Journal of Cancer Therapy
Vol.2 No.4(2011), Article ID:7803,6 pages DOI:10.4236/jct.2011.24079
Does Photofrin II Combined with a Radio-Adaptive Dose Lead to a Synergistic or Additive Effect after Ionising Irradiation in Vitro?
![]()
1Department of Oncology, Sefad Medical Center, Bar Ilan School of Medicine, Sefad, Israel; 2Department of Anaesthesiology and Reanimation, University of Regensburg, Regensburg, Germany; 3Institution of Clinical Radiology, University of Munich, Munich, Germany; 4Department of Radiation Therapy, Bad Trissl Clinic, Oberaudorf, Germany; 5Department of Surgery, University of Milan, Milan, Italy; 6Laser Research Laboratory, University of Munich, Munich, Germany; 7Radiation Protection and Health, Federal Office for Radiation Protection, Munich, Germany.
Email: *m.schaffer@onlinehome.de
Received June 7th, 2011; revised July 20th, 2011; accepted July 29th, 2011.
Keywords: Adative Dose, Photofrin II, Ionising Irradiation, Synergistic/Additive Effect
ABSTRACT
Background: The radiosensitizing effect of Photofrin II has been demonstrated in vitro and in animal models, even in tumor models known to be highly radioresistant, such as glioblastoma and bladder carcinoma. Radio-adaptive doses are also known to lead to an augmented cell or tissue reaction. The aim of this study was to investigate potential synergistic or additive effects when combining the two methods in vitro for an improved therapeutic concept in bladder cancer. Material and Methods: RT4 human bladder carcinoma cell line and HCV29 human bladder epithelium cells were seeded and incubated with various concentrations of Photofrin II. The cells were additionally irradiated with ionizing radiation (0.05 Gy/2 Gy/0.05 Gy + 2 Gy). Cells without Photofrin II incubation and irradiation served as controls. The cell survival was evaluated. Results: The survival rate of both cell lines, RT4 and HCV29, did not differ significantly when incubated with a non-toxic concentration of Photofrin II and exposed to a pre-irradiation dose of 0.05 Gy prior to the 2 Gy radiation fraction, compared to cells exposed to Photofrin II plus a 2 Gy ionizing radiation. Conclusions: The combination of both methods did neither demonstrate a synergistic or additive effect nor did it lead to a negative influence of both modulating factors in an in vitro setting.
1. Introduction
The biological effects of radiation manifest themselves both in the irradiated tumor and in the surrounding tissue. The selectivity of radiation can be improved by using 1) sophisticated computerized dosimetry, e.g. 3D irradiation, 2) tailor-made irradiation protocols, e.g. high precision radiation therapy, or 3) various irradiation sources such as electrons, photons, or protons. Another approach to optimize the effects on the tumor tissue is based on the introduction of a radiation reaction effect, which depends on cellular biology (i.e. oxygenation, cell cycle, etc.) and which can be modified by chemicals (sensitizers, protectors and chemotherapy) acting as radiosensitizers [1-3]. The observations published in the 1950s and 1960s by Cohen and Schwartz [4,5] showed that hematoporphyrin derivate (HpD), a highly heterogeneous chemical derivative of Hp, of which Photofrin II represents a partially purified form [6], can act as a radiosensitizer for tumours. Several studies in vitro, on murine tumour models, and on patients with different tumours and tumour stages have demonstrated the in vivo and in vitro efficacy of Photofrin II as both a specific and a selective radiosensitizing agent [7-13].
The radio-adaptive response is a biopositive effect induced by a low priming dose, which can be observed after the application of a higher challenging dose on different tissues. In 1984, the adaptive response was first recognized, when Olivieri et al. demonstrated that human lymphocytes exposed to low concentrations of radioactive thymidine showed fewer chromosomal aberrations caused by a 1.5 Gy challenging dose than those not preexposed to irradiation [14]. Several publications have studied the effect with different cell lines, different preirradiation doses, and variable challenging doses [15-18]. The exact mechanism of the effect is still unknown. An altered gene expression caused by low-dose ionizing radiation has been identified. A radio-adaptive response seems to be associated with an up-regulation of DNA repair and stress response genes and a down-regulation of cell cycle control and apoptosis genes. TP53 (Tumour Protein 53) is supposed to play an important role in this mechanism [19]. Protein synthesis, metabolism and signal transduction appear to be involved in the adaptive response as well [15].
It was previously described by Schwarz et al. that normal bladder cells (HCV29) and bladder cancer cells (RT4) demonstrated a different reaction to radio-adaptive doses. After a pre-irradiation dose of 0.05 Gy, an induced radio-resistance was demonstrated in HCV29 cells, while RT4 cells showed an augmented radiosensitivity [20]. The effect of radio-adaptive doses was also evaluated in the HT29 cell line (human colorectal cancer cells) and in the GM637 cell line (human fibroblasts). The results demonstrated that the application of 0.05 Gy prior to a 2 Gy fraction enhanced the response of colorectal cancer cells, while the response of normal fibroblasts was not augmented [21].
When combining these two methods considerable synergistic or additive effects on tumor cell lines could potentially ensue.
The aim of our study was therefore to assess the effect of a combined application of the radiosensitizer Photofrin II and a radio-adaptive dose on RT4 and HCV29 cells.
2. Material and Methods
2.1. Chemicals
Photofrin II was purchased from AXCAN PHARMA (Mont-Saint-Hilaire, Canada) as a freeze-dried porfimer. It was stored as a stock solution in 5% dextrose solution (DeltaSelect, Pfullingen, Germany) at a concentration of 2.5 mg/ml and kept at –20˚C until use. Storage, dilution steps, and the incubation period were performed under experimental conditions avoiding the exposure to light. The cell proliferation reagent WST-1 was obtained from Roche Diagnostics (Mannheim, Germany).
Chemicals and additives for cell culture were purchased from Gibco Invitrogen (Karlsruhe, Germany) unless otherwise specified.
2.2. Cell-Lines and Cultures
The human bladder carcinoma cell line RT4, which is known to be radioresistant in vivo, and the HCV29 human bladder epithelium cell line were grown separately in RPMI 1640 medium containing Glutamax (LAlanylL-Glutamine), supplemented with 10% foetal calf serum (FCS), 1% sodium pyruvate (100 mM, Sigma-Aldrich), and 1% Eagles minimal essential medium with Earl’s salts [22,23].
The cells were maintained in a humidified incubator with 5% CO2 at 37˚C. Stock cultures were kept in 80 cm3 flasks (Nalgene Nunc, Wiesbaden, Germany) and were passaged once per week in the exponential growing phase, using 0.05% trypsin plus 0.02% EDTA in PBS at 37˚C, but not more than ten times.
2.3. Experimental Set Up
All experiments were performed under protection from light. RT4 and HCV29 cells were seeded in 96-well culture dishes at a density of 500 cells per well and allowed to adapt for 24 h. In addition, one 96-well plate (standard plate) with an increasing cell number per row (0-63-125- 250-500-750-1000-1250-1500-2000 cells per well), but no further treatment or irradiation, was prepared to monitor cell growth and to serve as a survival reference. After 24 h, Photofrin II was added at a final concentration of 2.5 μg/ml, 5 μg/ml, 7.5 μg/ml or 10 μg/ml to one row of each of the four culture dishes. Different concentrations were administered to simulate the accumulation in vivo. In two rows without radiosensitizer addition, culture medium was added to reach the same final volume per well. All experiments were repeated at least 4 times for a minimum of 24 single data analyses.
2.4. Irradiation
After a Photofrin II incubation period of 24 h, half of the plates were irradiated with a dose of 0.05 Gy at a dose rate of 0.03 Gy/min (225 kV, 5 mA, 0.35 mm Cu). 4 h after pre-irradiation, cells in one plate with and one plate without pre-irradiation were further irradiated with 2 Gy at a dose rate of 1.0 Gy/min (225 kV, 15 mA, 0.35 mm Cu). As a result, four different irradiation groups were evaluated: 0 Gy (control), 0.05 Gy, 2 Gy, 0.05 Gy prior to 2 Gy.
The ionizing irradiation was performed using a Mueller RT 250 X-ray device (Table 1). Cells were kept at 37˚C during the entire irradiation process. The non-irradiated plates underwent the same procedure as irradiated cells to simulate the same conditions.
2.5. Cell Viability Test
After irradiation, the cells were cultured for 4 days. The response of the cells to irradiation was evaluated by determining the survival of the proliferating cells. Possible effects of the irradiation and Photofrin II incubation were
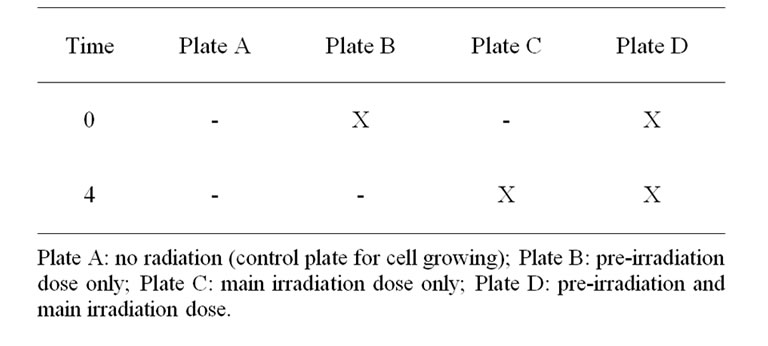
Table 1. Irradiadion mode.
evaluated by a tetrazolium-based colorimetric WST test where the number of metabolically active cells was quantified spectrophotometrically by an ELISA reader (MRX, Dynatec Laboratories) at 450 nm. The background of each well was measured prior to the addition of the cell proliferating reagent WST-1 (final dilution 1:10). After 3 h incubation, the optical density was measured. Each plate, including the control plate was analyzed by using the same experimental conditions. In order to verify whether the cell growth was still in the exponential phase, the standard plate with increasing seeded cell numbers was also analyzed.
2.6. Cell Survival Analysis
Each combination of irradiation on the plates was examined in four repeated separate experiments resulting in 24 single data sets. After debugging of the background, the evaluation of the cell survival and a calculation of the relative decrease after irradiation were performed. A Student t-test with a significance level set at p < 0.05 was adapted to evaluate the statistical significance of the results.
3. Results
3.1. RT4 Cells
Photofrin II-incubated but not irradiated cells showed a cytotoxic effect at a Photofrin II concentration of 7.5 μg/ml (p = 0.0000753) and 10.0 μg/ml (p = 6.0386E–14) in comparison with RT4 cells not incubated with Photofrin II.
After irradiation of PhotofrinII incubated RT4 cells at a concentration of 7.5 mg/ml and 10 µg/ml with a preirradiation dose of 0.05 Gy, a significantly lower cell survival rate was observed. At a Photofrin II dose of 2.5 µg/ml, the cell survival rate was significantly enhanced, while the survival rate tended to decrease at a concentration of 5 µg/ml. with a radiation dose of 2 Gy led to a significant decrease in survival rates for Photofrin II doses of 5 µg/ml (p = 0.04947688), 7.5 mg/ml (p = 0.00000011) and 10 µg/ml (p = 4.65461E–15).
The experiments performed with a pre-irradiation dose of 0.05 Gy prior to the 2 Gy radiation fraction demonstrated a significant decrease in cell survival as well. The cell survival rate was significantly higher for 7.5 µg/ml and 10 µg/ml Photofrin II concentrations compared to an irradiation at a dose of 2 Gy without Photofrin II incubation (Figure 1).
3.2. HCV29 Cells
Photofrin II incubation led to a significantly toxic effect on HCV29 cells at concentrations from 5 mg/ml to 10 µg/ml. At a concentration of 2.5 µg/ml, no significant alteration in the survival rate was observed.
A 0.05 Gy pre-irradiation did not lead to a significant decrease of HCV29 cell survival at a Photofrin II concentration of 2.5 µg/ml. The cell survival, however, decreased significantly from 5 mg/ml (p = 2.396E–16) to 10 µg/ml (1.114E–30) after 0.05 Gy.
After an irradiation with 2 Gy, the cell survival was not significantly reduced at a concentration of 2.5 mg/ml. A toxic effect was demonstrated at concentrations above 5 mg/ml.
The combination of a pre-irradiation with 0.05 Gy prior to the 2 Gy and a Photofrin II concentration of 2.5 µg/ml tended to enhance HCV29 cell survival. Both irradiation schemes tended to enhance cell survival after a combined pre-irradiation and challenging irradiation at a non-toxic Photofrin II concentration; however, the differences were not statistically significant (Figure 2).
4. Discussion
Former studies have demonstrated Photofrin II to be a sensitive and selective radiosensitizer [9-13]. An important aspect of Photofrin II is the ability to accumulate in
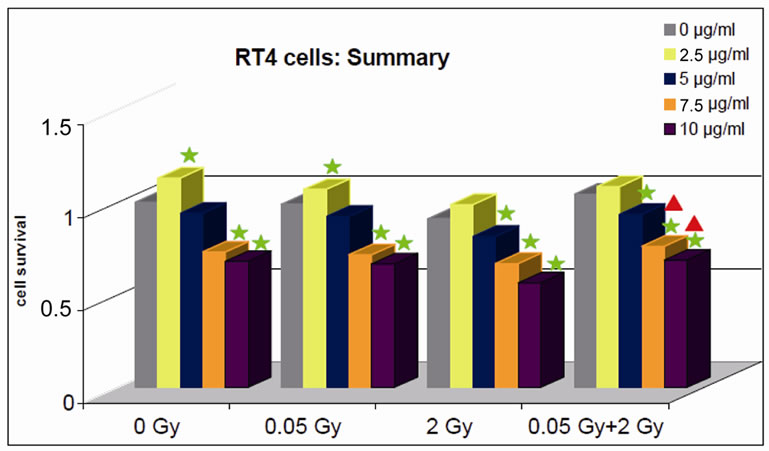
Figure 1. RT4 cells incubated with various concentrations of Photofrin II under the different irradiation schemes. A significant difference compared to the control is marked by  in the colour of the relevant dose. A significant difference of 0.05 Gy + 2 Gy compared to the irradiation with 2 Gy is marked by
in the colour of the relevant dose. A significant difference of 0.05 Gy + 2 Gy compared to the irradiation with 2 Gy is marked by .
.
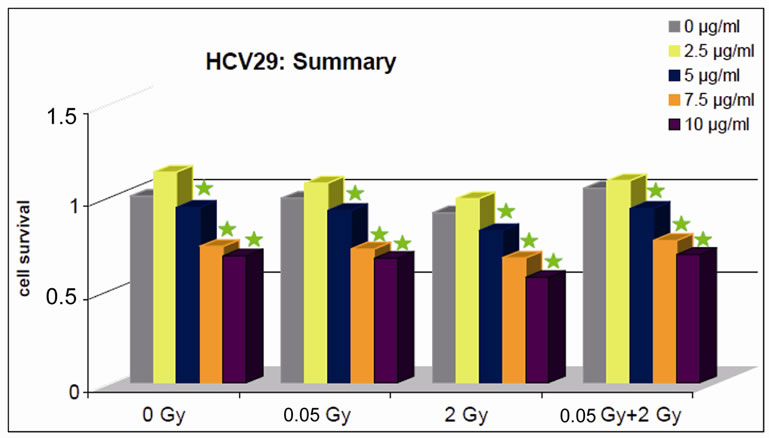
Figure 2. HCV29 cells incubated with various concentrations of Photofrin II under the different irradiation schemes. A significant deviation compared to the controls is marked by a  in the colour of the relevant dose.
in the colour of the relevant dose.
human tumour tissue in vivo. In the in vitro setting, it is not possible to achieve this accumulation of Photofrin II; however, the effect can be simulated by increasing the Photofrin II concentration.
The results of our study imply that the application of Photofrin II at a concentration of 2.5 µg/ml tends to result in an increase of HCV29 cell survival, while the RT4 cell survival tends to decrease at a concentration above 5 µg/ml after irradiation. In an in vivo model the application of a small Photofrin II dose may cause a higher radiosensitation targeted in the tumour cells.
HCV29 cells primed with 0.05 Gy and incubated with Photofrin II showed a better survival than non pre-irradiated cells. The application of a radio-adaptive dose may be viewed as a form of targeting therapy in oncology [24-25]. Radio-adaptive doses of 0.01 Gy to 1.5 Gy have been shown to render cells less susceptible to the induction of chromosomal aberrations, micronuclei formation, mutations and cell killing [25-28]. Schwarz et al. demonstrated the maximum effect of the adaptive response on HCV29 cells to occur at a pre-irradiation dose of 0.05 Gy [21]. The time frame between the radioadaptive dose and the application of high dose irradiation is important. A 4h interval between the application of the low and the high dose appears to be most effective [29].
Our results regarding the response of RT4 cells to Photofrin II are in agreement with previous studies, which had demonstrated the radiosensitizing activity of Photofrin II on bladder carcinoma and other tumours like Lewis sarcoma both in vivo and in vitro models [7,11,20, 21]. A detailed understanding of the mechanisms involved in tumour radiosensitization by Photofrin II is partially hindered by the highly heterogeneous chemical composition of this photosensitizer. There are suggestions that Photofrin II reacts with hydroxyland O- radicals intracellularly generated by the radiolysis of water [12]. Other possible mechanisms involve the inhibition of cellular repair processes after ionizing radiation damage. The presence of oxygen enhances the Photofrin IIpromoted radiation damage involving the formation of radical derivates of high reactivity, such as e.g. hydroxy radicals [13].
The radio-adaptive response is a biopositive effect induced by a low priming dose, which can be observed after the application of a higher challenging dose on different tissues [14]. The exact mechanism of the effect is still unknown. A radio-adaptive response seems to be associated with an up-regulation of DNA repair and stress response genes and a down-regulation of cell cycle control and apoptosis genes. TP53 (Tumour Protein 53) is supposed to play an important role in this mechanism The effect of the adaptive dose has been described as being dependent on the timing in relation to the phase in the cell cycle [15,16,21]. The duration of the processes induced by the priming dose is about four hours or two to three cell cycles; after this period the effect tapers [18,19, 27,29]. These processes are considered to mainly reflect mechanisms of induced repair.
The combination of Photofrin II and a radioadaptive dose could potentially lead to a novel radiotherapeutic regimen that enhances the destruction of the tumor while simultaneously protecting normal tissues.
Further studies will be needed to fully comprehend the mechanism of Photofrin II, the radio-adaptive response and the combination of these two mechanisms. In addition, potential clinical applications of both modulation factors need to be studied.
5. Conclusions
In conclusion, our study confirmed a radiosensitizing effect of Photofrin II on the human bladder carcinoma cells (RT4) and a protective effect induced by a pre-irradiation dose as a radio-adaptive response on human bladder epithelium (HCV29).
While these effects did not interact in a negative way in our in vitro model, a significant additive or synergistic could not be demonstrated. Considering the fact that Photofrin II accumulates in tumour tissue in vivo, a combination of both methods appears to be a feasible concept. Further studies will be needed to elucidate whether a synergistic or additive effect of these two approaches may be present in vivo.
REFERENCES
- W. H. McBride and H. R. Withers, “Biologic Basis of Radiation Therapy,” In: C. A. Perez and L. W. Brady, Eds., Principles and Practice of Radiation Oncology, Lippincott Williams and Wilkins, Philadelphia, 2004, pp. 96-136.
- T. H. Wassermann and J. D. Chapman, “Radiation Response Modulation,” In: C. A. Perez and L. W. Brady, Eds., Principles and Practice of Radiation Oncology, Lippincott Williams and Wilkins, Philadelphia, 2004, pp. 663-679.
- I. Patties, J. Jahns, G. Hildebrandt, R. D. Kortmann and A. Glasow, “Additive Effects of 5-Aza-2’-deoxycytidine and Irradiation on Clonogenic Survival of Human Medulloblastoma Cell Lines,” Strahlentherapie und Onkologie, Vol. 185, No. 5, 2009, pp. 331-338. doi:10.1007/s00066-009-1956-1
- L. Cohen and S. Schwartz, “Modification of Radiosensitivity by Porphyrins II. Transplanted Rhabdomyosarcoma in Mice,” Cancer Research, Vol. 26, 1966, pp. 1769-1773.
- M. Schaffer, P. Schaffer, M. Vogesser, B. Ertl-Wagner, et al., “The Application of Photofrin II as a Specific Radiosensitzing Agent, in Patients with Bladder-Cancer, Report of Two Cases,” Photochemical & Phobiological Science, Vol. 1, 2002, pp. 686-689. doi:10.1039/b203732g
- C. J. Byrne, L. V. Morshallsay and A. D. Wand, “The Chemical Composition of Photofrin,” Journal of Photochemistry and Photobiology B: Biology, Vol. 6, 1990, pp. 13-27.
- U. Kulka, M. Schaffer, P. M. Schaffer, et al., “Photofrin as an Efficient Radiosensitizer in an in Vitro Cell Survival Assay,” Biochemical & Biophysical Research Comunications, Vol. 311, No. 1, 2003, pp. 98-103. doi:10.1016/j.bbrc.2003.09.170
- U. Kulka, M. Schaffer, P. M. Schaffer, et al., “Untersuchungen zum Mechanismus der radiosensitivierenden Wirkung von Photofrin II,” Strahlentherapie und Onkologie, Vol. 181, No. 1, 2005, p. 104.
- M. Schaffer, P. M. Schaffer, L. Corti, G. Sotti, A. Hofstetter, G. Jori and E. Dühmke, “Photofrin II as an Efficient Radiosensitising Agent in an Experimental Tumour,” Onkologie, Vol. 24, No. 5, 2001, pp. 482-485. doi:10.1159/000055130
- M. Schaffer, P. M. Schaffer, L. Corti, et al., “Photofrin as a Specific Radiosensitising Agent for Tumours: Studies in Comparison to Other Porphyrins, in an Experimental in Vivo Model,” Journal of Photochemistry and Photobiology B: Biology, Vol. 66, 2002, pp. 157-164.
- M. Schaffer, B. Ertl-Wagner, P. M. Schaffer, et al., “The Application of Photofrin II as a Sensitizing Agent for Ionizing Radiation—A New Approach in Tumour Therapy?” Current Medicinal Chemistry, Vol. 12, No. 10, 2005, pp. 763-771. doi:10.2174/0929867053764653
- M. Schaffer, U. Kulka and P. M. Schaffer, “The Role of Radical Derivatives of High Reactivity in the Radiosensitizing Action of Photofrin II,” Journal of Porphyrins and Phthalocynines, Vol. 10, No. 12, 2006, pp. 1398-1402. doi:10.1142/S1088424606000776
- P. M. Schaffer, U. Kulka and B. Ertl-Wagner, “The Influence of Oxygen on the Radiosensitizing Activity of Photofrin II and Hypericin,” Journal of Porphyrins and Phthalocyanines, Vol. 11, No. 10, 2007, pp. 1-6. doi:10.1142/S1088424607000849
- G. Olivieri, J. Bodycote and S. Wolff, “Adaptive Response of Human Lymphocytes to Low Concentrations of Radioactive Thymidine,” Science, Vol. 223, No. 4636, 1984, pp. 594-597. doi:10.1126/science.6695170
- M. A. Coleman, E. Yin, L. E. Peterson, D. Nelson and K. Sorensen, “Low-Dose Irradiation Alters the Transcript Profiles of Human Lymphoblastoid Cells Including Genes Assosiated with Cytogenetic Radioadaptive Response,” Radiation Research, Vol. 164, No. 4, 2005, pp. 369-382. doi:10.1667/RR3356.1
- P. Cramers, P. Atanasova and H. Vrolijk, “Pre-exposere to Low Doses: Modulation of X-Ray-Induced DNA Damage and Repair?” Radiation Research, Vol. 164, No. 4, 2005, pp. 383-390. doi:10.1667/RR3430.1
- S. Sadekova, S. Lehnert and T. Y. K. Chow, “Induction of PBP74/Mortalin/Grp75, a Member of the hsp70 Family, by Low Doses of Ionizing Radiation: A Possible Role in Induced Radioresistance,” International Journal of Radiation Biology, Vol. 72, No. 6, 1997, pp. 653-660. doi:10.1080/095530097142807
- M. S. Sasaki, Y. Ejima, A. Tachibana, et al., “DNA Damage Response Pathway in Radioadaptive Response,” Mutation Research, Vol. 504, No. 1-2, 2002, pp. 101-118. doi:10.1016/S0027-5107(02)00084-2
- R. Okazaki, A. Ootsuyama and T. Norimura, “TP53 and TP53-Related Genes Associated with Protection from Apoptosis in the Radioadaptive Response,” Radiation Research, Vol. 167, No. 1, 2007, pp. 51-57. doi:10.1667/RR0623.1
- M. Schaffer, S. B. Schwarz and U. Kulka, “Adaptive Doses of Irradiation—An Approach to a New Therapy Concept for Bladder Cancer?” Radiation and Environmental Biophysics, Vol. 43, No. 4, 2004, pp. 271-276. doi:10.1007/s00411-004-0256-5
- S. B. Schwarz, P. M. Schaffer and U. Kulka, “The Effect of Radio-Adaptive Doses on HT29 and GM637 Cell,” Radiation Oncology, Vol. 3, 2008, p. 12. doi:10.1186/1748-717X-3-12
- M. A. Bean, H. Pees, J. E. Fogh, H. Grabstald and H. F. Oettgen, “Cytotoxicity of Lymphocytes from Patients with Cancer of the Urinary Bladder: Detection by a 3Hproline Microtoxicity Test,” International Journal of Cancer, Vol. 14, No. 2, 1974, pp. 186-197. doi:10.1002/ijc.2910140207
- J. R. W. Masters, P. J. Hepburn, L. Walker et al., “Tissue Culture Model of Transitional Cell Carcinoma: Characterization of Twenty-Two Human Urothelial Cell Lines,” Cancer Research, Vol. 46, 1986, pp. 3630-3636.
- P. Balermpas, M. Hambek, O. Seitz, C. Rödel and C. Weiss, “Combined Cetuximab and Reirradiation for Locoregional Recurrent and Inoperable Squamous Cell Carcinoma of the Head and Neck,” Strahlentherapie und Onkologie, Vol. 185, No. 12, 2009, pp. 775-781. doi:10.1007/s00066-009-2092-7
- K. Dittmann, C. Mayer and H. P. Rodemann, “Nuclear EGFR as Novel Therapeutic Target Insights into Nuclear Translocation and Function,” Strahlentherapie und Onkologie, Vol. 186, No. 1, 2010, pp. 1-6. doi:10.1007/s00066-009-2026-4
- J. D. Shadley and S. Wolff, “Very Low Doses of X-Rays can Cause Human Lymphocytes to Become Less Susceptible to Ionising Radiation,” Mutagenesis, Vol. 2, No. 2, 1987, pp. 95-96. doi:10.1093/mutage/2.2.95
- J. D. Shadley, “Chromosomal Adaptive Response in Human Lymphocytes,” Radiation Research, Vol. 138, No. 1, 1994, pp. 9-12. doi:10.2307/3578750
- A. M. Ueno, D. B. Vannais, D. L. Gustafson, J. C. Wong and C. A. Waldren, “A Low, Adaptive Dose of GammaRays Reduced the Number and Altered the Spectrum of S1-Mutants in Human-Hamster Hybrid AL Cells,” Mutation Research, Vol. 358, No. 2, 1996, pp. 161-169. doi:10.1016/S0027-5107(96)00117-0
- S. Venkat, S. K. Apte, R. C. Chaubey and P. S. Chauhan, “Radioadaptive Response in Human Lymphocytes in Vitro,” Journal of Environmental Pathology, Toxicology and Oncology, Vol. 20, 2001, pp. 165-175.