American Journal of Plant Sciences
Vol. 4 No. 7 (2013) , Article ID: 34755 , 8 pages DOI:10.4236/ajps.2013.47182
In Vitro Mass Scale Propagation of Wild Cymbidium lowianum with a Rare and Endangered Plant
![]()
1Institute of Landscape Plants, Yunnan Agricultural University, Kunming, China; 2Yunnan Vocational and Technical College of Agriculture, Kunming, China; 3Associate Innovation Center of Biodiversity Species and Yunnan Characteristic Agriculture, Kunming, China; 4Foreign Languages Institute, Yunnan Agricultural University, Kunming, China; 5Institute of Horticulture, Yunnan Academy of Agricultural Sciences, Kunming, China.
Email: *lzl-yn@sohu.com
Copyright © 2013 Yuying Wang et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received April 30th, 2013; revised May 31st, 2013; accepted June 15th, 2013
Keywords: Wild Cymbidium lowianum; Seed Germination; Propagation; Protocorm; Multiple Shoots
ABSTRACT
The wild Cymbidium lowianum, a national-level rare and endangered species of Orchid, is an excellent garden plant with ornamental flowers with striking, deep red lips in a V-shaped formation. The objective of this study was to establish a micropropagation protocol system via immature seeds of wild Cymbidium lowianum, evaluate the Murashige and Skoog (MS), half-strength Murashige and Skoog (1/2 MS) medium, 6-benzylaminopurine (BA), a-napthaleneacetic acid (NAA), organic additions activated charcoal (AC) and banana pulp (BP) effects on the different morphogenesis (seed germination, multiple shoot and rooting) in vitro. The optimal combination for the germination of seed was 1.0 mg·L−1 BA with 0.5 mg·L−1 NAA in 1/2 MS, and addition 0.3%AC, which resulted in 95% seed germination in 90 days. The best formulation for multiple shoot was 1/2 MS medium containing 2.5 mg·L−1 BA, 0.5 mg·L−1 NAA and addition 8% BP in which produced 19.8 shoots per protocorm in 60 days. Multiple shoots were cut and rooted in 1/2 MS supplemented with 1.5 mg·L−1 NAA,0.1 mg·L−1 BA and 0.3% AC, roots initiated 20 days after culture, the rooting percentages reached to 100%, in which 4.7 per shoot produced roots in 60 days. The survival rate of plantlet was up to 92% in moss after 30 days. This finding reveals that it is possible to obtain in vitro culture of Cymbidium lowianum using immature seeds in asymbiotic culture.
1. Introduction
Orchids are highly valued flowers and are counted among the top ten most known flowers in China [1]. The wild C. lowianum is a plant of the Orchidaceae, Cymbidium, Subgen. Cyperorchis (Bl.) Seth et Cribb, Sect. Iridorchis (Bl.) P. F. Hunt, and is considered as a wild endangered plant under state protection (category II) recorded in the China Plant Red Data Book, while is an excellent garden plant with ornamental flowers with striking, deep red lips in a V-shaped formation from April to May. It is distributed in Longling, Baoshan Cangyuan, Mengla, Menghai, Jinghong, Luchun, and in Tibet of Yunnan province. Due to its unique shape and long flowering period, the C. lowianum used as a material for bonsai and cut-flower arrangements, and is threatened by over-collection from its natural habitat for horticultural purposes.
Orchids usually adopt the traditional ramet reproduction method, which has shortcomings of long cycle and low propagation coefficient. In addition to long-term asexual reproduction, the growing number of plant viruses has also contributed to the decline in the quality of orchids and the serious degradation of the species, making it difficult to meet market requirements. In 1921, Knudson created the nonsymbiotic germination of orchid seeds, Since then protocols for rapid propagation of different Cymbidium sp. by using mature seeds [2], immature seeds [3], flower stalks, pseudo bulbs [4], shoot segments [5], flower buds, thin cell layers (TCLs) of protocorm-like bodies (PLBs) [6] and artificial seeds based on PLBs [7] have been reported. Due to the fact that there are no existing studies concerning the tissue culture of this species, the objective of this paper is to establish a protocol for the in vitro propagation system of C. lowianum using immature seeds.
2. Materials and Methods
2.1. Artificial Pollination
The plants of Wild Cymbidium lowianum were collected in Baoshan of Yunnan province, China, on 16 May 2007, cultivated 3 years in the greenhouse of Yunnan Agricultural University, China. After two days the flowers opened (Figure 1(A)), artificial pollination was conducted at 3 PM on sunny days (Apr to May 2010) under the conditions: temperature 23˚C to 25˚C, humidity 80 to 98%. The hangings were tagged, while observing the development of the fruit after pollination.
2.2. Seed Harvesting and Sterilization
The mature but not yet cracked capsules of C. lowianum (Figure 1(B)) were collected after artificial pollination (111 days). The freshly collected capsules were surface-sterilized in 75% alcohol for 2 min and washed thoroughly in water. The capsules were immersed in 75% alcohol for disinfection (3 - 6 min) and in 10% sodium hypochlorite for disinfection (12 - 15 min). Next, they were washed four times in sterile water. The sterilized capsules were cut longitudinally with the help of a sharp sterilized surgical blade to extract the seeds. The seeds (Figure 1(C)) were scooped out from sterilized capsules and sown by spreading thinly as possible over the surface of the asepsis culture medium in a glass bottle, with each bottle containing 40 ml of medium.
2.3. Seed Germination, Multiple Shoots and Rooting of Shoots
Seeds were inoculated in two salt formulation basic medium media, MS (Murashige and Skoog) (1962) and 1/2 MS (Half-strength Murashige and Skoog), and were supplemented with 6-Benzylaminopurine (BA) (0.0, 0.5, 1.0, and 2.0 mg·L−1), a-naphthalene acetic acid (NAA) (0.0, 0.1, 0.5, and 1.0 mg·L−1), and additional 0.3% of the activated charcoal (AC) to determine the suitable medium for seed germination. The embryos enlarged and occupied the entire seed coat and developed into the pre-protocorm stage (Figure 1(D)), the germination percentage was calculated using the total number of seeds. The average number of seeds per individual inoculated was 100 ± 30.
The shoot buds first appeared as small green protuberances all over the protocorm, which eventually developed into multiple shoots within 8 - 10 weeks (Figures 1(E) and (H)). PLBs (diameter > 2 mm) were individually inoculated in the MS and 1/2 MS basic medium, and supplemented with BA (0.0, 0.5, 1.5, and 2.5 mg·L−1), NAA (0.0, 0.1, 0.5, and 1.0 mg·L−1), additional 8% of the banana pulp (BP) for further proliferation of protocorm and shoot development.

Figure 1. Seed germination and seedling development in C. lowianum: (A) Flower; (B) Fruit; (C) Seed at the time of inoculation; (D) Swelling of the embryo; (E) Initiation of multiple shoots; (F) Appearance of white seedling on ½ MS medium with BA (2.5 mg·L−1), NAA (0.5 mg·L−1), and BP (8%); (G) Multiple white seedling on 1/2 MS medium with BA (2.5 mg·L−1), NAA (0.5 mg·L−1), and BP (8%); (H) Development of multiple shoots on 1/2 MS medium with BA (2.5 mg·L−1), NAA (0.5 mg·L−1), and BP (8%); (I) Development of roots on 1/2 MS medium with NAA (1.5 mg·L−1), BA (0.1 mg·L−1, and AC (0.3%); (J) Ex vitro establishment of plantlets on the moss; (K) Individually potted plants.
For root development, each individual shoot with 3 - 5 expanded leaves and 3 - 4 cm in height were detached from the shoot clumps and transferred to MS, 1/2 MS, and supplemented with BA (0.0, 0.1, and 0.5 mg·L−1), NAA (0.0, 1.0, 1.5, and 2.0 mg·L−1), and additional 0.3% AC.
2.4. Ex Vitro Plant Establishment
For ex vitro establishment, well-rooted plantlets were rinsed thoroughly in sterile water to remove residual nutrients from the plant body. The plantlets were transplanted to plastic pots containing a mixture of ceramisite and tree shell (1:1), sand, muskeg, and leaf litter. The transplants were immediately transferred into greenhouse under the following conditions: temperature 23˚C to 25˚C, humidity 80% to 95%. The hardened plantlets were maintained in the greenhouse at Yunnan Agricultural University, China.
2.5. Culture Conditions
All culture mediums were supplemented with 3% sucrose, 0.7% article agar (made in China), pH at 5.4 - 5.8, temperature at 25˚C ± 2˚C, and humidity at 80% - 85%. All cultures adopted light culture, except for seed germination, with intensity of 1200 lux.
2.6. Experimental Design and Data Analysis
The layout of the experiment was according to a completely randomized design with three replications. Germination seed, multiple shoots, rooting and plant ex vitro plant establishment experiments were measured on days 90, 60, 30 and 30. Data were subjected to analysis of variance (ANOVA) and means were performed by the Duncan’s multiple range test (Duncan, 1995), and the profile plots performed by General Linear Model using the SPSS (SPSS Inc., Chicago, IL, USA). Mean separation was carried out using the least significant difference method at an alpha 0.05 level.
3. Results
3.1. Asymbiotic Seed Germination
After different 20 kinds of culture medium treatment for 90 days, different kinds of culture medium led to different seed germination (Table 1). The results showed that the highest seed germination (95%) was observed in 1/2 MS + BA 1.0 mg·L−1 + NAA 0.5 mg·L−1 and 0.3% AC medium, and there were significantly difference compared to others treatment (P ≤ 0.05).
Followed by that in 1/2 MS + BA 1.0 mg·L−1 + NAA 0.1 mg·L−1 + 0.3% AC medium (80%) and in MS + BA 1.0 mg·L−1 + NAA 0.5 mg·L−1 + 0.3% AC medium (75%). The lowest seed germination (4%) was in MS + 0.3% AC. The average rates of seed germination were
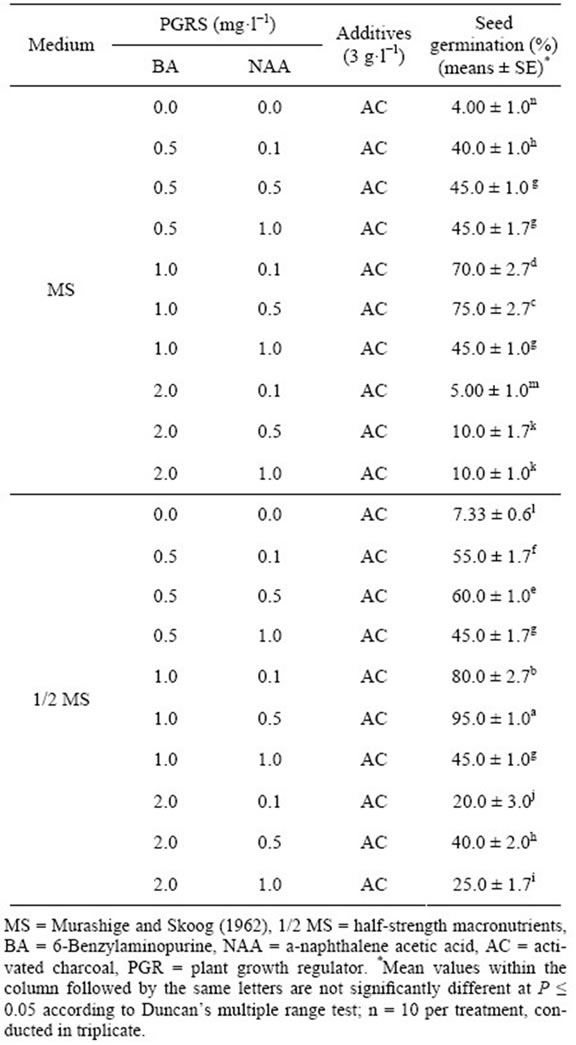
Table 1. Comparative effect of culture media on asymbioticgermination of seeds of C. lowianum after 90 d.
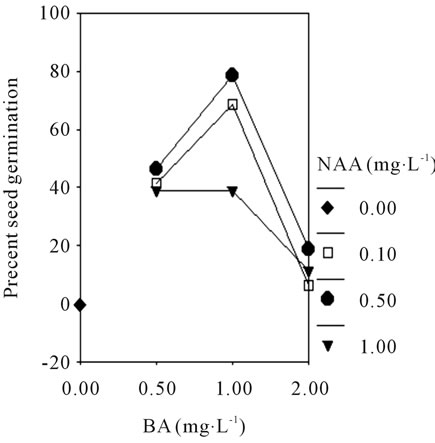
34.9% in MS basic medium and 47.2% in 1/2 MS basic medium with different plant growth regulators. While the interaction effects of BA and NAA on seed germination in MS or 1/2 MS medium both of them showed a fold line (Figures 2(A) and (B)). The results showed that the media with low concentration of salt ions is beneficial to asymbiotic seed germination of orchid, and interactions of plant hormones is found in this stage.
3.2. Development of Multiple Shoots
In order to determine the optimal medium for development of multiple shoots, 20 different kinds of medium were analyzed. The Table 2 showed that the 2.5 mg·L−1 + NAA 0.5 mg·L−1 + 8% BP culture medium was most effective on multiple shoots, up to 19.8 shoots/explant,
 (A)
(A)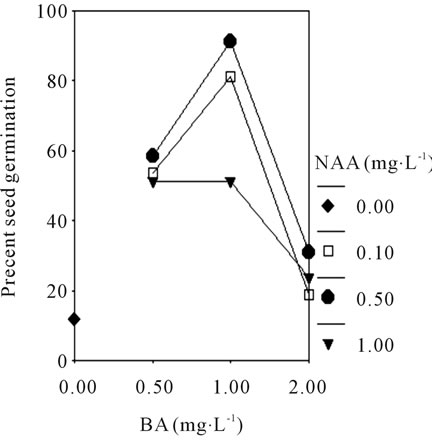 (B)
(B)
Figure 2. The profile plots of seed germination under interaction effect between BA and NAA in MS (A); The profile plots of seed germination under interaction effect between BA and NAA in 1/2 MS (B).
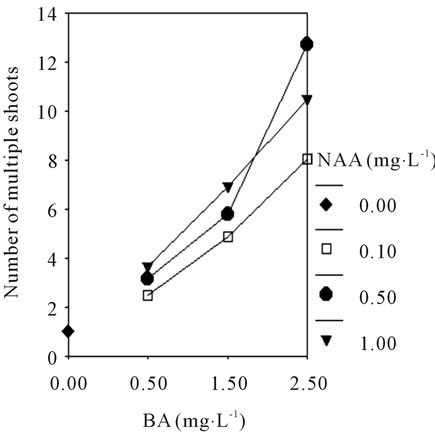
and there was significantly difference compared to others treatment (P ≤ 0.05). Followed by 1/2 MS + BA 2.5 mg·L−1 + NAA 0.1 mg·L−1 + 8% BP (17.6 shoots/explant) and MS + BA 2.5 mg·L−1 + NAA0.5 mg·L−1 + 8% BP (12.7 shoots/explant), the lowest culture medium (1.03 shoots/explant) was in MS + 8% BP. While the plant growth was robust and the seedling division was strong on the 1/2 MS medium supplemented with different plant growth regulators. The Figures 3(A) and (B) shown that interaction effect between BA (0.5, 1.5, and 2.5 mg·L−1) and NAA (0.1, 0.5, and 1.0 mg·L−1) are significantly different in No. of multiple shoots of C. lowianum in MS or 1/2 MS basic medium. The results indicated that the media with low concentration of salt ions is beneficial to development of multiple shoots, and the plant growth regulators NAA and BA play an important role during in vitro propagation of multiple shoots.
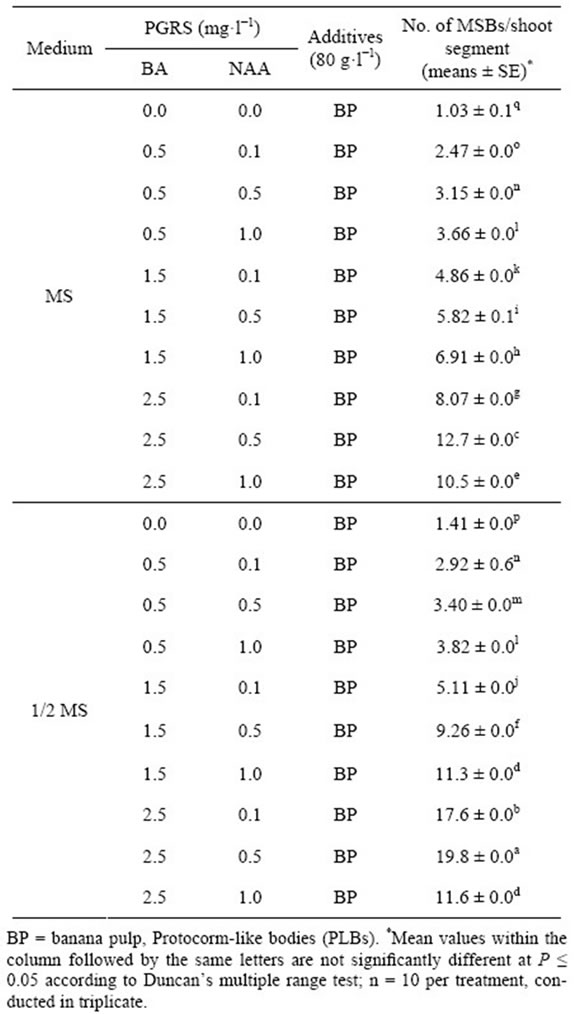
Table 2. Development of multiple shoot buds (MSBs) in C. lowianum after 60 d from Old PLBs on agar-solidified 1/2 MS medium supplemented with BA and NAA.
3.3. Rooting of Shoots
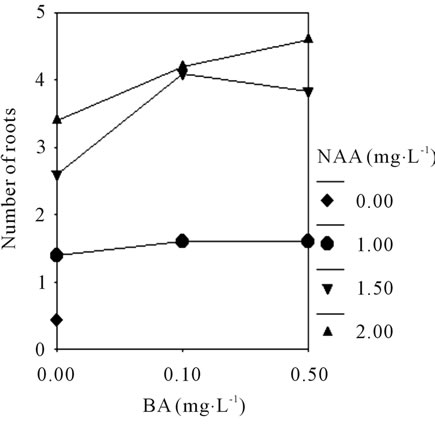
To better understand the rooting of shoots response to different basic medium and the plant growth regulators, the rooting efficiency was investigated (Table 3). The result showed that the rooting efficiency was increased after treatment with the concentration of NAA from 0.0 to 2.0 mg·L−1, supplemented with BA 0.5 mg·L−1, the higher numbers of roots (4.6 per shoot) formed in MS medium. A maximum value (4.7 per shoot) was observed in 1/2 MS + BA 0.1 mg·L−1 + NAA 1.5 mg·L−1 + 0.3% AC (Figure 1(I)). There was no significant difference between the two groups (P > 0.05). However, the lack of BA caused the failure to produce healthy roots (<3.5 per shoot) in the mediums. While there were fold lines (Figures 4(A) and (B)), shown that interaction effect between
 (A)
(A)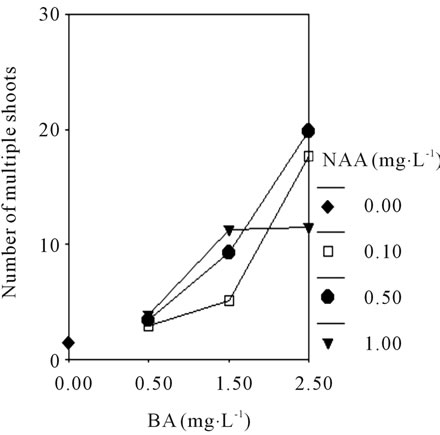 (B)
(B)
Figure 3. The profile plots of No. of multiple shoots under interaction effect between BA and NAA in MS (A); The profile plots of seed germination under interaction effect between BA and NAA in 1/2 MS (B).
BA (0.0, 0.1, and 0.5 mg·L−1) and NAA (1.0, 1.5, and 2.0 mg·L−1) are significantly different in No. of roots of C. lowianum in MS or 1/2 MS basic medium. The change of the rooting efficiency in different medium suggested that the plant growth regulators very important in root induction.
3.4. Ex Vitro Establishment
After transplanting thirty days, the Figure 5 shown that the survival rate was established at 92% in moss (Figure 1(J)), 67% in leaf litter, 60% in ceramisite and tree shell, and 14% in sand. The moss was best for growth of C. lowianum, and there were significantly difference compared to others treatment (P ≤ 0.05). After 90 days, each plant was transplanted individually in pot a filled with a mixture of black peat and tree shell (1:1) (Figure 1(K)) (data not shown).
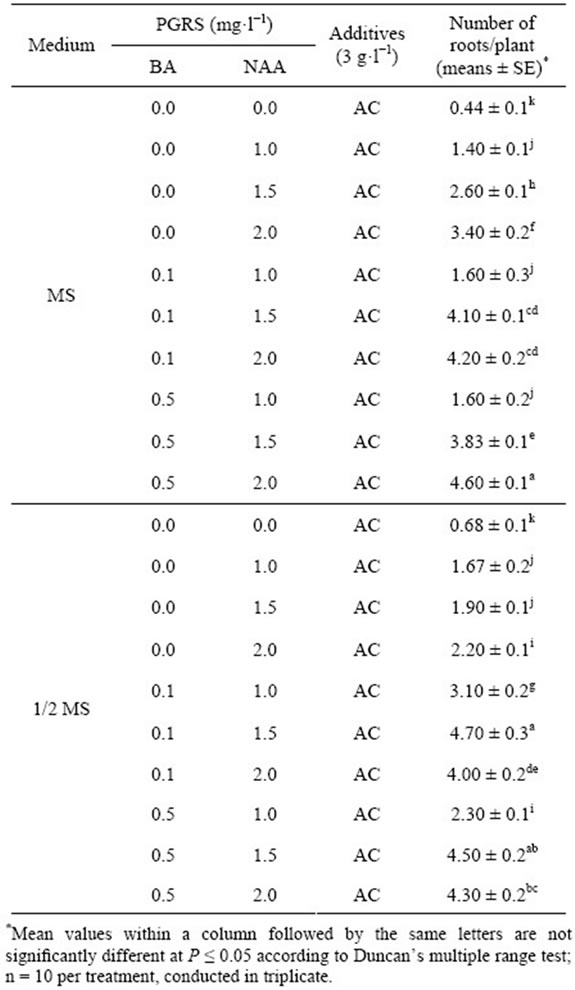
Table 3. Rooting response in multiple shoot buds (MSBs) of C. lowianum after 30 d.
4. Discussion
Many studies on the Orchidaceae indicate that viability testing is often not a good indicator of germinability [8-10], and that the validity of viability estimates should be confirmed with germination tests. Thus, seed viability was not tested in the present study. In vitro seed germination has been suggested as a suitable propagation method for conservation of orchids [11-16]. A number of studies have focused on determining the nutritional requirements for optimal germination and early seedling development in vitro. Rasmussen (1995) reported 40 types of media as being capable of sustaining orchid germination in vitro, and that the beneficial modifications included reduced concentration of inorganic salt [17,18]. In the present study, the media used yielded significantly different (P ≤ 0.05) results in seed germination, and multiple shoots and roots; a higher percentage of
 (A)
(A)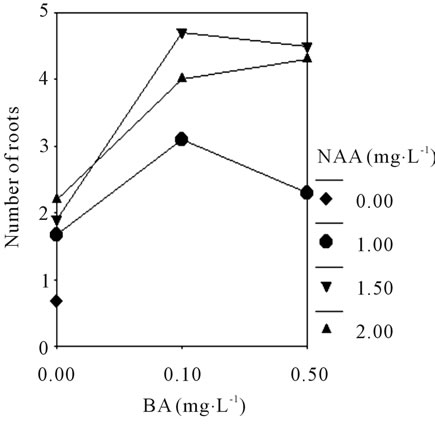 (B)
(B)
Figure 4. The profile plots of No. of roots/plant under interaction effect between BA and NAA in MS (A); The profile plots of seed germination under interaction effect between BA and NAA in 1/2 MS (B).
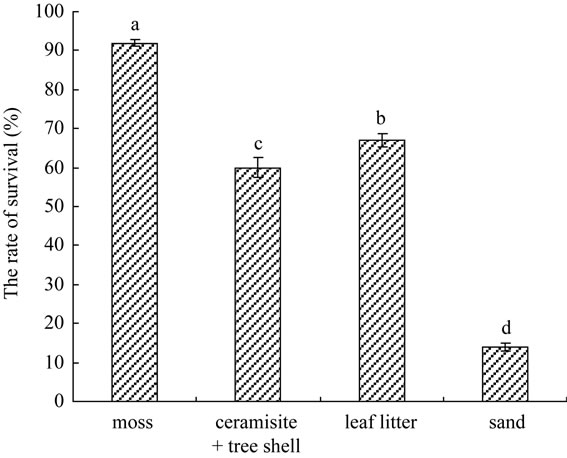
Figure 5. Effect of different matrix type on rate of survival of seedling. Mean totals with a common letter are not different (P ≤ 0.05) by ANOVA on ranks test.
seed germination, and multiple shoots and roots were observed in 1/2 MS medium. The same medium was also beneficial for the Cymbidium tracyanum seed [19].
The type and concentrations of plant growth regulators play an important role during in vitro propagation of many orchid species [20]. Interactions of plant hormones have been found in many cases [21]. Recently, some genes that encode transporters of glutathione-conjugate were isolated, and the nature of the transport of secondary metabolites has began to become clear [22]. Because the orchid seed is tiny, does not have the endosperm, it is very difficult to sprout under natural condition. In our laboratory, the researchers have detected that the plant growth regulators can accelerate the germination of orchid seeds, yet the mechanism is still not clear. Consequently, growth regulators (NAA and BA) were used in the during seed germination, yielding multiple shoots and roots in MS or 1/2 MS medium in the present study. The results indicate that there were interaction effects between NAA and BA, while the lack of BA failed to produce healthy roots (<3.5 roots). This could be because hormones interact differently with the other hormones in each organ, and hormone interactions can be dramatically affected by organ type [23]. NAA and BA were the most suitable choices for tissue culture not only because they was cheaper than others Zeatin (ZT), but also because their active ingredients do not break down under high temperature (121˚C). Thus, the application of NAA and BA during production is a viable technique to reduce production cost.
During the culture period, the exudation of phenol was very common and often influenced the result. The addition of AC to medium also increased the frequency of subculturing, which considerably reduced this problem [24]. Manjula et al. (1997) noted that 0.05 g/l AC was sufficient to reduce polyphenol exudation from A. indica. Phenol exudation from the ex-plant is a major constraint in Ensete ventricosum tissue culture [25]. Birmeta and Welander (2004) overcame this problem by adding 0.05 - 0.1 g·L−1 AC together with dark incubation of the cultures [26]. Nguyen et al. (2007) tried to eradicate this problem in sorghum by using AC in the medium. AC (1 - 5 g·L−1) had a positive effect on reducing the black pigments released into the media by immature embryos [27]. AC induced shoot growth and rooting has also been reported in short leaf pine (Pinus spp.) by Jang and Tainter (1991) [28]. In the present study, the exudation of phenol did not appear and induced shoot rooting with adding 3 g·L−1 AC, but there was appear the exudation of phenol without adding AC to medium during the period of multiple shoots (data not shown), this change may be related to AC. In this study, the proliferation of buds reached 19.8 in the 1/2 MS medium with adding 80 g·L−1 BP in the proliferative phase. Islam et al. (2000) observed that the incorporation of banana-enhanced germination and growth of Cattleya seedlings was relative to control [29]. Van Staden and Stewart (1975) suggested that cell division-inducing compounds present in banana fruit may be responsible for its enhancing effect on orchid embryo development and differentiation [30].
In the present study, discovered seedling discoloration, from seed germination with the addition of 3 g·L−1 AC to the proliferation addition of 80 g·L−1 BP of culture medium a month after subculture, with even some of the leaves of the seedlings showing white (Figures 1(F) and (G)). In orchid Rhynchostylis retusa, the in vitro seedling growth was promoted by the addition of AC in the medium. Seedlings were discolored on MS medium supplemented with BA, NAA, and 0.5 g·L−1 AC; however, seedling color was normal in the same medium with 1 g·L−1 AC. The multiple shoots grew green on MS medium fortified with 1-phenyl-3-(1, 2, 3-thiadiazol-5-yl)- urea (TDZ) and 1 g·L−1 AC [31]. In this experiment, a portion of the seedling was discolored on MS or 1/2 MS medium supplemented with BA, NAA, and lack of AC. However, the discoloration did not emerge during seed germination or root development on MS or 1/2 MS medium supplemented with BA, NAA, and 3 g·L−1 AC. This shows there is a correlation between seedling discoloration and a certain range of AC concentration, but the relationship between the concentration of AC and discolored seedling needed further study.
5. Acknowledgements
We thank the National Science Foundation of China (Nos. 30160074, 30760155) Foundation of Education of China (No. Z2005-2-65008) and the Yunnan Natural Science Fund (No. 2009BB013, No 2012BB008) for the financial support of this research.
REFERENCES
- J. Y. Chen, “The Top Ten Most Known Flowers of China,” Shanghai Culture Publishing House, Shanghai, 1989.
- J. D. Chung, C. K.Chun and S. O. Choi, “Asymbiotic Germination of Cymbidium ensifolium 2; Effects of Several Supplements to the Medium, pH Values and Light and/or Dark Culture Periods on Growth of Rhizome and Organogenesis for Rhizome,” Journal of The Korean Society for Horticultural Science, Vol. 26, No. 2,1985, pp. 186-192.
- K. Shimasaki and S. Uemoto, “Micropropagation of a Terrestrial Cymbidium Species Using Rhizomes Developed from Seeds and Pseudo Bulbs,” Plant Cell, Tissue and Organ Culture, Vol. 22, No. 3, 1990, pp. 237-244. doi:10.1007/BF00033642
- X. Wang, “Tissue Culture of Cymbidium: Plant and flower Induction in Vitro,” Lindleyana, Vol. 3, 1988, pp. 184-189.
- N. R.Nayak, M. Tanaka and J. A. Teixeira da Silva, “Biotechnology of Cymbidium—An Overview of Recent Progress and Future Opportunities,” In: J. A. Teixeira da Silva, Ed., Floriculture, Ornamental and Plant Biotechnology: Advances and Topical Issues, Vol. 4, Global Science Books, London, 2006, pp. 558-562.
- R. B. Malabadi, J. A. Teixeira da Silva, K. Nataraja and G. S. Mulgund, “Shoot Tip Transverse Thin Cell Layers and 2,4-Epibrassinolide in the Micropropagation of Cymbidium bicolor Lindl,” Floriculture and Ornamental Biotechnol, Vol. 3, 2008, pp. 44-48.
- D. T. Nhut, T. N. T. Tien, M. T. N. Huong, N. T. H. Hien, P. P. X. Huyen, V. Q. Luan, B. V. Le and S. Teixeira da, “Artificial Seeds for Preservation and Propagation of Cymbidium spp,” Prop Ornamental Plants, Vol. 5, 2005, pp. 67-73.
- D. Lauzer, M. S. Arnaud and D. Barabe, “Teterazolium Straining and in Vitro Germination of Mature Seeds of Cypripedium acaule (Orchidaceae),” Lindeleyana, Vol. 9, 1994, pp. 197-204.
- V. Vujanovic, S. M. Arnaud, D. Barabe and G. Thibeault, “Viability Testing of Orchid Seed and the Promotion of Coloration and Germination,” Annals of Botany, Vol. 86, No. 1, 2000, pp. 79-86. doi:10.1006/anbo.2000.1162
- T. R. Johnson, S. L. Stewart, D. Dutra, M. K. Kane and L. Richardson, “A Symbiotic and Symbiotic Seed Germination of Eulophia alta (Orchidaceae)—Preliminary Evidence for the Symbiotic Culture Advantage,” Plant Cell, Tissue and Organ Culture, Vol. 90, No. 3, 2007, pp. 313- 323.
- J. Arditti, J. D. Michaud and A. P. P. Oliva (1981), “Seed Germination of North American Orchids. I. Native California and Related Species of Calypso, Epipactis, Goodyera, Piperia and Plantantheria,” Botanical Gazette, Vol. 142, No. 4, 1981, pp. 442-453. doi:10.1086/337245
- A. Thornhill and H. Koopowitz, “Viability of Disa Uniflora Berg (Orchidaceae) Seeds under Variable Storage Conditions: Is Orchid Gene Banking Possible?” Biological Conservation, Vol. 62, No. 1, 1992, pp. 21-27. doi:10.1016/0006-3207(92)91148-L
- L. W. Zettler and T. M. McInnis, “Symbiotic Seed Germination and Development of Spiranthes cernua and Goodyera pubescens (Orchidaceae: Spiranthoideae),” Lindleyana, Vol. 8, 1993, pp. 155-162.
- P. P. J. Kauth, W. A. Vendrame and M. E. Kane, “In Vitro Seed Culture and Seedling Development of Calopogon tuberosus,” Plant Cell, Tissue and Organ Culture, Vol. 85, No. 1, 2006, pp. 91-102. doi:10.1007/s11240-005-9055-1
- S. L. Stewart and M. K. Kane, “A Symbiotic Seed Germination and in Vitro Seedling Development of Habenaria macroceratitis (Orchidaceae) a Rare Florida Terrestrial Orchid,” Plant Cell, Tissue and Organ Culture, Vol. 86, No. 2, 2006, pp. 147-158. doi:10.1007/s11240-006-9098-y
- G. Mahendran and V. Narmatha Bai, “Mass Propagation of Satyrium nepalense D. Don.—A Medicinal Orchid via Seed Culture,” Scientia Horticulturae, Vol. 119, No. 2, 2009, pp. 203-207. doi:10.1016/j.scienta.2008.07.029
- R. Mitchell, “Growing Hardy Orchids from Seeds at Kew,” Plantsman, Vol. 1, 1989, pp. 152-169.
- A. B. Anderson, “Symbiotic and Asymbiotic Germination and Growth of Spiranthes magnicamporum (Orchidaceae),” Lindleyana, Vol. 6, 1991, pp. 183-186.
- Z. L. Li, C. X. Yu, Y. Y. Wang and B. Q. Wang, “Research on Aseptic Budding and Fast Propagation of the Cymbidium tracyanum Seed,” Chinese Agricultural Science Bulletin, Vol. 21, No. 8, 2005, pp. 269-281.
- V. L. Sheela, S. Sarada and S. Anita, “Micropropagation of Orchids,” Journal of Ornamental Horticulture, Vol. 9, No. 1, 2006, pp. 1-11.
- P. P. D. Davies, “The Plant Hormones: Their Nature, Occurrence and Functions,” Plant Hormones, Springer, 2010, pp. 1-15.
- K. A. Marrs, W. R. Alfenito, A. M. Lloyd and V. Walbot, “A Glutathione S-Glucosyltransferase Involved in Vacuolar Transfer Encoded by the Maize Gene Bronze-2,” Nature, Vol. 375, No. 6530, 1995, pp. 397-400. doi:10.1038/375397a0
- J. Ross and Damian O’Neill (2001), “New Interactions between Classical Plant Hormones,” Plant Science, Vol. 6, No. 1, 2001, pp. 2-4. doi:10.1016/S1360-1385(00)01795-7
- E. V. Soniya and M. Sujitha, “An Efficient in Vitro Propagation of Aristolochia indica,” Biologicl Plantarum, Vol. 50, No. 2, 2006, pp. 272-274. doi:10.1007/s10535-006-0018-0
- S. Manjula, A. Thomas, B. Daniel and G. M. Nair, “In Vitro Plant Regeneration of Aristolochia Indica through Axillary Shoot Multiplication and Organogenesis,” Plant Cell Tissue Organ Culture, Vol. 51, No. 2, 1997, pp. 145- 148. doi:10.1023/A:1005978125424
- G. Birmeta and M. Welander, “Efficient Micropropagation of Ensete Ventricosum Applying Meristem Wounding: A Three-Step Protocol,” Plant Cell Reports, Vol. 23, No. 5, 2004, pp. 277-283. doi:10.1007/s00299-004-0832-9
- T. V. Nguyen, T. T. Thu, M. Claeys and G. Angenon, “Agro Bacterium Mediated Transformation of Sorghum (Sorghum bicolor (L.) Moench) Using an Improved in Vitro Regeneration System,” Plant Cell, Tissue and Organ Culture, Vol. 91, No. 2, 2007, pp. 64-155.
- J. C. Jang and F. H. Tainter, “Micropropagation of Short Leaf, Virginia and Loblolly Short Leaf Pine Hybrids via Organogenesis,” Plant Cell, Tissue and Organ Culture, Vol. 25, No. 1, 1991, pp. 57-61. doi:10.1007/BF00033914
- M. O. Islam, S. Matsui and S. Ichihashi, “Effects of Complex Organic Additives on Seed Germination and Carotenoid Content in Cattleya Seedlings,” Lindleyana, Vol. 15, 2000, pp. 81-88.
- J. VanStaden and J. Stewart, “Cytokinins in Banana Fruit,” Zeitschrift für Pflanzenphysiologie, Vol. 76, No. 3, 1975, pp. 280-283.
- T. D. Thomas and A. Michael, “High-Frequency Plantlet Regeneration and Multiple Shoot Induction from Cultured Immature Seeds of Rhynchostylis retusa Blume, an Exquisite Orchid,” Plant Biotechnology Reports, Vol. 1, No. 1, 2007, pp. 243-249. doi:10.1007/s11816-007-0038-z
NOTES
*Corresponding author.