Natural Science
Vol.11 No.06(2019), Article ID:93401,7 pages
10.4236/ns.2019.116020
The Characteristics of Biophotonic Activity Induced by Aspartate May Be Related to the Evolution of Species
Shuangqiong Tan1,2, Chi Xu1,2, Jiapei Dai1,2*
1Wuhan Institute for Neuroscience and Neuroengineering (WINN), South-Central University for Nationalities, Wuhan, China; 2Department of Neurobiology, College of Life Sciences, South-Central University for Nationalities, Wuhan, China
Correspondence to: Jiapei Dai, 
Copyright © 2019 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: May 7, 2019 ; Accepted: June 27, 2019 ; Published: June 30, 2019
ABSTRACT
Glutamate, the most abundant excitatory neurotransmitter in the nervous system, can induce biophotonic activity and transmission in mouse brain slices. As a signaling molecule, aspartate is not considered to be an independent neurotransmitter during the long evolution process, which may be just a co-transmitter or neuromodulator. In the view of structure and physiological similarities of aspartate and glutamate, as well as some differences between them, we attempted to investigate whether aspartate could also induce biophotonic activity in mouse brain slices and its effect characteristics. The ultraweak biophoton imaging system (UBIS) was used to carry out a real-time observation of biophoton activity induced by aspartate in mouse brain slices. It was found that the biophotonic emissions induced by aspartate at different concentrations (12.5 mM, 25 mM and 50 mM) presented concentration-dependent effects and 50 mM aspartate could obviously induce biophoton activities with the characteristic changes of initiation, maintenance, washing and reapplication, which were also different from that induced by 50 mM glutamate as reported before. Considering the species differences in excitatory neurotransmitters, these findings indicate that aspartate-induced biophotonic activity may imply the evolutionary differences in the animal brains.
Keywords:
Biophoton, Biophoton Imaging, Aspartate, Glutamate, Brain Slice

1. Introduction
Aspartate, as an excitatory amino acid, has an important but controversial implication in the central nervous system (CNS), and as a signaling molecule, has a long evolutionary history, for example, it can promote bacterial chemotaxis [1]. Earlier studies found that using K+ or electrical stimulation of the brain slices, there was a Ca2+-dependent release of glutamate, aspartate and γ-aminobutyric acid (GABA) [2 - 4]. Glutamate is currently recognized as a neurotransmitter in the excitatory pathways of CNS in almost all mammals, and GABA is the main inhibitory transmitter. However, the implication of aspartate in the CNS is still not clear as it was only known that aspartate can selectively activate N-methyl-D-aspartate (NMDA) receptor as a role of non-metabolism in the mammalian brains [5], but cannot activate the amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA) receptors because of its low affinity to AMPA receptors [6], so it is difficult to believe that it has the function of regulating the rapid transmission of excitatory synapses in the CNS. Moreover, aspartate is not the transport substrate of vesicular glutamate transporter [7 , 8]. Some synaptic vesicles of hippocampal neuron and synaptic-like microvesicles of pinealocytes exocytose the aspartate and glutamate simultaneously [9], but the release of aspartate and glutamate from hippocampal CA1 synaptosome are regulated by different mechanism [10 , 11]. Therefore, it has been generally believed that aspartate is not considered to be an independent neurotransmitter and maybe just a co-transmitter or neuromodulator [12]. Despite the activity of aspartate in neural synapses, aspartate is an elementary amino acid that can be utilized for the synthesis of protein or be catabolized to take part in Tricarboxylic acid (TCA) cycle. It is shown that cultured astrocytes can uptake the exogenous aspartate, which is then hydrolyzed into intermediate metabolites, such as oxaloacetate, pyruvate, acetyl coenzyme A, taking part in TCA cycle [12 , 13].
Biophoton, also called ultra-weak photon emission (UPE), is present in all organisms including animals, plants, microbe and human beings [14 - 16]. Organisms can spontaneously radiate biophotons in the physiological or pathological conditions. There are plenty of evidences indicating that biophoton is related to neural functions. For example, it was demonstrated that biophoton emissions increased significantly from the right sides of volunteer’s heads when they imagined light in a very dark environment, and the neural activity measured by electroencephalography (EEG) is significantly correlated with the biophoton emissions in the human brain [17]. Our previous study found that glutamate could induce the typical biophoton activities in mouse brain slices [18], which are related to the biophotonic signal transmission [18 , 19]. Because of structural similarity between glutamate and aspartate, we tried to investigate whether aspartate could also induce biophoton activity in neural cells as glutamate does, and to provide new research evidence for further analysis of biophotonic activity and functional importance in the brain.
2. Materials and methods
2.1. Mouse Train Slice Preparation
Adult male Kunming mice (2 - 3 months, 35 ± 5 g) were purchased from Hubei Provincial Laboratory Animal Public Service Center (Wuhan, China) and housed under standard conditions (12-h light/dark cycle, room temperature 18˚C - 25˚C, 40% - 50% humidity) with access to food and water adlibitum. The protocols were approved by the Committee on the Ethics of Animal Experiments of South-Central University for Nationalities. Mice were decapitated and the brain was quickly removed and placed in ice-cold (0˚C - 4˚C) artificial cerebral spinal fluid (ACSF) for about 3 min. ACSF contained (in mM) 2.5 KCl, 125 NaCl, 20 D-glucose, 26 NaHCO3, 1.25 NaH2PO4, 1 MgCl2, and 2 CaCl2; pH 7.6, Osmotic pressure 300 mOsm/L. For the preparation of a standardized coronal brain slice (450 µm thickness) [18], beginning at the rostral part of hippocampus, and then, the brain slices were incubated in ACSF at room temperature (23˚C) for a minimum of 1 h before imaging and continuously bubbling with 95% O2 and 5% CO2.
2.2. Biophoton Imaging and Aspartate Applications
The incubated brain slices were transferred to perfusion chamber in which they were submerged in a perfusion bath of ACSF (100 ml) that was contained in a glass bottle. A mixture of 95% O2 + 5% CO2 was constantly supplied in the ACSF during the perfusion period. The perfusion was maintained through an input micropump and an output micropump (5 ml/min) outside the ultraweak biophoton imaging system (UBIS). The temperature (36.8˚C) of the medium in the perfusion chamber was maintained with an electrical heater.
Biophotonic activities were detected and imaged with the UBIS using an EMCCD camera (C9100-13, Hamamatsu Photonics K. K., Hamamatsu, Japan) in water-cool mode (in this situation, the working temperature at the EMCCD camera can be maintained as low as −90˚C), controlled by an image analysis software (HCImage Version 1.1.3.1, Hamamatsu Photonics K. K., Hamamatsu, Japan). The other setup parameters during imaging were photon detection model 3, 1 × 1 binning, 1200 gain, and cooling mode Max. The specific steps for biophoton detection and imaging were as follows: 1) the brain slices were transferred to a perfusion chamber and kept in complete darkness for about 30 min before imaging to exclude the effects of ambient light; 2) real-time imaging by automatically image capture every 1 min using HCImage; 3) take a regular image under ambient light conditions before real-time imaging.
To ensure that the osmotic pressure of aspartate solution are equal, 12.5 and 25 mM aspartate (Sigma, St. Louis, MO, USA) were dissolved in the ACSF, 50 mM aspartate was dissolved in the low osmotic ACSF (in mM) (2.5 KCl, 115 NaCl, 10 D-glucose, 26 NaHCO3, 1.25 NaH2PO4, 1 MgCl2, 2 CaCl2; PH7.6, Osmotic pressure 270 mOsm/L). Osmotic pressure is measured by freezing point osmometer. First, 300 and 800 mOsm/L were calibrated, and then the Osmotic pressure of the solution was measured by the freezing point temperature.
2.3. Image Processing and Data Analysis
The main processes of image processing and data analysis were as follows: 1) all original images were processed with a program running at the MATLAB platform, eliminating the effect of cosmic rays from the original gray images resulted in the processed biophoton gray images; 2) the processed images were analyzed for the average gray values (AGVs) in the regions of interest (ROIs) and the background gray values (BGVs) using a same image analysis software (HCImage) as reported before [18], and the relative gray values (RGVs) were obtained as follow:
RGVs = AGVs − BGVs
2.4. Statistical Analysis
Statistical analyses were performed using Microsoft Excel and GraphPad Prism5. Paired Two-tailed t-test was used to compare the effects of biophoton emission at different concentrations of aspartate with the change of time.
3. Results
3.1. Concentration-Dependent Effects of Aspartate-Induced Biophotonic Activity in Mouse Brain Slices
A concentration-dependent increase effect was found in the biophotonic activities induced by aspartate in mouse brain slices. There were no obvious effects after long-lasting application of 12.5 mM (Figure 1(a) and Figure 1(b), green line, Table 1, n = 5), but a gradual increase of biophotonic activities was observed after the application of 25 mM aspartate and achieved the significant difference within approximately 75 min (Figure 1(a) and Figure 1(b), blue line, Table 1, n = 4). Such effects were very obvious after the application of 50 mM aspartate, presenting a significant increase within approximately 25 min (Figure 1(a) and Figure 1(b), red line, Table 1, n = 6), and then a further enhancement was found, showing a similar trend to that of 25 mM aspartate with the extension of time (Figure 1(a) and Figure 1(b)).
3.2. Characteristic Changes in Biophotonic Activities Induced by Aspartate in Mouse Brain Slice
For comparing the effect of biophoton activity induced by 50 mM aspartate with that by 50 mM glutamate in mouse brain slice as reported earlier [18], we further observed the characteristic changes of 50 mM aspartate-induced biophotonic activities under different conditions. The application of 50 mM aspartate to mouse brain slices led to a significant increase in biophotonic activities and achieved the maximal effect within approximately 69 min (69 ± 2.4 min, n = 6, Figures 2(a)-(c), the effect of initiation), which remained stable for a relatively long time although a slightly gradual increase of biophotonic activities was
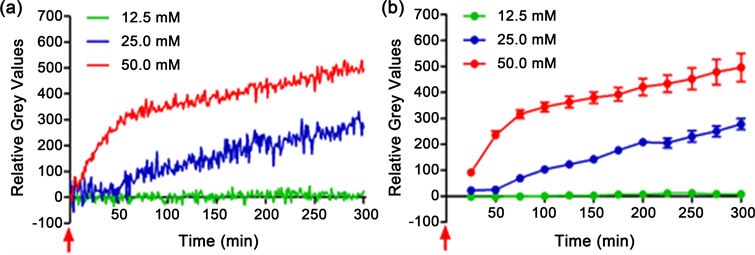
Figure 1. Aspartate induced biophotonic activities present concentration-dependent effects in mouse brain slices. (a) Each curve (green, blue or red) represents the dynamic change of biophotonic activities in a representative brain slice after the long-lasting application of 12.5 mM, 25 mM and 50 mM, respectively. (b) The sum of the time course of the average change of relative grey values of 25 continuously processed original gray images in A. Data show mean ± s.e.m. and the statistical analysis was shown in Table 1. N = the number of slices from the same number of mice (5, 4, 6 for 12.5 mM, 25 mM and 50 mM, respectively). Red arrow indicates the time point for the application of aspartate.

Table 1. Relative gray values (RGVs) of brain slices after application of different concentrations of aspartate at different time periods.
Asterisks indicate a significant difference between 25 mM and 12.5 mM, **p < 0.01; ***p < 0.001; plus signs indicate a significant difference between 50 mM and 25 mM, ++p < 0.01; +++p < 0.001.
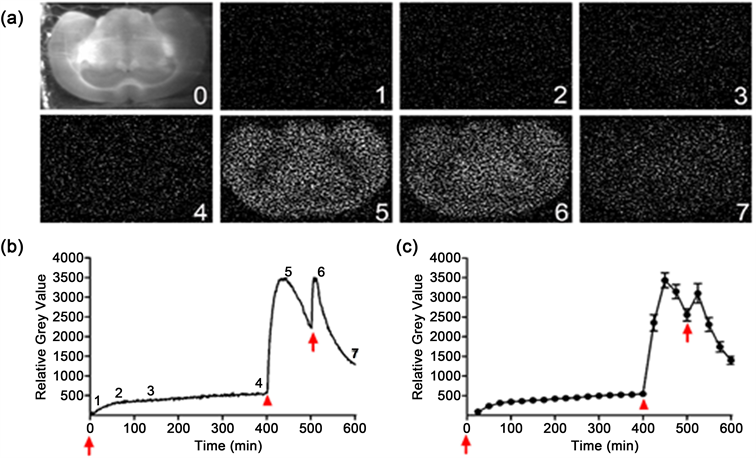
Figure 2. Imaging biophotonic activities induced by aspartate in mouse brain slices. ((a), (b)) representative biophoton gray images in a brain slice after the application of 50 mM aspartate (a) and the dynamic changes in biophotonic activities demonstrated by relative gray values (RGVs) in this slice after a lasting imaging for 600 min (b). In Figure 2(a), the images of digit 0 presents 450 μM coronal brain slice of hippocampal, the digits 1 - 7 corresponding to the same digits in B present the representative biophoton gray images at the selected time points after the application of 50 mM aspartate, 1 - 2 represent the initiation phase, 3 - 4 represent the maintenance phase, 5 represents the washing phase, and 6 - 7 represent the aspartate reapplication phase, respectively. (c) The sum of the time course of the average change of RGVs in B from 25 continuously processed original gray images (n = 6). Two arrows in B and C indicate the time points for the application of 50 mM aspartate and arrowhead for slice washing. Data are shown as the mean ± sem.
observed over time (Figure 2(b) and Figure 2(c), the effect of maintenance); a lasting slice washing with ACSF at 400 min after application of aspartate led to a further increase of biophotonic activities within approximately 4 min (4 ± 3.1 min, n = 6), which then decayed gradually (Figure 2(b) and Figure 2(c), washing effect). The second application of 50 mM aspartate after slice washing for 100 min produced a quick increase of biophotonic activities and achieved the maximal effect within approximately 10 min (10 ± 0.88 min, n = 6), but the peak value was slightly smaller than the maximum effect of washing, and then rapidly decayed (Figure 2(b) and Figure 2(c), reapplication effect).
4. Discussion
In this study, we found that aspartate can significantly induce biophotonic activity in mouse brain slices, and the intensity and effect characteristics of aspartate are consistent with those of glutamate as reported previously; both of them present the characteristic changes of initiation, maintenance, washing and reapplication [18]. However, some differences should be emphasized as follows: 1) aspartate has a faster effect than glutamate, and relatively stable effect is achieved within 60 min for aspartate, while it takes about 90 min for glutamate; 2) the intensity of biophotonic activity induced by aspartate was more obvious than that by glutamate; 3) the maximum effect of aspartate reapplication was not significantly higher than that of washing. The mechanisms of these differences are unclear and may be related to the glutamate receptors.
Glutamate receptors can be divided into two categories: one is the ionic receptors, including NMDA receptor, kainite receptor (KAR) and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR), which are coupled with ion channels to form receptor channel complexes that mediate fast signal transmission [20]. Another is the metabolic receptors (mGluRs), which are coupled with G-protein in the membrane [20]. When these metabolic receptors are activated, the relatively slow physiological effects can be produced through the signal transduction system composed of G-protein effector enzyme and the intracellular second messenger, etc. Electrophysiological studies have shown that aspartate can specifically activate NMDA receptors and induce neural action potentials [5], which is consistent with the function of glutamate. However, aspartate has no obvious effect on AMPA receptor [6], so whether the receptor differences also determines the differences in biophotonic activities induced by glutamate and aspartate is unknown and needs to be further clarified. Since the glutamate-induced biophotonic activities and transmission in neural circuits may be involved in the mechanisms of neuronal signal transmission [18], such differences in biophotonic activities induced by glutamate and aspartate may reflect the evolution of species. It has been demonstrated that the content of aspartate is more abundant in the brain of lower organisms as compared to that of glutamate [21], and therefore, glutamate gradually replaced aspartate as the most abundant neurotransmitter in the nervous system when the evolution of organisms is higher. Although whether this evolutionary significance is related to the advanced brain functions of organisms deserves further determination, however, it is understandable that lower organisms may need more rapid and efficient information processing for brain functions (fast response) in the process of adaptation to the environment, while the brain activity in higher organisms, such as human beings, may need more sustained information processing and low energy consumption (persistent response).
Acknowledgements
This work was supported by the Sci-Tech Support Plan of Hubei province (2014BEC086) and the research funds of South-Central University for Nationalities (XTZ15014 and CZP 18008).
Conflicts of Interest
The authors declare no conflicts of interest regarding the publication of this paper.
References
- 1. Falke, J.J., Bass, R.B., Butler, S.L., Chervitz, S.A. and Danielson, M.A. (1997) The Two-Component Signaling Pathway of Bacterial Chemotaxis: A Molecular View of Signal Transduction by Receptors, Kinases, and Adaptation Enzymes. Annual Review of Cell and Developmental Biology, 13, 457-512. https://doi.org/10.1146/annurev.cellbio.13.1.457
- 2. Roberts, P.J. (1974) The Release of Amino Acids with Proposed Neurotransmitter Function from the Cuneate and Gracile Nuclei of the Rat in Vivo. Brain Research, 67, 419-428. https://doi.org/10.1016/0006-8993(74)90491-0
- 3. Nadler, J.V., Vaca, K.W., White, W.F., Lynch, G.S. and Cotman, C.W. (1976) Aspartate and Glutamate as Possible Transmitters of Excitatory Hippocampal Afferents. Nature, 260, 538-540. https://doi.org/10.1038/260538a0
- 4. Collins, G.G.S. (1979) Evidence of a Neurotransmitter Role for Aspartate and Gamma-Aminobutyric Acid in the Rat Olfactory Cortex. The Journal of Physiology, 291, 51-60. https://doi.org/10.1113/jphysiol.1979.sp012799
- 5. Patneau, D.K. and Mayer, M.L. (1990) Structure-Activity Relationships for Amino Acid Transmitter Candidates Acting at N-methyl-D-aspartate and Quisqualate Receptors. Journal of Neuroscience, 10, 2385-2399. https://doi.org/10.1523/JNEUROSCI.10-07-02385.1990
- 6. Curras, M.C. and Dingledine, R. (1992) Selectivity of Amino Acid Transmitters Acting at N-methyl-D-aspartate and Amino-3-hydroxy-5-methyl-4-isoxazolepropionate Receptors. Molecular Pharmacology, 41, 520-526. https://doi.org/10.1002/med.2610120203
- 7. Bellocchio, E.E., Reimer, R.J., Fremeau, R.T.Jr. and Edwards, R.H. (2000) Uptake of Glutamate into Synaptic Vesicles by an Inorganic Phosphate Transporter. Science, 289, 957-960. https://doi.org/10.1126/science.289.5481.957
- 8. Fremeau, R.T.Jr., Troyer, M.D., Pahner, I., Nygaard, G.O., Tran, C.H., Reimer, R.J., Bellocchio, E.E., Fortin, D., Storm-Mathisen, J. and Edwards, R.H. (2001) The Expression of Vesicular Glutamate Transporters Defines Two Classes of Excitatory Synapse. Neuron, 31, 247-260. https://doi.org/10.1016/S0896-6273(01)00344-0
- 9. Miyaji, T., Echigo, N., Hiasa, M., Senoh, S., Omote, H. and Moriyama, Y. (2008) Identification of a Vesicular Aspartate Transporter. Proceedings of the National Academy of Sciences of the United States of America, 105, 11720-11724. https://doi.org/10.1073/pnas.0804015105
- 10. Zhou, M., Peterson, C.L., Lu, Y.B. and Nadler, J.V. (1995) Release of Glutamate and Aspartate from CA1 Synaptosomes: Selective Modulation of Aspartate Release by Ionotropic Glutamate Receptor Ligands. Journal of Neurochemistry, 64, 1556-1566. https://doi.org/10.1046/j.1471-4159.1995.64041556.x
- 11. Bradford, S.E. and Nadler, J.V. (2004) Aspartate Release from Rat Hippocampal Synaptosomes. Neuroscience, 128, 751-765. https://doi.org/10.1016/j.neuroscience.2004.06.065
- 12. Bakken, I.J. (1997) Lactate Formation from [U-13C] Aspartate in Cultured Astrocytes: Compartmentation of Pyruvate Metabolism. Neuroscience Letters, 237, 117-120. https://doi.org/10.1016/S0304-3940(97)00834-3
- 13. Bakken, I.J., White, L.R., Aasly, J., Unsgård, G. and Sonnewald, U. (1998) [U-13C] Aspartate Metabolism in Cultured Cortical Astrocytes and Cerebellar Granule Neurons Studied by NMR Spectroscopy. Glia, 23, 271-277. 3.0.CO;2-7>https://doi.org/10.1002/(SICI)1098-1136(199807)23:3<271::AID-GLIA9>3.0.CO;2-7
- 14. Prasad, A. and Pospísil, P. (2013) Towards the Two-Dimensional Imaging of Spontaneous Ultra-Weak Photon Emission from Microbial, Plant and Animal Cells. Scientific Reports, 3, Article No. 1211. https://doi.org/10.1038/srep01211
- 15. Bertogna, E., Bezerra, J., Conforti, E. and Gallep, C.M. (2013) Acute Stress in Seedlings Detected by Ultra-Weak Photon Emission. Journal of Photochemistry and Photobiology B: Biology, 118, 74-76. https://doi.org/10.1016/j.jphotobiol.2012.11.005
- 16. Van Wijk, E., Kobayashi, M., van Wijk, R. and van der Greef, J. (2013) Imaging of Ultra-Weak Photon Emission in a Rheumatoid Arthritis Mouse Model. PLoS ONE, 8, e84579. https://doi.org/10.1371/journal.pone.0084579
- 17. Dotta, B.T., Saroka, K.S. and Persinger, M.A. (2012) Increased Photon Emission from the Head While Imagining Light in the Dark Is Correlated with Changes in Electroencephalographic Power: Support for Bokkon’s Biophoton Hypothesis. Neuroscience Letters, 513, 151-154. https://doi.org/10.1016/j.neulet.2012.02.021
- 18. Tang, R. and Dai, J. (2014) Spatiotemporal Imaging of Glutamate-Induced Biophotonic Activities and Transmission in Neural Circuits. PLoS ONE, 9, e85643. https://doi.org/10.1371/journal.pone.0085643
- 19. Tang, R. and Dai, J. (2014) Biophoton Signal Transmission and Processing in the Brain. Journal of Photochemistry and Photobiology B, 139, 71-75. https://doi.org/10.1016/j.jphotobiol.2013.12.008
- 20. Ozawa, S. (1998) Glutamate Receptors in the Mammalian Central Nervous System. Progress in Neurobiology, 54, 581-618. https://doi.org/10.1002/jmv.20941
- 21. Trams, E.G. (1981) On the Evolution of Neurochemical Transmission. Differentiation, 19, 125-133. https://doi.org/10.1111/j.1432-0436.1981.tb01140.x