E. NOWAK ET AL. 51
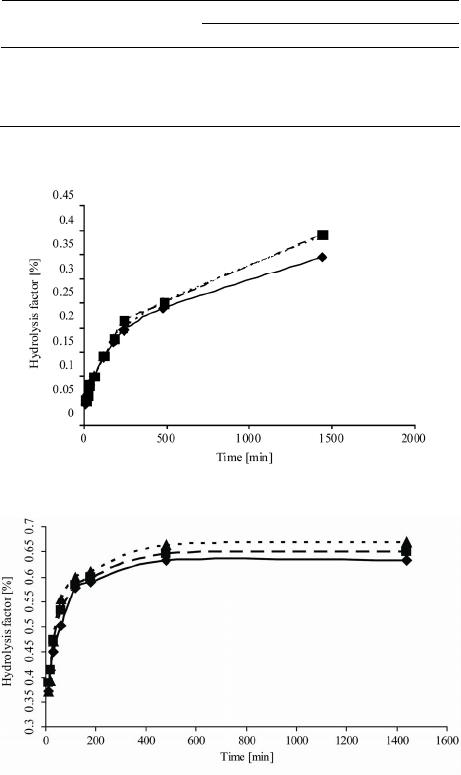
solutions with WLPL enhanced the starch hydrolysis rate
and extent. Simultaneously, the illumination of α-amy-
lase led to significant changes in the secondary confor-
mation of the enzyme protein molecule. It has been as-
sumed that conformational changes in the enzyme pro-
tein structure were, at least partially, responsible for the
observed enhancement of hydrolytic activity of pre-il-
luminated α-amylase. Protein compositions of α-amylase
and cellulases WT and TR are entirely different. Amy-
lase is a single chain glycoprotein of about 475 residues
and molecular weight in the range 50,000 [22]. Cellu-
lases WT and TR are complex of three physically distinct
components [23]. Such differences in molecular structure
between amylase and cellulases studied could explain
lack of conformational changes in proteins of WLPL
activated cellulases.
4. Conclusions
1) Stimulation with white linearly polarized light of
two cellulases WT and TR isolated from Trichoderma
reesei increases the enzyme specific activity of the
microcrystalline cellulose digestion and rate constants of
both stages of enzymatic hydrolysis of microcrystalline
cellulose.
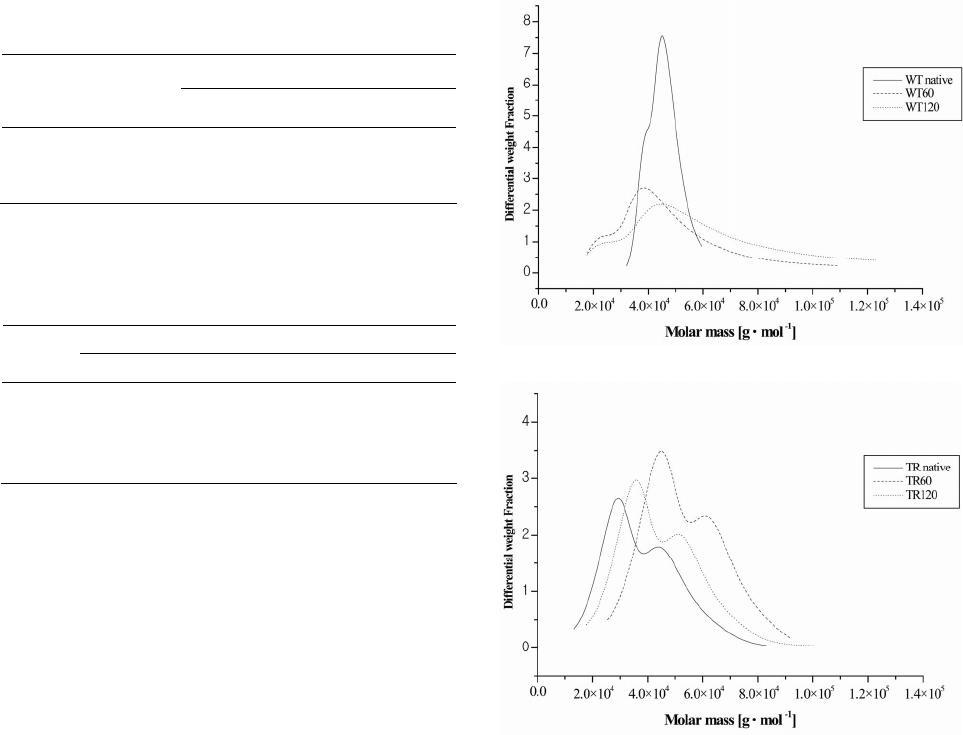
2) The WLPL stimulation of cellulases significantly
influences molecular structure of cellulose chains in di-
gested microcrystalline cellulose compared to these re-
sulting from the hydrolysis with original enzyme.
3) Illumination of both WT and TR enzymes led to
significant changes of molecular weight of enzyme protein
chains, however, activation of enzymes with WLPL did
not change enzyme protein secondary structure.
5. Acknowledgements
This work was financed by grant N N313 265936 from
National Science Center, Poland.
REFERENCES
[1] K. Wickholm, E. Hult, P. Larsson, T. Iversen and H.
Lennholm, “Quantification of Cellulose Forms in Com-
plex Cellulose Materials: A Chemometric Model,” Cel-
lulose, Vol. 8, No. 2, 2001, pp. 139-148.
doi:10.1023/A:1016700325434
[2] L. R. Lynd, “Overview and Evaluation of Fuel Ethanol
from Cellulosic Biomass: Technology, Economics, the
Environment, and Policy,” Annual Review of Energy and
the Environment, Vol. 21, 2001, pp. 403-465.
doi:10.1146/annurev.energy.21.1.403
[3] L. R. Lynd, R. T. Elander and C. E. Wyman, “Likely
Features and Costs of Mature Biomass Ethanol Technol-
ogy,” Applied Biochemistry and Biotechnology, Vol. 57-
58, 1996, pp. 741-761. doi:10.1007/BF02941755
[4] L. Laureano-Perez, F. Teymouri, H. Alizadeh and B. E.
Dale, “Understanding Factors That Limit Enzymatic Hy-
drolysis of Biomass,” Applied Biochemistry and Bio-
technology, Vol. 121, 1996, pp. 1081-1100.
[5] H. Hoeksema, S. Monstrey, K. Van Landuyt, Ph. Blon-
deel, P. Tonnard and A. Verpaele, “The Use of Polarised
Light in the Treatment of Severely Burned Patients (Ab-
stract),” 10th Congress of the International Society for
Burn Injuries, Jerusalem, 1-6 November 1998, pp. 1-6.
[6] K. Depuydt, S. Monstrey and H. Hoeksma, “The Stimu-
lating Effects of Polarized Light on Wound Healing and
Avoiding Surgery in the Treatment of Deep Dermal Burn
Wounds Using Polarized Light,” 10th Annual Meeting of
the European Association of Plastic Surgeons, Madrid, 21
May 1999, pp. 21-25.
[7] W. Vanscheidt, “The Effect of Polarized Light on Wound
Healing,” European Journal of Plastic Surgery, Vol. 24,
No. 8, 2002, pp. 383-390.
doi:10.1007/s00238-001-0306-z
[8] E. Bazso, Sz. Varju, P. Szego, K. Roza and P. Apai, “Ap-
plication of Incoherent Wide Band Polarised Light to
Promote Healing of Wounds,” Central Research Institute
for Physics, Budapest, 1982, pp. 121-130.
[9] W. Stegmann, “Behandlung des Ulcus Cruris mit Polar-
isiertem Licht,” Phlebologie und Proktologie, Vol. 14,
1985, pp. 96-97.
[10] M. Fenyö, “Theoretical and Experimental Basis of Bio-
stimulation,” Optics & Laser Technology, Vol. 16, 1984,
pp. 209-215. doi:10.1016/0030-3992(84)90029-X
[11] M. Fiedorowicz and G. Chaczatrian, “Effect of Illumina-
tion with the Visible Polarized and Non-Polarized Light
on α-Amylolysis of Starches of Different Botanical Ori-
gin,” Journal of Agricultural and Food Chemistry, Vol.
51, No. 26, 2003, pp.7815-7819. doi:10.1021/jf026202r
[12] A. Konieczna-Molenda, V. M. F. Lai, M. Fiedorowicz, G.
Khachatryan and P. Tomasik, “Effect of Linearly Polar-
ized Light upon Xylanase Activity,” Biotechnology Pro-
gress, Vol. 24, No. 2, 2008, pp. 385-388.
doi:10.1021/bp070394z
[13] A. Konieczna-Molenda, M. Fiedorowicz, W. Zhong and P.
Tomasik, “Polarized Light-Stimulated Enzymatic Hy-
drolysis of Chitin and Chitosan,” Carbohydrate Research,
Vol. 343, No. 18, 2008, pp. 3117-3119.
doi:10.1016/j.carres.2008.09.007
[14] M. Fiedorowicz, A. Konieczna-Molenda and G. Khacha-
tryan, “Stimulation of Cyclodextrin-Glycosyltransferase
(Turozyme) Activity by Illumination with Linearly Po-
larized Visible Light,” Biotechnology Progress, Vol. 25,
No. 1, 2009, pp. 147-150. doi:10.1002/btpr.90
[15] A. Konieczna-Molenda, M. Fiedorowicz and P. J. Toma-
sik, “Stimulation of Glucose Oxidase with White Linearly
Polarized Light,” Biotechnology Progress, Vol. 26, No. 2,
2010, pp. 393-396.
[16] G. L. Miller, “Use of Dinitrosalicylic Acid Reagent for
Determination of Reducing Sugars,” Analytical Chemis-
try, Vol. 31, No. 3, 1959, pp. 426-428.
doi:10.1021/ac60147a030
[17] A. Dupont and G. Harrison, “Conformation and dn/dc
Determination of Cellulose in N,N-Dimethylacetamide
Copyright © 2012 SciRes. OJOPM