American Journal of Plant Sciences
Vol.05 No.11(2014), Article ID:46102,7 pages
10.4236/ajps.2014.511183
Effect of Various Intensities of Drought Stress on δ13C Variation among Plant Organs in Rice: Comparison of Two Cultivars
Mana Kano-Nakata1,2, Jiro Tatsumi3, Yoshiaki Inukai1,2, Shuichi Asanuma2, Akira Yamauchi1*
1Graduate School of Bioagricultural Sciences, Nagoya University, Aichi, Japan
2International Corporation Center for Agricultural Education (ICCAE), Nagoya University, Aichi, Japan
3Center for Bioresource Field Science, Kyoto Institute of Technology, Kyoto, Japan
Email: *ayama@agr.nagoya-u.ac.jp
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 7 April 2014; revised 6 May 2014; accepted 17 May 2014
ABSTRACT
The δ13C value is widely used to assess the effects of drought on water status in plants. However, there is little information regarding the δ13C signature in different organs of rice. We conducted a field study to examine whether the δ13C among different plant parts would be affected by the intensities of drought, and to evaluate genotypic variation in δ13C fluctuation among plant parts affected by drought intensities. Two cultivars, “Nipponbare” (Oryzasativa ssp. japonica) and “Kasalath” (O. sativa ssp. indica), were grown in the field with a line-source sprinkler system. The δ13C values of panicles, flag leaves, straws, culms, and roots were measured from plant samples. The δ13C value increased as drought stress increased, especially in the panicles and roots. “Nipponbare” showed higher values of δ13C than “Kasalath” under the well-watered and mild drought stress conditions, but there was no significant difference between the genotypes in the δ13C value under the severe drought stress condition. The variation in δ13C value among different plant parts was also increased with increasing drought stress. In contrast, these variations were small under well-watered conditions. Furthermore, there was much greater variation in the δ13C value among different plant parts in “Kasalath” than in “Nipponbare” when the plants were grown under drought stress conditions. A significant negative relationship was observed between the δ13C value of panicles and shoot dry matter production, suggesting that the δ13C value of panicles may be the best indicator of plant water status in rice.
Keywords:
δ13C, Drought, Plant Parts, Rice

1. Introduction
Drought is the major limitation to crop yields. The stable carbon isotope ratio (δ13C) or carbon isotope discrimination (CID), which corrects for variations in source air 13C in plants, is a useful indicator to evaluate the impact of water stress on crop performance and to estimate water use efficiency (WUE) [1] [2] . It has been suggested by some authors that δ13C or CID could be an effective selection criterion for grain yield under drought [3] [4] . In general, water stress increased δ13C and decreased CID values in rice [5] - [8] and other crops, such as wheat [4] [9] , barley [10] , chickpea [11] and soybean [12] . Genotypic variation has been reported for δ13C or CID values in rice; the genotypes for the japonica subspecies showed higher δ13C values or lower CID values than the indica genotypes did [13] - [16] . Kondo et al. [5] also reported that CID values varied less in response to water regimes than they did between different genotypes in pot experiments. However, the impacts of the different intensities of drought stress may differ among different genotypes in the field. Therefore, special attention is required to improve understanding of genetic responses of δ13C under various intensities of drought, which is considered to occur in the real field.
It is well known that significant differences in δ13C may exist among different organs with the lower value of δ13C in assimilating organs, such as leaves [17] [18] . Although understanding the δ13C fluctuation among organs should be important, most of the studies that have been conducted measured δ13C of a specific organ or of the whole plant, rather than comparing its value between organs [7] [8] [14] - [16] . Recently, much attention has been focused on the differences in δ13C between plant organs in relation to carbon metabolism [19] - [21] , but information on δ13C in different organs is not well documented in rice. In addition, the effect of soil water conditions on the variation of 13C among plant organs has not been examined in the field.
We previously used chromosome segment substitution lines derived from “Nipponbare” (japonica) and “Kasalath” (indica) crosses, and found that plant response in their shoot and root growth to various intensities of drought stress was different among genotypes under field conditions [22] . In that study, significant genotypic difference in dry matter production was observed among parent genotypes under drought stress conditions. “Kasalath” produced more shoot dry matter than “Nipponbare” through the greater photosynthetic rate and stomatal conductance due to the greater root development especially under mild drought stress conditions.
Thus, we hypothesized that the differences in δ13C values among different plant parts might be affected by drought intensities and there might be genotypic variation in δ13C fluctuation among plant parts affected by drought intensities. To examine these hypotheses, we conducted field experiment using “Nipponbare” and “Kasalath”.
2. Material and Methods
2.1. Experimental Design
The field experiment was conducted at the experimental farm at Nagoya University, Nagoya, Japan (lat. 35˚6ʹ42"N, long. 137˚4ʹ57ʺE) during the summer, 2006. “Nipponbare” (japonica) and “Kasalath” (indica), which differed in their shoot growth and root development in our previous study [22] , were grown in a watertight experimental bed with a line source sprinkler system under a rain-out shelter to create a gradient in soil moisture, previously described [22] .
Twenty-five-day-old seedlings of each cultivar were transplanted perpendicular to the water pipe so that they received different amounts of water. Each genotype was replicated for three rows along a 1.8 m row and spaced within a 45 × 20 cm area. The placements were randomized. Each row contained eight plants of a given genotype that received different amounts of water from the line source; the closest one received the most and the furthest one the least amount of water. The fields were kept well-watered by sprinkler irrigation for one week after transplanting during the period of establishment, after which drought stress was imposed. The soil moisture content (SMC) at a depth of 10 cm at the base of each plant in every line was measured by using Hydrosense (Decagon, Utah, USA) in % v/v, and then converted into the value in % w/w. Figure 1 shows that the line source sprinkler system successfully created and maintained a soil moisture gradient ranging from 5% to 40% w/w of SMC, which is equivalent to a water potential of −0.163 MPa to 0 MPa.
2.2. Measurements of Gas Exchange
Several physiological traits―photosynthetic rate (Pn), transpiration rate, and stomatal conductance (gs)―were

Figure 1. Relationships between shoot dry weight (a), photosynthetic rate (Pn) (b), stomatal conductance (gs) (c), and water use efficiency (WUE) (d) and soil moisture content for “Nipponbare” and “Kasalath”.
measured with a portable photosynthesis analyzer (LI-6400, LI-COR, USA) in the morning at 56 - 58 days after transplanting.
2.3. Plant Sampling and Analysis
Plants were harvested 70 days after transplanting. Harvested plants were separated into panicles (above the panicle base), flag leaves, straws (aboveground parts without panicles, flag leaves and culms), culms (5 cm long from the ground surface), and roots. All samples were wrapped with aluminum foil and immediately frozen in liquid nitrogen. Plant samples were oven dried at 70˚C for two days and the dry weights were measured. Dried samples were milled into a fine powder using a sample mill (MF10, IKA, Germany). The δ13C values of samples were analyzed by an isotope mass spectrometer (Finnigan MAT GmbH, Germany). The results of δ13C analysis were expressed in per mil using the following equation:
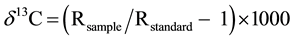
where R refers to 13C/12C and standard refers to the international Pee Dee Belemnite standard (a fossil Belemnite from the Pee Dee formation in South Carolina). Measurements were performed with three repetitions for each sample.
2.4. Statistical Analysis
Fisher’s Least Significant Difference (LSD) test was used for mean comparison, and differences were considered significant at p < 0.05. The relationship between shoot dry weight and δ13C values in different plant parts were determined using regression analysis.
3. Results
Both “Nipponbare” and “Kasalath” apparently reduced their production of shoot dry matterwith increasing drought stress. “Kasalath” showed higher shoot dry weight than “Nipponbare” at any SMC (Figure 1(a)). Similar trends were observed in Pn and gs with reduction of those values for both genotypes as drought stress increased (Figure 1(b), Figure 1(c)). However, there was no effect of drought stress on WUE for either cultivar (Figure 1(d)).
Figure 2 shows the effects of various SMC on the δ13C in different plant parts for “Nipponbare” and “Kasalath”. The δ13C value of the shoot tended to increase with increasing drought stress, and the variations in δ13C in different plant parts were more pronounced as drought intensified in both “Nipponbare” and “Kasalath”. The effect of drought stress on δ13C fluctuation among plant parts was greater in “Kasalath” than in “Nipponbare”. In “Kasalath”, the δ13C value among plant parts ranged from −26.83‰ (panicle) to −30.78‰ (flag leaf), whereas in “Nipponbare” it ranged from −27.09‰ (panicle) to −29.69‰ (straw). Thus “Kasalath” showed 1.5 times larger fluctuation in 13C due to drought stress as compared with “Nipponbare”.
To examine the effects of various intensities of drought stress on the δ13C value, the SMC was divided into the following three categories: the well-watered (>30% w/w of SMC), the mild drought stress (15% - 30% w/w of SMC) and the severe drought stress (<15% w/w of SMC) [22] in Figure 3. At the well-watered, the δ13C value of plant parts ranged within 0.6‰ from −29.7‰ (straw) to −29.1‰ (culm) in “Nipponbare”, and within 0.5‰ from −31.7‰ (panicle) to −31.2‰ (culm) in “Kasalath”. Genotypic variation was observed in δ13C with significantly higher δ13C value in “Nipponbare” than in “Kasalath” at well-watered condition. At the mild drought stress, the δ13C value of plant parts ranged within 0.6‰ from −29.4‰ (flag leaf) to −28.8‰ (culm) in “Nipponbare”, and within 1.2‰ from −31.0‰ (flag leaf) to −29.8‰ (culm) in “Kasalath”. The δ13C value was also significantly higher for “Nipponbare” than for “Kasalath” in any of plant parts except culms at mild drought stress condition. At the severe drought stress, the δ13C value of plant parts ranged within 0.9‰ from −28.7‰ (straw) to −27.8‰ (panicle) in “Nipponbare”, and showed greater variation of 3.0‰, ranging from −30.2‰ (flag leaf) to −27.2‰ (panicle) in “Kasalath”. Therefore, we confirmed statistically that drought effect on δ13C fluctuation among plant parts was greater in “Kasalath” than in “Nipponbare”. In contrast, the δ13C value did not significantly vary with the genotype under severe drought stress condition. Among the plant parts, panicles showed the greatest change in δ13C induced by drought stress in both cultivars. In “Nipponbare”, the δ13C values of panicles and roots significantly increased in severe drought stress conditions over the values in well-watered conditions, but there was no significant difference in the δ13C value of panicles and roots between mild drought and well-watered conditions. However, “Kasalath” showed a significantly higher δ13C value in panicles and roots in response not only to severe stress but also to mild drought stress. In contrast, the δ13C values in flag leaves did not significantly increase in response to drought in either genotype. On the other hand, the effects of soil moisture on the δ13C value of culm and straw were significant only in “Kasalath”.


Figure 2. Relationships between δ13C values of plant parts and soil moisture content for “Nipponbare” (a) and “Kasalath” (b).


Figure 3. Differences in δ13C values among plant parts under well-watered (>30% w/w of SMC), mild drought (15% - 30% w/w of SMC), and severe drought (<15% w/w of SMC) conditions for “Nipponbare” (a) and “Kasalath” (b). Within the same plant parts, values followed by the same letter genotype are not significantly different at the 5% level by Fisher’s LSD test. between 13C values of plant parts and soil moisture content for “Nipponbare” (a) and “Kasalath” (b).
Correlation coefficient among shoot dry weight and δ13C values of plant parts is shown in Table 1. The δ13C values of panicles and roots were significantly correlated with shoot dry matter production in both cultivars and highest value of correlation coefficient was found between δ13C in panicles and shoot dry weight. In contrast, there was no correlation between the δ13C in flag leaf and shoot dry weight in either genotype. The δ13C values of culm and straw were significantly correlated with dry matter production in “Kasalath”. In addition, correlation coefficients between δ13C value and shoot dry matter production were higher in “Kasalath” than in “Nipponbare” in any of plant parts except flag leaves.
4. Discussion
As shown in Figure 1, the drought-induced decrease in biomass production was largely associated with reduced Pn in both cultivars. This decrease could be explained by reductions in gs. On the other hand, WUE remained relatively stable at all levels of SMC, and WUE was not highly correlated with shoot dry matter production in this study (data not shown). Water uptake capacity may be more significant than WUE in determining biomass production under drought conditions, as reported by previous studies [23] [24] .

Table 1. Correlation coefficient among shoot dry weight and δ13C values of plant parts.
* and ** indicate significant correlation at the 5% and 1% level, respectively.
The general trend of increasing 13C in the shoot with drought stress found in the present study agrees with the results of previous studies on rice [5] - [8] . A tendency for higher values of δ13C (lower CID) to appear in the japonica genotype than in the indica genotype shown in the present results also was consistent with previous reports [13] - [16] . Under the well-watered and mild drought stress conditions, higher δ13C values of the shoot in “Nipponbare” indicate that “Nipponbare” assimilated CO2 more enriched in 13C than “Kasalath”, and this accords with that Pn and gs in Nipponbare was lower than Kasalath. However, such genotypic variation in δ13C was not observed in any of the individual plant parts under the severe drought stress condition in this study (Figure 3). This finding indicates that the cultivar effects on δ13 C values become less pronounced as drought stress intensifies. Therefore, the differences in δ13 C value between drought stress and non-stressed conditions are critical.
There was a general trend toward enrichment in 13C in grains and roots, compared with flag leaves and stems, in previous studies on rice [5] [6] [13] . In this study, a similar trend in δ13C variation among different plant parts was also observed, but such variations were affected by SMC and by genotypes (Figure 2, Figure 3). Drought had stronger effects on δ13C levels in panicles and roots than in other plant parts (Figure 3). In the present study the panicles and roots were the prominent sink for photo-assimilated carbon. The translocation of photo-assimilated carbohydrates from leaves to sink organs may be largely prevented by drought stress. The difference in carbon isotope composition among plant parts is related to the differences in fractionation processes during transport, the synthesis of metabolites, and the chemical composition of different organs, such as the amounts of lipids and lignin [25] . However, the reason for these differences is not clear.
Interestingly, the variance in δ13C value among different plant parts was much higher in “Kasalath” than in “Nipponbare” when the plants were grown under drought stress conditions (Figure 2, Figure 3). “Kasalath” is considered to be well adapted to mild drought stress conditions, compared with “Nipponbare”, in our previous studies [22] [26] . Therefore, the δ13C value in response to mild drought may be related to plant adaptations to such stress conditions.
The stable isotope 13C technique has been used for screening plants with higher tolerance under drought stress conditions. In rice (Oryza sativa), putative quantitative trait loci were detected for δ13C or CID values [13] -[16] . So far, despite various efforts, δ13C values have not been used to produce drought-resistant varieties of any crop. In this study, the δ13C value of panicles was better correlated with shoot dry matter production than was the δ13C of other plant parts in both “Nipponbare” and “Kasalath” (Table 1). This result indicates that the δ13C value of panicles may be the best indicator of plant water status in rice, although further research is needed to evaluate how δ13C variation in different plant parts can related to plant adaptation to drought stress in rice.
Acknowledgements
This research was supported by a Grant-in-Aid for Scientific Research (No. 22380013) and a Grant-in-Aid for JSPS Fellows (No. 25040128) from the Japan Society for the Promotion of Science, and partially supported by the Japan Science and Technology Agency (JST)/Japan International Cooperation Agency (JICA), the Science and Technology Research Partnership for Sustainable Development (SATREPS).
References
- Farquhar, G.D. and Richards, R.A. (1984) Isotope Composition of Plant Carbon Correlates with Water-Use Efficiency of Wheat Genotypes. Australian Journal of Plant Physiology, 11, 539-552. http://dx.doi.org/10.1071/PP9840539
- Farquhar, G.D., Ehleringer, J.R. and Hubick, K.T. (1989) Carbon Isotope Discrimination and Photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology, 40, 503-537. http://dx.doi.org/10.1146/annurev.pp.40.060189.002443
- Condon, A.G., Richards, R.A., Rebetzke, G.J. and Farquhar, G.D. (2002) Improving Water-Use Efficiency and Crop Yield. Crop Science, 42, 122-131. http://dx.doi.org/10.2135/cropsci2002.0122
- Xu, X., Yuan, H., Li, S., Trethowan, R. and Monneveux, P. (2007) Relationship between Carbon Isotope Discrimination and Grain Yield in Spring Wheat Cultivated under Different Water Regimes. Journal of Integrative Plant Biology, 49, 1497-1507. http://dx.doi.org/10.1111/j.1672-9072.2007.00562.x
- Kondo, M., Pablico, P.P., Aragones, D.V. and Agbisit, R. (2004) Genotypic Variations in Carbon Isotope Discrimination, Transpiration Efficiency, and Biomass Production in Rice as Affected by Soil Water Conditions and N. Plant and Soil, 267, 165-177. http://dx.doi.org/10.1007/s11104-005-4884-6
- Zhao, B., Kondo, M., Maeda, M., Ozaki, Y. and Zhang, J. (2004) Water-Use Efficiency and Carbon Isotope Discrimination in Two Cultivars of Upland Rice during Different Developmental Stages under Three Water Regimes. Plant and Soil, 261, 61-75. http://dx.doi.org/10.1023/B:PLSO.0000035562.79099.55
- Impa, S.M., Nadaradjan, S., Boominathan, P., Shashidhar, G., Bindumadhava, H. and Sheshshayee, M.S. (2005) Carbon Isotope Discrimination Accurately Reflects Variability in WUE Measured at a Whole Plant Level in Rice. Crop Science, 45, 2517-2522. http://dx.doi.org/10.2135/cropsci2005.0119
- Centritto, M., Lauteri, M., Monteverdi, M.C. and Serraj, R. (2009) Leaf Gas Exchange, Carbon Isotope Discrimination, and Grain Yield in Contrasting Rice Genotypes Subjected to Water Deficits during the Reproductive Stage. Journal of Experimental Botany, 60, 2325-2339. http://dx.doi.org/10.1093/jxb/erp123
- Zhu, L., Li, S.H., Liang, Z.S., Xu, X. and Li, Y. (2009) Relationship between Carbon Isotope Discrimination, Mineral Content and Gas Exchange Parameters in Vegetative Organs of Wheat Grown under Three Different Water Regimes. Journal of Agronomy and Crop Science, 196, 175-184. http://dx.doi.org/10.1111/j.1439-037X.2009.00404.x
- Chen, J., Chang, X.S. and Anyia, A.O. (2013) Physiological Characterization of Recombinant Inbred Lines of Barley with Contrasting Levels of Carbon Isotope Discrimination. Plant and Soil, 369, 335-349. http://dx.doi.org/10.1007/s11104-012-1578-8
- Krishnamurthy, L., Kashiwagi, J., Tobita, S., Ito, O., Upadhyaya, H.D., Gowda, C.L.L., Gaur, P.M., Sheshshayee, M.S., Singh, S., Vadez, V. and Varshney, R.K. (2013) Variation in Carbon Isotope Discrimination and its Relationship with Harvest Index in the Reference Collection of Chickpea Germplasm. Functional Plant Biology, 40, 1350-1361. http://dx.doi.org/10.1071/FP13088
- Bellaloui, N. (2011) Effect of Water Stress and Foliar Boron Application on Seed Protein, Oil, Fatty Acids, and Nitrogen Metabolism in Soybean. American Journal of Plant Sciences, 2, 692-701. http://dx.doi.org/10.4236/ajps.2011.25084
- Laza, M.R., Kondo, M., Ideta, O., Barlaan, E. and Imbe, T. (2006) Identification of Quantitative Trait Loci for δ13Cand Productivity in Irrigated Lowland Rice. Crop Science, 46, 763-773. http://dx.doi.org/10.2135/cropsci2005.05.0084
- Xu, Y., This, D., Pausch, R.C., Vonhof, W.M., Coburn, J.R., Comstock, J.P. and McCouch, S.R. (2009) Leaf-Level Water Use Efficiency Determined by Carbon Isotope Discrimination in Rice Seedlings: Genetic Variation Associated with Population Structure and QTL Mapping. Theoretical and Applied Genetics, 118, 1065-1081. http://dx.doi.org/10.1007/s00122-009-0963-z
- Takai, T., Ohsumi, A., San-oh, Y., Laza, M.R.C., Kondo, M., Yamamoto, T. and Yano, M. (2009) Detection of a Quantitative Trait Locus Controlling Carbon Isotope Discrimination and Its Contribution to Stomatal Conductance in Japonica Rice. Theoretical and Applied Genetics, 118, 1401-1410. http://dx.doi.org/10.1007/s00122-009-0990-9
- This, D., Comstock, J., Courtois, B., Xu, Y., Ahmadi, N., Vonhof, W.M., Fleet, C., Setter, T. and McCouch, S. (2010) Genetic Analysis of Water Use Efficiency in Rice (Oryza sativa L.) at the Leaf Level. Rice, 3, 72-86. http://dx.doi.org/10.1007/s12284-010-9036-9
- Hall, A.E., Mutters, R.G., Hubick, K.T. and Farquha, G.D. (1990) Genotypic Differences in Carbon Isotope Discrimination by Cowpea under Wet and Dry Field Conditions. Crop Science, 30, 300-305. http://dx.doi.org/10.2135/cropsci1990.0011183X003000020011x
- Acevedo, E. (1993) Potential of Carbon Isotope Discrimination as a Selection Criterion in Barley breedIng. In: Ehleringer, J.R., Hall, A.E. and Farquhar, G.D., Eds., Stable Isotopes and Plant Carbon-Water Relations, Academic Press, New York, 399-417.
- Monti, A., Amaducci, M.T., Pritoni, G. and Venturi, G. (2006) Variation in Carbon Isotope Discrimination during Growth and at Different Organs in Sugar Beet (Beta vulgaris L.). Field Crops Research, 98, 157-163. http://dx.doi.org/10.1016/j.fcr.2006.01.002
- Peuke, A.D., Gessler, A. and Rennenberg, H. (2006) The Effect of Drought on C and N Stable Isotopes in Different Fractions of Leaves, Stems and Roots of Sensitive and Tolerant Beech Ecotypes. Plant Cell and Environment, 29, 823- 835. http://dx.doi.org/10.1111/j.1365-3040.2005.01452.x
- Grant, O.M., Davies, M.J., James, C.M., Johnson, A.W., Leinonen, I. and Simpson, D.W. (2012) Thermal Imaging and Carbon Isotope Composition Indicate Variation amongst Strawberry (Fragaria × ananassa) Cultivars in Stomatal Conductance and Water Use Efficiency. Environmental and Experimental Botany, 76, 7-15. http://dx.doi.org/10.1016/j.envexpbot.2011.09.013
- Kano, M., Inukai, Y., Kitano, H. and Yamauchi, A. (2011) Root Plasticity as the Key Root Trait for Adaptation to Various Intensities of Drought Stress in Rice. Plant and Soil, 342, 117-128. http://dx.doi.org/10.1007/s11104-010-0675-9
- Matsunami, M., Matsunami, T., Ogawa, A., Toyofuku, K., Kodama, I. and Kokubun, M. (2012) Genotypic Variation in Biomass Production at the Early Vegetative Stage among Rice Cultivars Subjected to Deficient Soil Moisture Regimes and Its Association with Water Uptake Capacity. Plant Production Science, 15, 82-91. http://dx.doi.org/10.1626/pps.15.82
- Matsunami, M., Matsunami, T., Kon, K., Ogawa, A., Kodama, I. and Kokubun, M. (2013) Genotypic Variation in Nitrogen Uptake during Early Growth among Rice Cultivars under Different Soil Moisture Regimes. Plant Production Science, 16, 238-246. http://dx.doi.org/10.1626/pps.16.238
- Brugnoli, E. and Farquhar, G.D. (2000) Photosynthetic Fractionation of Carbon Isotopes. In: Leegood, R.C., Sharkey, T.D. and Von Caemmerer, S., Eds., Photosynthesis: Physiology and Metabolism, Kluwer Academic Publishers, Dordrecht, 352-434.
- Tran, T.T., Kano-Nakata, M., Takeda, M., Menge, D., Mitsuya, S., Inukai, Y. and Yamauchi, A. (2014) Nitrogen Application Enhanced the Expression of Developmental Plasticity of Root Systems Triggered by Mild Drought Stress in Rice. Plant and Soil, 378, 139-152. http://dx.doi.org/10.1007/s11104-013-2013-5
NOTES

*Corresponding author.