from the Z280 base haplotype. The BC branch has two distinct sub-branches (BC-1 and BC-2) with common ancestors of 2550 ± 300 and 2200 ± 250 ybp, respectively. They deviate by 4 and 9 mutations from the parent (BC) base haplotype, or by 850 and 2025 “lateral” years respectively. This places their common ancestor with the parent branch at 3850 and 4300 ybp, or the BC parent branch itself (4300 ± 500 ybp). In contrast to the “core” branches of Z280, the Balto-Car- pathian shows distinct geographical distribution, as reflected by its name (Map 5). Its BC-2 sub-branch is characteristic for Slovenians and Croatians, being their “trademark” genealogical lineage. Northern Eurasian (NEA) Branch (4450 ± 450 ybp) This branch is characterized by SNP Z92. It consists of two major sub-branches. The older branch (NEA-1) (3600 ± 400 ybp) has the following base haplotype: 13 25 16 11 11 14 12 12 11 13 11 30—15 9 10 11 11 24 14 20 33 12 14 15 16—11 12 19 23 16 16 18 18 34 40 14 11—11 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 13 23 22 12 12 11 13 11 11 12 13 Its base haplotype deviates from that of Z280 by 9 mutations (2025 years), which places their common ancestor at 5260 ybp, within the margin The base haplo 300 ybp) is: 13 25 16 11 11 15 12 12 10 13 11 30—15 9 10 11 10 25 14 20 32 12 14 14 16—12 12 19 23 15 16 18 20 34 38 13 11—12 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 13 23 22 12 12 11 13 11 11 12 13 It has 12 mutations (2775 years) from the base Z280 haplo- type and 13 mutations (3025 years) from the NEA-1 base hap- lotype. The common ancestor of these two sub-branches of Z92 lived 4450 ± 450 ybp. Present-day bearers of both NEA branches populate Lithua- nia, Belarus, and the north-eastern part of Poland (Map 6). It is the most widespread genealogical lineage among ethnic Rus- sians. Northern European (NE) Branch (2600 ± 350 ybp) 13 25 15 11 11 14 12 12 10 13 11 30—17 9 10 11 11 23 14 20 33 13 15 15 16—11 11 19 23 16 15 18 19 34 38 14 11—11 8 17 17 8 12 10 8 11 10 12 21 22 16 10 12 12 13 8 14 24 21 13 12 11 13 1111 12 13 The Northern European base haplotype differs by 13 muta- tions (3025 “lateral” years) from that of Z280, and is inde- pendently defined by SNP L365. It again places its common ancestor at 5260 ybp, which is Z280 itself. Northern Carpathian (NC) Branch (2150 ± 300 ybp) 9 10 11 11 24 14 5 40 14 11—11 3 8 14 23 21 12 of error from 4900 ± 500 ybp for Z280 itself. type of the younger branch (NEA-2) (2350 ± 13 25 17 11 11 14 12 12 10 13 11 31—14 20 32 13 15 15 16—11 12 19 23 16 16 18 19 3 9 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 1 Map 5. The map of the Balto-Carpathian branch, depicting birthplaces of the furthermost ancestors recorded in branch, deep-blue and green pins correspond to BC-1 available databases. Yellow pins denote the parent and BC-2 sub-branches, respectively. Copyright © 2012 SciRes. 148  I. L. ROZHANSKII, A. A. KLYOSOV Map 6. The map of the Northern Eurasian branch, depicting birthplaces of the furthermost ancestors recorded in available databases. Yellow pins denote the older branch (NEA-1), magenta pins correspond to NEA-2. 12 11 13 11 11 12 13 This base haplotype differs by 7 mutations (1550 years) from the base Z280 haplotype. There is also a minor branch, which is defined by subclade L784 (2500 ± 450 ybp): 13 24 17 11 11 14 12 12 10 14 11 30—15 9 10 11 11 24 14 20 31 13 15 15 16—11 12 19 23 16 17 17 19 34 39 12 12—11 8 17 17 8 12 10 8 11 10 12 22 22 15 10 12 12 13 8 14 23 21 12 12 11 13 11 11 12 13 The latter branches seem to be geographically limited to pre- sent day Poland and Eastern Germany, and—in lower frequent- cies—the Baltic countries, Ukraine, and Russia (Map 7). Overall Z28 The eight base Plain, West Eurasian, Balto-Carpathian, North European, East Carpathian, North Eurasian, North Carpathian, and West Car- pathian) differ among themselves by 68 round-up or 64 full mutations (some mutations are fractional), resulting in the Z280 base haplotype, listed above. It gives 64/8/.12 = 67 → 72 gene- rations, or 1800 years down from their average “age” (3100 years). This results in 4900 years from their common ancestor, which is Z280 itself (4900 ± 500 ybp, see above). European Subclade M458 (4200 ± 450 ybp) This subclade consists of two principal branches, the Central European (CE) and West Slavic (WS). A division between them is clearly seen on their haplotype tree (Figure 7). The CE branch, in fact, consists of two sub-branches, both arose 2900 ± 400 ybp. One of them (CE-1) has the following base haplotype: 13 25 16 10 11 14 12 12 11 13 11 29—16 9 10 11 11 23 14 20 32 12 15 15 16—11 11 19 23 17 16 18 19 34 38 14 11—11 8 17 17 8 11 10 8 12 10 12 21 22 15 10 12 12 13 8 14 25 21 13 12 11 13 11 11 12 13 It differs by 14 mutations (3325 years) from the Z280 base haplotype (see above), which places their common ancestors at ≥5600 ybp. The “equal or higher” sign here indicates that the difference between them cannot be less that the “age” of sub- clades Z280 and M458. The second branch (CE-2) differs from CE-1 in only two loci, having there three mutations: a multi-copyuplication 12-15- 15-16 → 12-12-15-15-15-16 in DYS464 (presumably a one- in CDYb. These E-1 and CE-2 at 3100 ybp, if both branches arose independently from their com- mon ancestor. It might be thought that CE-2 is a CE-1 down- stream branch, which arose immediately after CE-1, within the indicated margin of error in their dating. This is likely, since even the 67 marker haplotype tree cannot cleanly separate the CE branches (see Figure 7). Their haplotypes are nearly iden- tical statistically, except that they descended from slightly dif- ferent ancestral (base) haplotypes. Also, there is a downstream subclade L1029 within the CE branch which embraces the most of bearers of both the sub-branches. Apparently, it arose at the same time as the CE branch itself. In contrast, the Western Slavic branch (L260) is quite distinct. It does not include any CE haplotype on the tree (Figure 7). It arose 2700 ± 300 ybp, and its base haplotype is as follows: 13 25 17 10 10 14 12 12 10 13 11 30—16 9 10 11 11 23 14 20 31 12 15 16 16—11 11 19 23 16 16 18 19 34 39 13 11—11 0 Base Haplotype and Its “Age” haplotypes of the main branches (Russian time event), and a two-step mutation 38 → 40 three mutations place a common ancestor of C d Copyright © 2012 SciRes. 149  I. L. ROZHANSKII, A. A. KLYOSOV Map 7. The map of the Northern European (green pins) and Northern Carpathian (magenta pins) branches, depict- ing birthplaces of the furthermost ancestors recorded in available databases. Blue pins denote the minor branch, which is characterized by SNP L784. Figure 7. A 67-marker haplotype tree of the R1a-M458 subclade, composed of 352 haplotypes. The Central European (CE) branch takes the right-hand side and the upper left-hand side. The distinct Western Slavic (WS) branch is on the left. 8 1 12 1 2 muta- tio Slavic branch, which makes the M458 subclade 4200 years the Z280 base haplotypes (1 7 17 8 12 10 8 12 10 12 22 22 15 10 12 12 13 8 14 25 21 12 1 13 11 11 12 13 It differs by 14 mutations (3325 years) from the Z280 base haplotypes, which places their common ancestor at the same ≥5600 years (see the explanation above). There are 1 ns between the CE-1 base haplotype and that of the Western ed on available haplotypes. “old” at least bas The European subclade M458 shows nearly the same geo- graphical distribution as the Central Eurasian Z280, but statis- tically it is more densely represented on its western edge (see also Underhill et al., 2009), reaching up to 70 % of all R1a1 bearers in the Czech Republic (Map 8). On the other hand, it is relatively rare among ethnic Lithuanians and Slovenians, who belong predominantly to Z280 subclade. Scandinavian Subclade Z284 (4300 ± 500 ybp) This subclade includes four principal branches, the Old Scan- dinavian (OS, Z287/Z288, which arose 3700 ± 400 ybp), Young Scandinavian (YS, L448, which arose 2700 ± 350 ybp), the Scottish Highland (SH, L176.1, which arose 1850 ± 290 ybp), and the parent branch, which is negative to the known downstream SNPs (which arose 4300 ± 500 ybp). Additionally, the R1a nomenclature also includes the L175 subclade as downstream of L176.1, however, it is most proba- bly a private SNP. The haplotype tree is shown in Figure 8. The lower part of the tree is represented by the Old Scandi- navian branch with its base haplotype as follows: 13 25 16 10 11 14 12 12 10 13 11 30—15 9 10 11 11 24 14 20 31 12 15 15 16—11 12 19 23 16 16 18 19 35 39 12 11—12 8 17 17 8 12 10 8 11 10 12 22 22 16 11 12 12 13 8 14 23 21 12 12 11 13 11 11 12 13 It differs by 8 mutations from 800 years), which places their common ancestor at ≥5200 years (see the explanation above regarding the “equal or higher” sign). Since there are 17 identical (base) 12-marker haplotypes Copyright © 2012 SciRes. 150  I. L. ROZHANSKII, A. A. KLYOSOV Map 8. The map of the M458 subclade, depicting birthplaces of the furthermost ancestors recorded in available da- tabases. Blue pins denote the Central European branch (CE-1 deep-blue, CE-2 light-blue), green pins corre- spond to the West Slavic branch. Figure 8. A 67-marker haplotype tree of the R1a-Z284 (Scandinavian) subclade, composed of 540 haplotypes. Three principal subclades/branches are shown: the Old Scandinavian (Z284), the Young Scandinavian (L448), and the Scottish Highland (L176.1). among 178 OS haplotypes in the dataset, the logarithmic me- thod gives [ln(178/17)]/.12 = 117 → 133 generations, or ~3325 ye 15 9 10 11 11 23 14 20 32 12 15 15 16—11 12 19 21 16 16 17 18 34 38 12 11—11 8 12 11 13 11—12 8 s to their common ancestor. The com- pa ars to the common ancestor; this is within the above 3500 ± 400 ybp, as determined by the linear method. The good fit (within 5%) means that practically all 178 OS haplotypes actu- ally descended from one common ancestor (Klyosov, 2009a). The Young Scandinavian branch (L448+) has the following base haplotype: 13 25 15 11 11 14 12 12 10 14 11 31— 17 17 8 12 10 8 11 10 12 22 22 15 11 12 12 13 8 14 23 21 12 11 11 12 12 Since the YS base haplotype is younger than the OS base, it is more distant from the Z280 base haplotype on the same hap- logroup tree; indeed, they deviate by 12 mutations (2775 years); this places their common ancestor at ≥5300 ybp, which is prac- tically identical with the date obtained above with the OS base haplotype. The same figure is reproduced with a rather loose set of hap- lotypes, positive at Z284 and negative at L448 and Z287/Z288 (4200 ± 500 ybp), with the base haplotype as follows: 13 25 15 10 11 14 12 12 10 13 11 30—15 9 10 11 11 24 14 20 32 12 15 15 16—11 11 19 23 16 16 17 18 35 39 12 17 17 8 12 10 8 11 10 12 22 22 15 11 12 12 13 8 14 23 21 12 12 11 13 11 11 12 13 Its 10-mutation difference (2275 years) with the base Z280 results in ≥5700 year rison of the above base haplotype with those for OS and YS suggests that it is identical or nearly identical to the base hap- lotype of the entire Z284 subclade, with its estimated “age” of Copyright © 2012 SciRes. 151  I. L. ROZHANSKII, A. A. KLYOSOV ~4300 years. 13 25 16 11 11 14 12 12 10 14 11 31—15 8 10 11 11 23 14 20 29 12 15 15 15—10 12 19 21 16 16 17 19 34 38 12 11—11 8 17 17 8 12 10 8 11 10 12 22 22 15 11 12 12 13 8 14 23 21 12 12 11 13 11 11 12 12 Seven mutations (1550 years) with the YS base haplotype confirms their relative position of the subclade tree. The known sub-branches of Z284 were found mostly in Scandinavian countries and on the Isles (Map 9). Norwegians and Swedish-speaking Finns seem to be the typical bearers of this genealogical DNA-lineage. Chronology of the Main M417+ Subclades The combined data of SNP mutations and extended STR haplotypes allowed us to make a step-wise reconstruction of the base haplotypes of the major subclades of the R1a1 haplogroup. A consensus tree was calculated from these data (Figure 9). Its topology follows closely that of SNP phylogeny, while the dates of its principal nodes are quite robust, as it is shown by the examples above. Conclusion and Some Historical Considerations It follows from the data presented in this study, that all three principal downstream R1a subclades, L664, Z283 and Z93, split from their common European ancestor at about the same time, around 7000 - 6000 ybp, and that all three went through a population bottleneck, and then expanded 1000 - 1500 years L664 apparently stayed in North-Western Europe; its lineage recovered and began expanding ~4575 ybp. The Z280 subclade (descendants of Z283) moved east to the Russian Plain ~4800 - 4600 ybp when the Arbins, bearers of haplogroup R1b, were arriving in Europe from different direc- tions—from North Africa to the Pyrenees (4800 ybp); then—as the Bell Beakers—to the continent; and finally from Asia Mi- nor and the Middle East to the Apennines and the Balkans (4800 - 4500 ybp). The Z280 subclade and its Russian Plain branch formed at least 16 sub-branches from the middle of the 3rd millennium BC and later in the course of the westward re- population of Europe in the 1st millennium BC-1st millennium CE. The Z93 subclade began to expand during the Aryan migra- tions, on their way to India and the Middle East in the 3rd-2nd millennia BC. The eastward migration of the Aryans continued to the east (the Ural Mountains and beyond, to the Altay), south east (Iran and Hindustan), south (Anatolia, Mitanni and further south to the Middle East) between 4500 and 3500 ybp. The Jewish R1a common ancestor lived 4525 ± 400 ybp; his de- scendants also almost vanished, but their long population bot- tleneck ended around the 8th century CE, in the Diaspora, ap- parently in Germany. During the 1st millennium BC bearers of R1a started to re- populate Europe west of the Carpathian Mountains. This led to the appearance of the Northern European branch (L365, 2600 ybp), Western Carpathian (P278.2, 2600 ybp), Eastern Carpa- thian (2600 ybp, SNP is unknown), Northernpathian (2150 ybp, SNP is unknown), two Balto-Carpathian branches (2550 The haplotype of the Scottish Highland branch (L176.1) is later. Car Map 9. The map of the Scandinavian Z284 subclade, depicting available databases. Magenta pins denote the parent br ond birthplaces of the furthermost ancestors recorded in anch, green pins mark the Old Scandinavian one to the young Scandinavi(Z287/Z288), deep-blue and light-blue pins corresp Scottish Highland sub-branch (L176.1), respectively. an branch (L448) and its Copyright © 2012 SciRes. 152 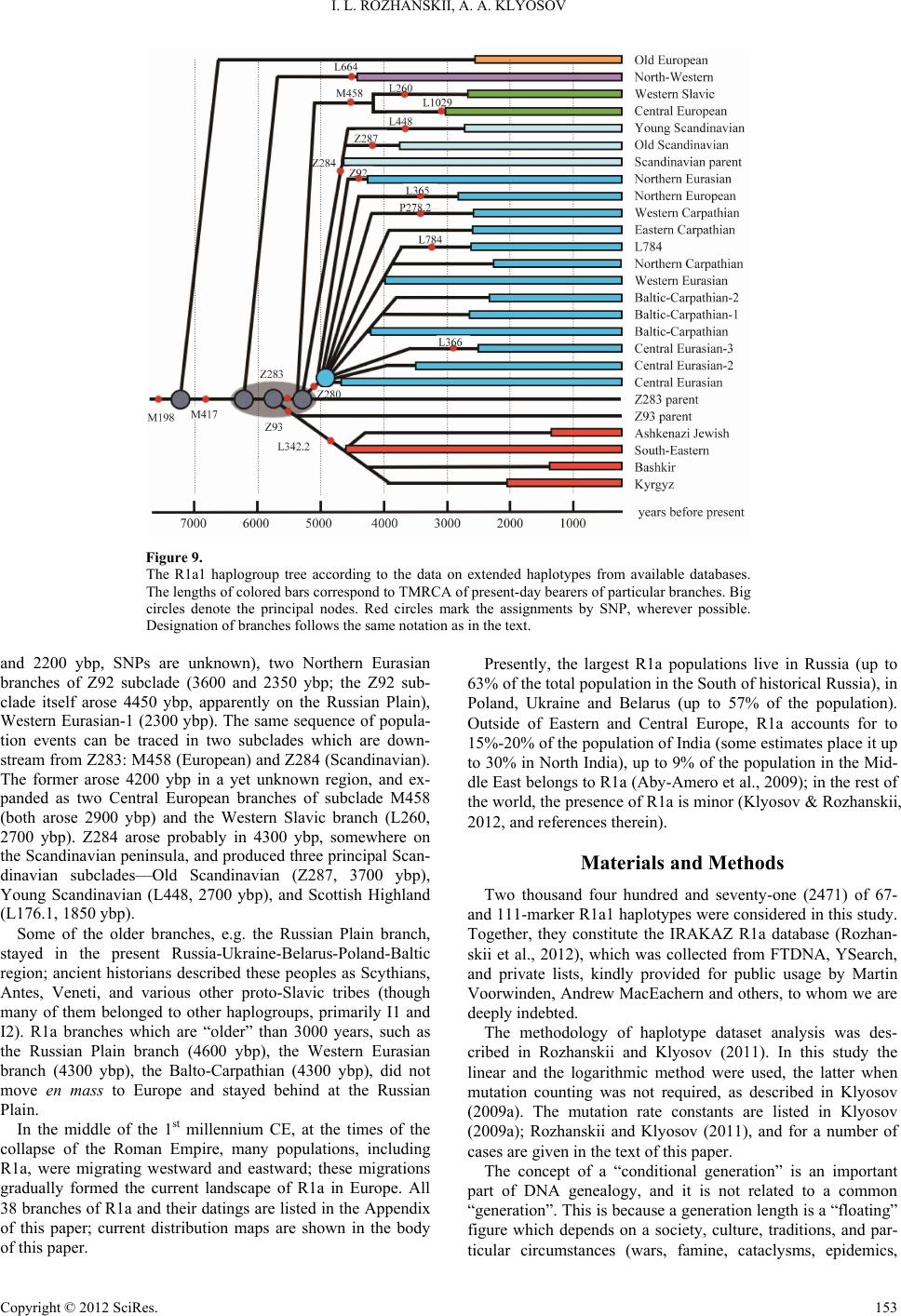 I. L. ROZHANSKII, A. A. KLYOSOV extended haplotypes from available databases. f present-day bearers of particular branches. Big k the assignments by SNP, wherever possible. in the text. Figure 9. a on A o mar as estern Eurasian-1 (2300 ybp). The same sequence of popula- tion events can be traced in two subclades which are down- stream from Z283: M458 (European) and Z284 (Scandinavian). The former arose 4200 ybp in a yet unknown region, and ex- panded as two Central European branches of subclade M458 (both arose 2900 ybp) and the Western Slavic branch (L260, 2700 ybp). Z284 arose probably in 4300 ybp, somewhere on the Scandinavian peninsula, and produced three principal Scan- dinavian subclades—Old Scandinavian (Z287, 3700 ybp), Young Scandinavian (L448, 2700 ybp), and Scottish Highland (L176.1, 1850 ybp). Some of the older branches, e.g. the Russian Plain branch, stayed in the present Russia-Ukraine-Belarus-Poland-Baltic region; ancient historians described these peoples as Scythians, Antes, Veneti, and various other proto-Slavic tribes (though many of them belonged to other haplogroups, primarily I1 and I2). R1a branches which are “older” than 3000 years, such as the Russian Plain branch (4600 ybp), the Western Eurasian branch (4300 ybp), the Balto-Carpathian (4300 ybp), did not move en mass to Europe and stayed behind at the Russian Plain. In the middle of the 1st millennium CE, at the times of the collapse of the Roman Empire, many populations, including R1a, were migrating westward and eastward; these migrations gradually formed the current landscape of R1a in Europe. All 38 branches of R1a and their datings are listed in the Appendix of this paper; current distribution maps are shown in the body of this paper. Presently, the largest R1a populations live in Russia (up to 63% of the total population in the South of historical Russia), in Poland, Ukraine and Belarus (up to 57% of the population). Outside of Eastern and Central Europe, R1a accounts for to 15%-20% of the population of India (some estimates place it up to 30% in North India), up to 9% of the population in the Mid- dle East belongs to R1a (Aby-Amero et al., 2009); in the rest of the world, the presence of R1a is minor (Klyosov & Rozhanskii, 2012, and references therein). Materials and Methods Two thousand four hundred and seventy-one (2471) of 67- and 111-marker R1a1 haplotypes were considered in this study. Together, they constitute the IRAKAZ R1a database (Rozhan- skii et al., 2012), which was collected from FTDNA, YSearch, and private lists, kindly provided for public usage by Martin Voorwinden, Andrew MacEachern and others, to whom we are deeply indebted. The methodology of haplotype dataset analysis was des- cribed in Rozhanskii and Klyosov (2011). In this study the linear and the logarithmic method were used, the latter when mutation counting was not required, as described in Klyosov (2009a). The mutation rate constants are listed in Klyosov (2009a); Rozhanskii and Klyosov (2011), and for a number of cases are given in the text of this paper. The concept of a “conditional generation” is an important part of DNA genealogy, and it is not related to a common “generation”. This is because a generation length is a “floating” figure which depends on a society, culture, traditions, and par- ticular circumstances (wars, famine, cataclysms, epidemics, The R1a1 haplogroup tree according to the dat The lengths of colored bars correspond to TMRC circles denote the principal nodes. Red circles Designation of branches follows the same notation and 2200 ybp, SNPs are unknown), two Northern Eurasian branches of Z92 subclade (3600 and 2350 ybp; the Z92 sub- lade itself arose 4450 ybp, apparently on the Russian Plain), c W Copyright © 2012 SciRes. 153  I. L. ROZHANSKII, A. A. KLYOSOV etc.). It means thions of “generations” into years should be done b The calibration wa in the preceding p skii & Klyosov, 2011). As it follows from basic rules of chemical kinetics, the ex- perimentally determined ratio n/N in the equation n/N = kt ultimately determines a product kt. In this equation: n = a number of mutations (from the base haplotype) in a se- ries of N haplotypes, N = a number of haplotypes, descending from one common ancestor (having the base haplotype) in the series, k = the average mutation rate constant for the haplotypes (per generation) of the given format, t = a number of generations (TMRCA). Therefore, if we set the length of the generation, for example as 25 years (making it “the “conditional generation”, which is a mathematical value rather that a floating “generation”), then the value of k can be calibrated with respect to a certain timespan during which n mutations occurred per N haplotypes. For ex- ample, in 84 of 67 marker haplotypes of the Clan Donald (the common ancestor of which, John, Lord of the Isles, died in 1386, with the TMRCA equaling to 650 years before present, which is 26 “conditional generations” of 25 years each), there were 262 mutations. This gives 262/84 = 26 k, and hence k = .12 mutations per the conditional generation (25 years) per the 67 marker haplotype. As it is shown inii & Klyosov, 2011), the ob- tained mutation rate ducible for many ser haplogroups from ma ct, it is reproducib lective number of mutations in the dataset having one common ogy is the loga- ation counting in haplotypes. It is based on the assumption (which largely or practically always holds true) that a transition of the base hap- lotypes into mutated ones (or, rather, belonging to descendants inherited mutated haplotypes) is described by the first-order kinetics: N = Aekt that is ln(N/A) = kt where: N = a total number of haplotypes in a set, A = a number of unchanged (identical, not mutated) base haplotypes in the set, k = an average mutation rate, t = a number of generations to a common ancestor. One can see the logarithmic method does not consider a number of mutations in haplotypes; only mutated and non- mutated (base) haplotypes are considered. The above equation is the fundamental one in chemical ki- netics. Radioactive decay, for example, is described by the same equation, though it does not involve an analogy with “back mutations”, since it is truly irreversible. Back mutations (or reverse mutations), though, are quite common in haplotypes, and their analysis and corrections for back mutations are given in (Klyosov, 2009a; Rozhanskii & Klyosov, 2011). Haplotype trees were composed using HYLIP, the Phylog- eny Inference Package program (Felsenstein, J., 2004; for mul- ee Klyosov, 2009a, for back mutations 2009a); Rozhanskii 011). Margins of error were calculated as de- scribed in Klyosov (2009a). at convers y calibrating using known historical events. s performed on many examples, as described aper in Advances in Anthropology (Rozhan- ancestor. Another principal method in DNA geneal rithmic method, which does not involve mut (Rozhansk constant of .12 is rather accurately repro- ies of 67 marker haplotypes from many ny tested populations around the world. In le for ANY tested population in the world tiple examples of usage and calculations s 2009b and references therein). Corrections were introduced as described in Klyosov ( and Klyosov (2 fa for which 67 marker haplotypes are available (Rozhanskii & Klyosov, 2011). This “calibration” connects a number of muta- tions in a series of haplotypes (in a given format and length, from the 6 marker haplotypes to the 111 marker haplotypes) to a known number of years to a known historical event or a known genealogy, setting a length of generation as 25 years, and verifying the mutation rate constant related to this “condi- tional generation”. It should be noted that we could have set the timespan to the common ancestor of the Clan Donald as 13 generations of 50 years each and it would not have changed a thing. There would still be 650 years to the common ancestor. The mutation rate constant would simply be twice as high. The mutation rate constant for the 111-marker haplotypes, also employed in this study, equals to .198 mutation per haplo- type per conditional generation (Rozhanskii & Klyosov, 2011). The above equation is essentially the basis for the “linear” method in DNA genealogy. It is based on mutation counting in haplotypes in the dataset compared with the “base” haplotype, which is (or the best available approximation to) the ancestral haplotype for the dataset. The “base” haplotype is the ancestral haplotype in an ideal case. However, since those base haplo- types often are deduced ones, it would be inaccurate to call them “ancestral” when that might not necessarily be true. Hence, we call them “base” haplotypes. Base haplotypes in the dataset were determined by minimization of mutations; by de- finition, the base haplotype is one which has the minimum col- Acknowledgements The authors are indebted to Dr. Judith Remy Leder for her va P luable help with the preparation of the manuscript. REFERENCES Abu-Amero, K. K., Hellani, A., Gonzalez, A. M., Larruga, J. M., Cabrera, V. M., & Underhill, P. A. (2009). Saudi Arabian Y-chro- mosome diversity and its relationship with nearby regions. BMC Genetics, 10, 59. doi:10.1186/1471-2156-10-59 Balanovsky, O., Rootsi, S., Pshenichnov, A., Kivisild, T., Churnosov, M., Evseeva, I., Pocheshkhova, E. et al. (2008). Two sources of the Russian patrilineal heritage in their Eurasian context. American Journal of Hum doi:10.1016/j.ajhg. an Genetics, 82, 236-250. 2007.09.019 Behar, D. M., Thomas, M. G., Skorecki, K., Hammer, M. F., Bulygina, E., Rosengarten, D., Jones, A. L. et al. (2003). Multiple origins of Ashkenazi Levites: Y chromosome evidence for both Near Eastern and European Ancestries. American Journal of Human Genetics, 73, 768-779. doi:10.1086/378506 Childe, G. (1926). The Aryans: A study of Indo-European origins. Dorchester: Dorset Press. Cinnioglu, C., King, R., Kivisild, T., Kalfoglu, E., Atasoy, S., Cavalleri, G. L., Lillie, A. S. et al. (2004). Excavating Y-chromosome haplo- type strata in Anatolia. Human Ge ne ti c s, 114, 127-148. doi:10.1007/s00439-003-1031-4 Felsenstein, J. (2004). PHYLIP (Phylogeny Inference Package). Ver- Copyright © 2012 SciRes. 154  I. L. ROZHANSKII, A. A. KLYOSOV Copyright © 2012 SciRes. 155 sion 3.6. Seattle: Department of Genome Sciences, University of Washington. Gamkrelidze, T. V., & Ivanov, V. V. (1995). Trends in linguistics 80: Indo-European an d th e Indo-Europeans. Berlin: Mouton de Gruyter. Gray, R. D., & Atkinson, Q. D. (2003). Language-tree divergence times support the Anatolian theory of Indo-European origin. Nature, 426, 435-439. doi:10.1038/nature02029 Haak, W., Brandt, G., de Jong, H. N., Meyer, C., Ganslmeier, R., Heyd, V., Hawkesworth, C. et al. (2008). Ancient DNA strontium isotopes, and osteological analyses shed light on social and kinship organiza- tion of the Later Stone Age. Proceedings of the National Academy of Sciences of the United States of America, 105, 18226-18231. doi:10.1073/pnas.0807592105 Kayser, M., Lao, O., Anslinger, K., Augustin, C., Bargel, G., Edelmann, J., Elias, S. et al. (2005). Significant genetic differentiation between Poland and Germany follows present-day political borders, as re- vealed by Y-chromosome analysis. Human Genetics, 117, 428-443. doi:10.1007/s00439-005-1333-9 Keyser, C., Bouakaze, C., Crubezy, E., Nikolaev, V. G., Montagnon, D., Reis, T., & Ludes, B. (200t DNA provides new insight an people. Human Genetics, 0683-0 9). Ancien into the history of south Siberian Kurg 0.1007/s00439-009-126, 395-410. doi:1 K Eastern Ukrainians as inferred from the Y- harkov, V. N., Stepanov, V. A., Borinskaya, S. A., Kozhekbaeva, Zh. M., Gusar, V. A., Grechanina, E. Ya., Puzyrev, V. P. et al. (2004). Gene pool structure of chromosome haplogroups. Russian Journal of Genetics, 40, 326-331. doi:10.1023/B:RUGE.0000021635.80528.2f K ans. Russian Journal of harkov, V. N., Stepanov, V. A., Feshchenko, S. P., Borinskaya, S. A., Yankovsky, N. K., & Puzyrev, V. P. (2005). Frequencies of Y- chromosome binary haplogroups in Belarussi Genetics, 41, 928-931. doi:10.1007/s11177-005-0182-x Klyosov, A. A. (2009a). DNA Genealogy, mutation rates, and some historical evidences written in Y-chromosome. I. Basic principles and the method. Journal of Genetic Geneal ogy, 5, 186-216. Klyosov, A. A. (2009b). DNA Genealogy, mutation rates, and some historical evidences written in Y-chromosome. II. Walking the map. Journal of Genetic Genealogy, 5, 217-256. Klyosov, A. A. (2010a). DNA lineages and origin of the “Tenths”, the North-Western European subfamily R1a1 with YS388 = 10. Pro- ceedings of the Russian Academy o f DNA Genealogy, 2, 983-1028. Klyosov, A. A. (2010b). DNA lineages and origin of the Tenths, the R1a1 subfamily with DYS388 = 10: The story in progress. Proceed- ings of the Russian Academy of DNA Genealogy, 3, 2205-2217. Klyosov, A. A. (2011). Biological chemistry as a foundation of DNA genealogy: The emergence of “molecular history”. Biokhimiya (Eng- lish translation), 76, 517-533. Klyosov, A. A. (2012). An emerging pattern of L664 SNP mutation in the Tenths (bearers of DYS388=10), North-Western branch of R1a1 haplotypes. Proceedings of the Russian Academy of DNA Genealogy, 5, 30-37. Klyosov, A. A., & Rozhanskii, I. L. (2012). Haplogroup R1a as the Proto Indo-Europeans and the legendary Aryans as witnessed by the DNA of their current descendants. Advances in Anthropology, 2, 1- 13. doi:10.4236/aa.2012.21001 Li, C., Li, H., Cui, Y., Xie, C., Cai, D., Li, W., Mair, V. H. et al. (2010). Evidence that a West-East admixed population lived in the Tarim Basin as early as the early Bronze Age. BMC Biology, 8, 15. doi:10.1186/1741-7007-8-15 Renfrew, C. (2000). 10,000 or 5000 year ago? Questions on time depth. In Renfrew, C., McMahon, & A., Trask, L. (Eds.), Time depth in historical linguistics (pp. 413-439). Cambridge: The McDonald In- stitute for Archaeological Research. Rozhanskii, I. L., & Klyosov, A. A. (2011). Mutation rate constants in DNA genealogy (Y chromosome). Advances in Anthropology, 1, 26- 34. doi:10.4236/aa.2011.12005 & ZolotareRozhanskii, databa I., Klyosov, A., v, A. (2012). IRAKAZ R1a se. Proceedings of the Academy of DNA Genealogy, 5, 553- 559. , S., Zhivotovsky, L. A., King, R., Mehdi, S. Q., Edmonds, C. arity and temporality s. Human Genetics, 78, 202-221. Sengupta A., Chow, C. E. T., Lin, A. A. et al. (2006). Pol of high-resolution Y-chromosome distributions in India identify both indigenous and exogenous expansions and reveal minor genetic in- fluence of central Asian pastoralist doi:10.1086/499411 Sh, Bhat, arma, S., Rai, E., Sharma, P., Jena, M., Singh, S., Darvishi, K. A. K. et al. (2009). The Indian origin of paternal haplogroup R1a1* substantiates the autochtonous origin of Brahmns and the caste sys- tem. Journal of Human Genetics, 54, 47-55. doi:10.1038/jhg.2008.2 Underhill, P. A., Myres, N. M., Rootsi, S., Metspalu, M., Zhivotovsky, M. A., King, R. J. et al. (2009). Separating the post-Glacial coances- try of European and Asian Y chromosomes within haplogroup R1a. European Journal of H um an Ge ne ti cs , 18, 479-484. doi:10.1038/ejhg.2009.194 Wells, R. S., Yuldasheva, N., Ruzibakiev, R., Underhill, P. A., Evseeva, I., Blue-Smith, J., Jin, L. et al. (2001). The Eurasian Heartland: A continental perspective on Y-chromosome diversity. Proceedings of the National Academy of Sciences of the United States of America, 98, 10244-10249. doi:10.1073/pnas.171305098  I. L. ROZHANSKII, A. A. KLYOSOV Appendix 1 list of R1a branches in Europe (dates in ybp). A 1 R1a1-M198− (or SRY10831.2*) ≥9000 2 R1a1a-M198+ M417− (or M17*) ≥9000 3 The Tenths, NW branch, ~4575 s, DYS448 null mutation 1125 ± 200 3700 ± 400 27 (WC) 2600 ± 300 28 Z280-Z92, North Eurasian (NEA) 4450 ± 450 29 Z280, NEA-1 3600 ± 400 30 Z280, NEA-2 2350 ± 300 31 Z280, Western Eurasian (WEA) 4100 ± 500 32 Z280, WEA-1 2300 ± 300 33 Z280, Eastern Carpathian (EC) 2600 ± 300 34 Z280, Balto-Carpathian (BC) 4300 ± 500 35 Z280, BC-1 2550 ± 300 36 Z280, BC-2 2200 ± 250 37 Z280, Northern Carpathian (NC) 2150 ± 300 38 Z280, L784 2500 ± 450 4 The Tenths, L664+ ~4575 5 The Tenth1500 ± 325 and 260 ± 60 (USA) 6 Z93+ ~5700, started to expand ~4050 7 Z93-L342.2+ 4900 ± 500 8 Z93-L342.2-L657+ 4050 ± 460 9 Z93-L342.2, Jewish branch 4475 ± 400, started to expand 1300 ± 150 10 Z93-L342.2, Bashkir branch 11 Z93-L342.2, Kyrgyz branch 2100 ± 250 12 Z283 (Eurasian branch) ~5500 13 M458 (European branch) ~4200 14 M458 CE parent ~3100 15 M458 CE-1 2900 ± 400 16 M458 CE-2 2900 ± 400 17 M458-L260 WS 2700 ± 300 18 Z284 (Scandinavian branch) 4300 ± 500 19 Z284-Z287, Old Scandinavian 20 Z284-L448, Young Scandinavian 2700 ± 350 21 Z284-L448-L176.1, Scottish Highland 1850 ± 290 22 Z280, Central Eurasian (CEA) 4900 ± 500 23 Z280, CEA-1, Russian Plain 4600 ± 500 24 Z280, CEA-2 3500 ± 400 25 Z280-L366, CEA-3 2500 ± 350 26 Z280-L365, North European (NE) 2600 ± 350 Z280-P278.2, Western Carpathian Appendix 2 lo as h io s N h o h h io h h olts u h ct p The following DNA projects were selected as primary hap- type databases: http://www.familytreedna.com/public/R1aY-Haploroup/default. px?vgroup=R1aY-Haplogroup§ion=yresults http://www.familytreedna.com/public/R1a/default.aspx? section =yresults ttp://www.familytreedna.com/public/r-arabi a/default.aspx?sect n=yresult http://www.familytreedna.com/public/Hungarian_Magyar_Y-D A_Project/default.aspx?section=yresults ttp://www.familytreedna.com/public/India/default.aspx?sectio n=yresults http://www.familytreedna.com/public/Turkic/default.aspx? secti n=yresults ttp://www.familytreedna.com/publicwebsite.aspx?vgroup=ger many§ion=yresults ttp://www.familytreedna.com/public/Finland/default.aspx?sect n=yresults http://www.familytreedna.com/public/Czech/default.aspx?secti on=yresults ttp://www.familytreedna.com/public/slovakia/default.aspx?sec tion=yresults ttp://www.familytreedna.com/public/polish/default.aspx? secti n=yresu http://www.familytreedna.com/public/russian_impire.aspx/defa lt.aspx?section=yresults ttp://www.familytreedna.com/public/russiadna/default.aspx? se ion=yresults Reference data were selected according to SNP assignment from YSearch database: (http://www.ysearch.org) and public rojects of FTDNA (http://www.familytreedna.com). Copyright © 2012 SciRes. 156 |