Advances in Bioscience and Biotechnology
Vol.2 No.5(2011), Article ID:7828,10 pages DOI:10.4236/abb.2011.25047
The effect of maternal thyroxine injection on growth, survival and development of the digestive system of Nile tilapia, Oreochromis niloticus, larvae
![]()
1Fish Reproduction Laboratory, National Institute of Oceanography and Fisheries, Alexandria, Egypt;
2Zoology Department, Faculty of Science, Al-Azhar University, Cairo, Egypt.
E-mail: *mostafa_mousa2002@yahoo.com
Received 19 July 2011; revised 8 September 2011; accepted 22 September 2011.
Keywords: Fish Larvae; O. niloticus; Thyroxine; Growth; Digestive System
ABSTRACT
The present study deals with the effect of exogenous treatment of O. niloticus females with L-thyroxine (T4) on the development of the digestive system during larval rearing, and its subsequent effect on larval growth and survival. The development of the digestive tract and accessory glands was investigated histologically and histochemically in the developing O. niloticus larvae, from control and T4-treated spawners. During yolk-sac absorption, the digestive system of the fish underwent further differentiation and the rudimentary alimentary canal became segmented into four different histological regions: the buccopharynx, oesophagus, stomach and intestine. The injection of females O. niloticus with thyroxine (1 or 10 μg·T4/g·BW) greatly enhanced the development of the digestive tract and accessory glands of larvae as indicated by the quantitative and qualitative changes of the mucus composition from predominantly neutral to a mixture of neutral and acid mucosubstances, or acid mucosubstances occurred during the rearing period for the larvae produced from T4-treated females. This may be due to the direct effect of exogenous thyroxine, which might have been transferred from maternal circulation into the oocytes and larvae, on the synthesis of proteins, which increased with larval development. Thus, thyroxine directly or indirectly improved O. niloticus larval growth, since a marked increase in both, length and weight of larvae occurred during the experimental period. In addition, larvae from treated females also gave a significantly higher survival rate than that of control. It could be concluded that exogenous T4 in maternal circulation might have been transferred into oocytes and larvae. The transferred thyroid hormone appears to play some role in the early development of larvae and may confer a distinct advantage for the growth of the offspring of the Nile tilapia, O. niloticus.
1. INTRODUCTION
For the successful propagation of any species, larval rearing is considered a major bottleneck [1,2]. During this period, transition from endogenous to exogenous feeding and weaning to artificial diets is characterized by high mortality rates, which are often linked to an inadequate food composition [3]. The underlying reason is that, the digestive tract at this time is largely undifferentiated, with low digestive enzyme activity, and its short length results in rapid evacuation of ingested food [4]. Knowledge of differentiation of the gut during ontogenetic development, associated with food digestion and assimilation processes, is essential for understanding the nutritional physiology of larval fishes, and synchronizing the physiological stage of development with feeding practices and rearing protocols. A detailed understanding of the development of the gut may help to reveal limiting factors in rearing protocols, reducing bottlenecks in larvae culture and weaning processes [5,6].
Attempts to rear the Nile tilapia, Oreochromis niloticus in the hatcheries in Egypt have yielded inconsistent and unreliable production of fry. Larval rearing has proven extremely difficult with heavy larval mortalities usually occurring during the second and third weeks from hatching. The cause of these mortalities may be physiological in nature given that starvation, due to the physical inability to feed after exhaustion of endogenous reserves. The period of mixed nutrition in rearing larvae of Nile tilapia, Oreochromis niloticus is considered as one of the most critical events during larval early life stages, since a several-day delay in feed availability influences posterior larval growth and survival. In Egyptian hatcheries, the transition from endogenous to exogenous nutrition often results in high mortalities of Nile tilapia larvae, which might be attributed to nutritional and infection problems. Several authors have implicated bacteria with problems of larval rearing in different finfish species [7,8].
Thyroid hormone (TH) plays an indispensable role during embryonic and larval periods of fish development [9,10]. Cognizant of the potentiating effect of thyroxine (T4) on fish larval growth and survival, it was worth investigating if exogenous T4 application in female brood fish could translate to better growth and survival of larvae. The action of thyroid hormones is mediated by their binding to nuclear receptors that act directly on target genes, binding to specific DNA sequences to increase or decrease expression of the gene and bring about the cellular response to thyroid hormones [11].
Histological and histochemical description of the development of the digestive system may provide important information for the establishment of physiologically sound rearing methods to improve commercial larval rearing [12]. Several studies have described the development of digestive tract organs in diverse fish species, including California halibut [8]; Japanese eel larvae [13]; large yellow croaker [6]; shi drum [14]; flatfish brill [15] and starlet [16]. However, there is no available information on the differentiation of the gastrointestinal tract of Nile tilapia during early life stages. Therefore, the objective of the present work was to describe the effect of thyroxine on the development of the gut and accessory glands in larvae of Nile tilapia, Oreochromis niloticus, to achieve a better understanding of their organization and functionality, and to provide a basis for future nutritional studies during larval rearing.
2. MATERIAL AND METHODS
2.1. Study Site
The present study was carried out at both El-Serw Fish Research Farm and El-Matareyya Research Station between 1 January and 30 July 2010.
2.2. Treatments and Spawning
Before spawning, brood fish were kept in two ponds, males and females were separated since January in two different ponds, they were fed daily with 40% protein diet to ensure a good quality and quantity of eggs. Medium-size tilapia brood fish (150 - 250 g) were used. Semi-natural spawning was carried in spawning hapas at the first of May (temperature: 23˚C - 25˚C). Brood O. niloticus were stocked into 30 fine-mesh 1-m2 spawning hapas at a rate of 2 males and 4 females per hapa. Females were randomly divided into three groups representing three treatments with ten replicates each. Fish in treatments 1 and 2 were injected once with thyroxin (T4) (Sigma Chemicals co., St. Louis, MO), dissolved in dimethylsulfoxide (DMSO), at 1 and 10 μg·T4/g·BW of fish, respectively. Fish in treatment 3 were injected only with DMSO and served as controls. After injection, females were stocked with hormonally-untreated running males and allowed to spawn. Breeding activity was monitored daily. Once breeding occurred, the other fish were removed and the brooding female left to incubate the progeny.
Immediately after hatching the larvae, obtained from each of the previous groups; control and thyroxine treated groups, were classified into three groups (three aquaria for each group). The density of larvae for each aquarium is about 500 larvae/aquarium i.e. 10 larvae/l. Since the sensitivity of fish to temperature is dependent on developmental stage (age), fry from a single brood were used in order to control for the time elapsed after fertilization (and therefore, developmental stage). The aquaria were maintained at 25˚C and ambient photoperiod. Water quality in the aquaria was maintained by partial water exchange (70%) daily. Gentle aeration was conducted with pressurized air. The larvae received natural feed of fresh plankton obtained from fertilized pond with plankton net.
2.3. Larvae Sampling and Processing
Both standard length (SL) and weight (W) of larvae at hatching, 2, 4, 7, 14, 21, 28 and 35 days old, were measured for each group. Twenty larvae were randomly sampled, anaesthetized, placed on paper towels for about 10 seconds to remove most of the adhering water, and individually both, measured and weighed. At the end of the experiment the surviving number of larvae for each treatment was determined.
For histological and histochemical study, the larvae were anaesthetized in a solution (40 mg/l) of clove oil (Sigma) and fixed in toto in Bouin’s fluid at room temperature for 48 h. Then, the samples were transferred to 70% alcohol after fixation and dehydrated through series of graded ethanol, cleared in xylene, embedded in paraplast (M.P. 56˚C - 58˚C) and serial transverse and longitudinal sections, 5 μm thick, were cut and mounted on glass slides. For each specimen, selected sections were stained with the following techniques.
1) Harris’ alum hematoxylin and aqueous solution of eosin (1%) was used as a counter stain (H & E) [17].
2) Alcian blue (AB)/periodic acid-Schiff reagent (PAS) pH 1.0 [18], counter stained with orange G. Positive reactions for mucous cell histochemistry at AB/PAS-OG pH 1.0 were blue or blue greenish for acid mucins, red for neutral mucins and reddish purple for a combination of neutral and acid mucins.
2.4. Statistical Analysis
Results were analyzed with the SPSS (Statistical Package for Social Sciences) statistical package. Paired-samples “t” test was applied to compare means. The level for accepted statistical significance was P £ 0.05.
3. RESULTS
3.1. The Effect of Thyroxine Treatment on Growth and Survival Rate
3.1.1. Growth in Length
The results in Table 1 showed that the injection of O. niloticus females with thyroxine (1 or 10 μg·T4/g·BW) improved larval growth. Moreover, treatment with the high dose of thyroxine (10 μg·T4/g·BW) gave significantly higher increase in length of larvae than low dose (1 μg·T4/g·BW) as shown in Table 1.
3.1.2. Growth in Weight
The larvae of T4-treated females exhibited significantly greater average body weight than those of control as represented in Table 2. Moreover, treatment with the high dose of thyroxine (10 μg·T4/g·BW) gave significantly higher values of larval body weight than low dose of thyroxine (1 μg·T4/g·BW) as represented in Table 2.
3.1.3. Survival Rate
The data represented in Table 3 showed that the injection of O. niloticus females with thyroxine (1 or 10 μg·T4/g·BW) improved larval rearing, since low mortalities occurred over the rearing period of 35 days. The larvae of T4-treated females exhibited significantly higher survival rate than control.
Moreover, the injection of females with high dose of thyroxine resulted in a significantly higher survival rate than low dose. The average values of survival rate were 91.24% ± 1.06%, 76.55% ± 1.32% and 40.75% ± 1.09% for larvae of T4-treated females with high dose of thyroxine (10 μg·T4/g·BW), low dose of thyroxine (1 μg·T4/g·BW), and control larvae, respectively as represented in Table 3.
3.2. The Effect of Thyroxine Treatment on the Histology and Histochemistry of the Digestive System
The development of the digestive system of Oreochromis niloticus was follow up during the development of larvae, obtained from control and thyroxine injected females, by both histological and histochemical techniques. During yolk-sac absorption, the digestive system of the fish underwent further differentiation and the rudimentary alimentary canal became segmented into four different histological regions: the buccopharynx, oesophagus, sto
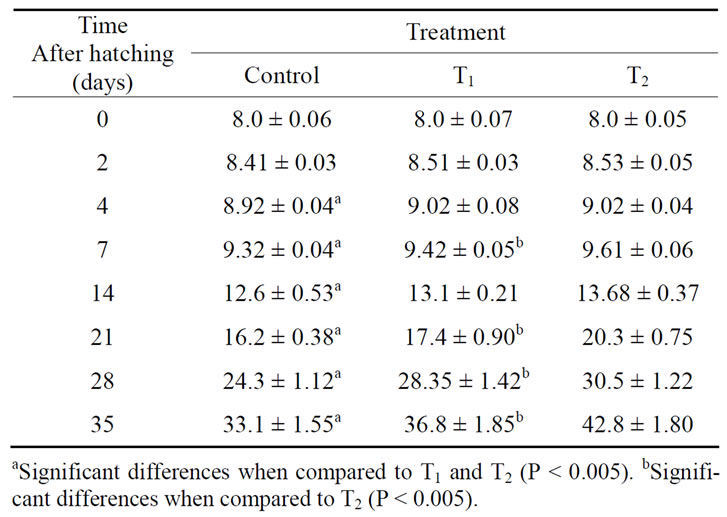
Table 1. Total length (mean ± SD mm), at different ages, of O. niloticus larvae from T4-treated and control spawners.
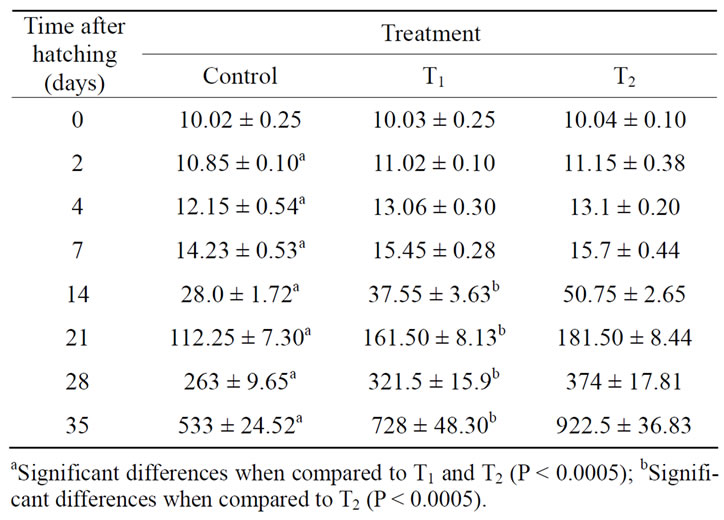
Table 2. Total weight (mean ± SD mg), at different ages, of O. niloticus larvae from T4-treated and control spawners.
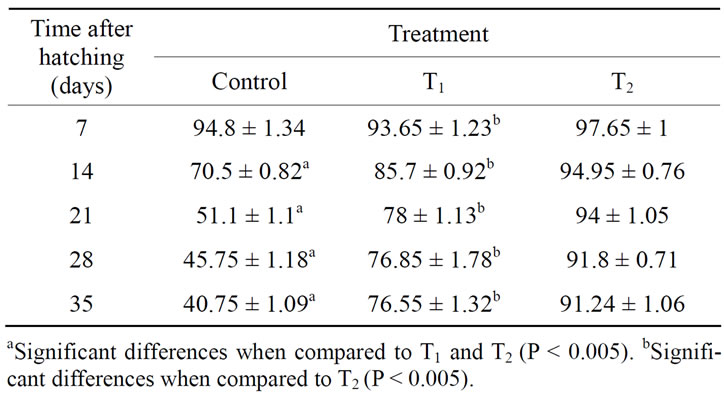
Table 3. Survival rate (%), after 7, 14, 21, 28 and 35, of O. niloticus larvae from T4-treated and control spawners.
mach and intestine. Furthermore, the histological and histochemical changes in the accessory glands were investigated.
3.2.1. Buccopharynx
At 4 days post hatching (dph), the mean total length of the larvae obtained from control females was 8.41 ± 0.03 mm. The buccopharyngeal epithelium is composed of a few numbers of squamous cells covered differentiated taste buds; scattered in the anterior and posterior region of the buccal cavity. Mucus-secreting cells are arranged in one layer in the buccopharyngeal epithelium. Their contents attain red colour with Alcian-PAS reaction, indicating the presence of neutral muco-polysaccharides (NMPS) (Figure 1(a)). The chondrocytes of the rudimentary gill arches are visible under the epithelium of the posterior buccopharynx. The larvae obtained from females treated with the high dose of thyroxin (10 μg·T4/ g·BW) had a mean total length of 8.53 ± 0.05 mm after 4 dph. In these larvae, the buccopharyngeal epithelium is thicker than in control larvae. In addition, mucussecreting cells and taste buds well developed (Figure 1(b)).
The larvae obtained from control females had a mean total length of about 33.1 ± 1.55 mm after 35 dph. The buccopharyngeal wall consists mainly of mucosa and submucosa. The stratified epithelium consists of several cell-layers. Mucus-secreting cells are numerous and constitute an important feature in the buccal epithelium. Their contents attain red colour with Alcian-PAS reaction, indicating the presence of NMPS. Taste buds are much concentrated in the anterior portion of the buccopharynx than in the posterior one. The submucosa is formed of an outer layer of fibrous connective tissue, the stratum compactum, and an inner layer of loose areolar connective tissue, richly supplied with blood capillaries. Muscle fibers, mostly of striated type, are occasionally encountered in the submucosa (Figures 1(c) and (e)). At 35 dph, the mean total length of the larvae obtained from females treated with the high dose of thyroxine (10 μg·T4/g·BW) had a mean total length of 42.8 ± 1.8 mm. The buccopharyngeal epithelium is thicker than in control larvae. The taste buds are larger in size and well developed. Mucus-secreting cells were numerous and well developed (Figures 1(d) and (f)).
3.2.2. Oesophagus
At 4 dph, the digestive tube of the larvae obtained from control female started to differentiate in the terminal region of the oesophagus. The epithelial mucosa is made up of 2 - 4 layers of large saccular mucus cells (Figures 1(g) and (i)). They are intermingled with much fewer of squamous epithelial cells and contain mixed of strong acid and weakly N MPS in the anterior part of the oesophagus and strong NMPS in the posterior one (Figures 1(g) and (i) and 2(a), (c) and (e)). The submucosa formed of a loose connective tissue, a wide circular tunica muscularis and a thin serosa (Figures 1(g) and (i) and 2(a), (c) and (e)). At 4 dph, the oesophagus mucus cells of the larvae obtained from treated female are large in size and contain mixed of strong acid and neutral mucopoly-sac-
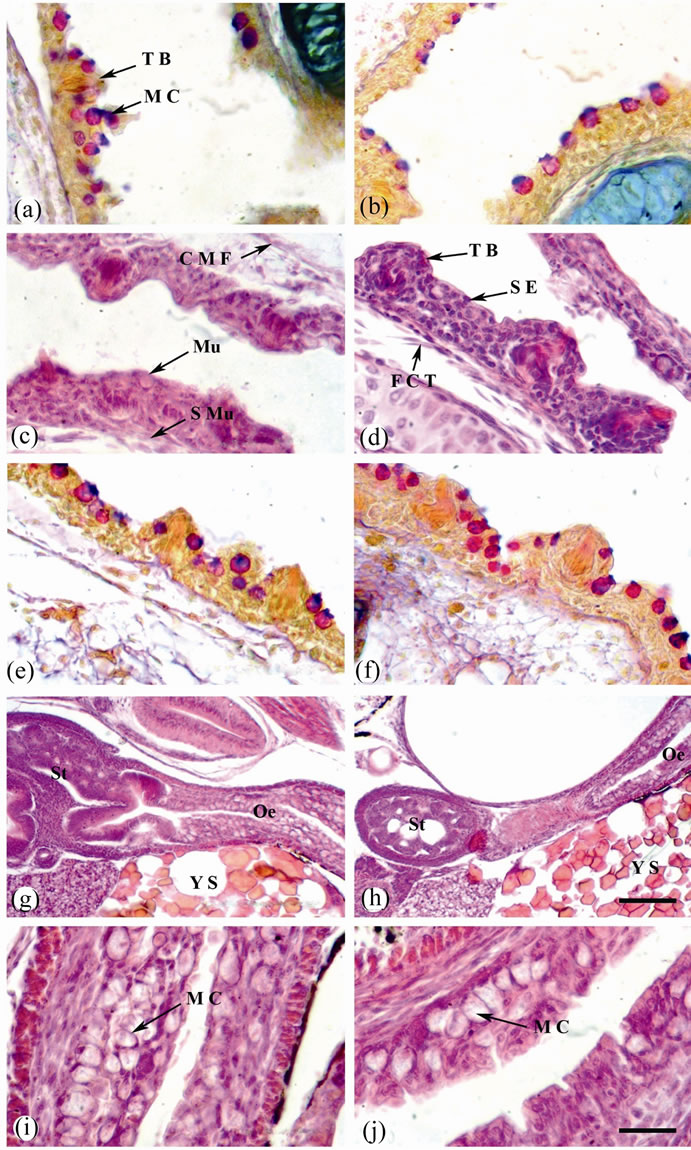
Figure 1. Sagittal sections of O. niloticus larvae stained with AB/PAS-OG (a), (b), (e) and (f) or H & E (c), (d) & (g-j). (a) Buccopharynx of 4 dph. larvae from control female. (b) Buccopharynx of 4 dph. larvae from treated female (with the high dose of thyroxine; 10 μg·T4/g·BW). (c) and (e) Buccopharynx of 35 dph. larvae from control female. (d) and (f) Buccopharynx of 35 dph. larvae from treated female. (g) Oesophagus and stomach of 4 dph. larvae from control female. (h) Oesophagus and stomach of 4 dph. larvae from treated female. (i) A magnified portion of (g) showing oesophagus. (j) A magnified portion of (h) showing oesophagus. Taste bud (T B); mucus cell (M C); mucosa (Mu); sub mucosa (S Mu); circular muscle fiber (C M F); stratified epithelium (S E); fibrous connective tissue (F C T); oesophagus (Oe); stomach (St) and yolk sac (Y S). Scale bar = 25 µm (a)-(f), (i) and (j) and 100 µm (g) and (h).
charides in the anterior part and weakly acid and neutral mucopoly-saccharides in the posterior one (Figures 1(h) and (j) and 2(b), (e) and (f)).
After 35 dph, in the oesophagus of the larvae obtained from control female the mucosal folds are formed. Mucus cells are highly abundant. Their contents attain mixed of strong blue and red colors with Alcian-PAS reaction, indicating the presence of mixture of strong acid and weakly NMPS in the anterior part of the oesophagus and weakly NMPS in the posterior one. The muscular coat is made up of one layer, only the circular. They are of striated type and arranged in small bundles held together by an areolar connective tissue. The serosa consists of a thin layer of pavement epithelium (Figures 2(g), (i) and (k)). In the oesophagus of the larvae obtained from treated female after 35 dph, the mucosa is greatly folded.
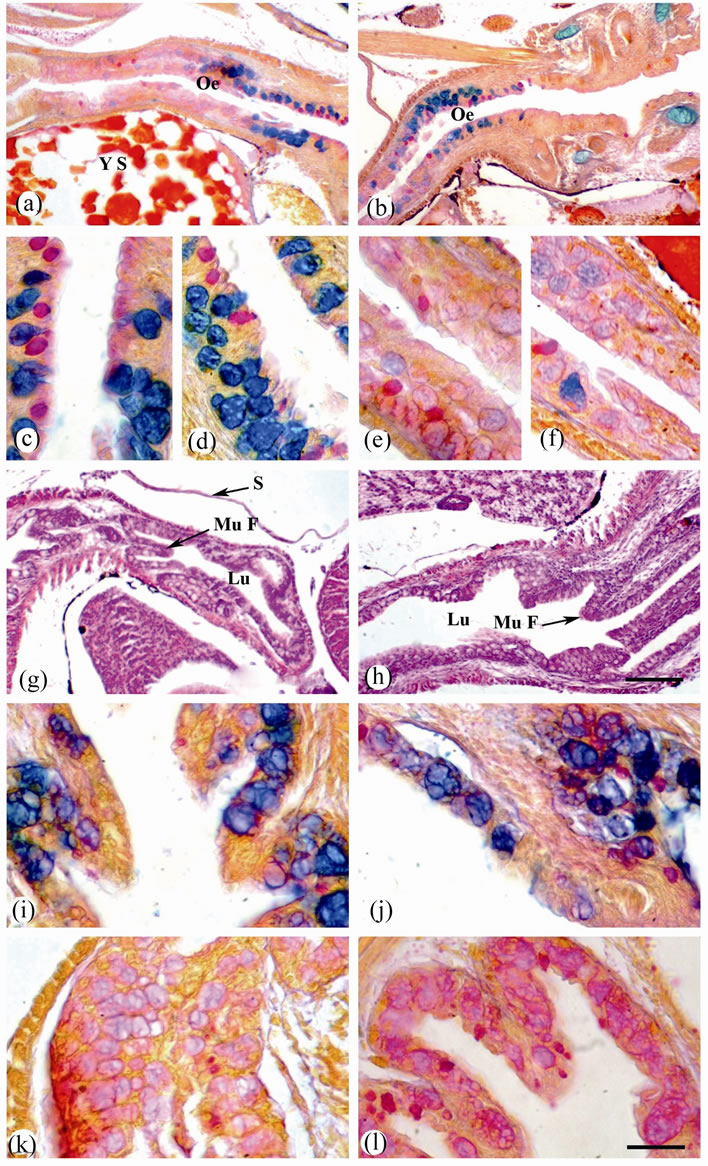
Figure 2. Sagittal sections of O. niloticus larvae stained with AB/PAS-OG (a-f) and (i-l) or H & E (g) and (h). (a) Oesophagus of 4 dph. larvae from control female. (b) Oesophagus of 4 dph. larvae from treated female (with the high dose of thyroxine; 10 μg·T4/g·BW). (c) Anterior oesophagus of 4 dph. larvae from control female. (d) Anterior oesophagus of 4 dph. larvae from treated female. (e) Posterior oesophagus of 4 dph. larvae from control female. (f) Posterior oesophagus of 4 dph. larvae from treated female. (g) Oesophagus of 35 dph. larvae from control female. (h) Oesophagus of 35 dph. larvae from treated female. (i) Anterior oesophagus of 35 dph. larvae from control female. (j) Anterior oesophagus of 35 dph. larvae from treated female. (k) Posterior oesophagus of 35 dph. larvae from control female. (l) Posterior oesophagus of 35 dph. larvae from treated female. Oesophagus (Oe); mucosal fold (Mu F); lumen (Lu) and serosa (S). Scale bar = 100 µm (a), (b), (g) and (h) and 25 µm (c)-(f) & (i)-(l).
The lumen is very wide (Figure 2(h)). Mucus cells are highly abundant; they tend to be more concentrated on the sides and the bases of the mucosal folds. Mucus cells show a variety of forms, being saccular, spherical or pyriform in shape. They are increase in size towards the posterior part of the oesophagus. Their contents attain black blue and red colors with Alcian-PAS reaction, indicating the presence of mixture of carboxylated and sulphated glycoproteins (Figures 2(h), (j) and (l)).
3.2.3. Stomach
After 4 dph, the stomach of the larvae obtained from control female was started to develop; its epithelial mucosa consists of simple columnar epithelium and develop into few gastric glands. Alcian-PAS reaction, however, gave a negative staining reaction. The submucosa is made up of areolar connective tissue, richly supplied with lymph spaces and blood vessels. Tunica muscularis consists of two layers: outer longitudinal and inner circular muscle fibres. The serosa is made up of simple squamous epithelium (Figures 3(a) and (c)). Also, in the stomach of the larvae obtained from treated female after 4 dph, the epithelial mucosa consisted of simple columnar epithelium and developed into gastric glands. AlcianPAS staining gave a negative reaction. However, muscular coat is well developed (Figures 3(b) and (d)).
In the larvae obtained from control female after 35 dph, the mucosa of the stomach was organized in folds. The epithelial mucosa consists of a thin layer of simple columnar epithelium towards the lumen and a thick layer of gastric glands towards the base. Gastric glands attain red colour with PAS reaction, indicating the presence of NMPS (Figures 3(e), (g) and (i)). In the larvae obtained from treated female after 35 dph, the stomach had highly abundant gastric glands which contained NMPS (Figures 3(f), (h) and (j)).
3.2.4. Intestine
At 4 dph, the intestinal wall of the larvae obtained from control female is relatively thick. Mucosal folds are very short and the lumen is narrow. Mucous cells are abundant and contain strong NMPS (Figures 4(a) and (c)). In the larvae obtained from treated female after 4 dph, the lumen is a wide. Mucous cells are highly abundant and contain NMPS. Muscular coat is well developed (Figures 4(b) and (d)).
After 35 dph, the intestine wall of the larvae obtained from control female is very thin and the lumen is very wide. Mucosal folds are short, numerous and closely set with blunt or round tips. The epithelium is made up of columnar cells interspersed among the mucous ones. The latter are fewer in number and contain NMPS. The submucosa consists of loose areolar connective tissue, richly supplied with blood vessels. The circular layer of muscle
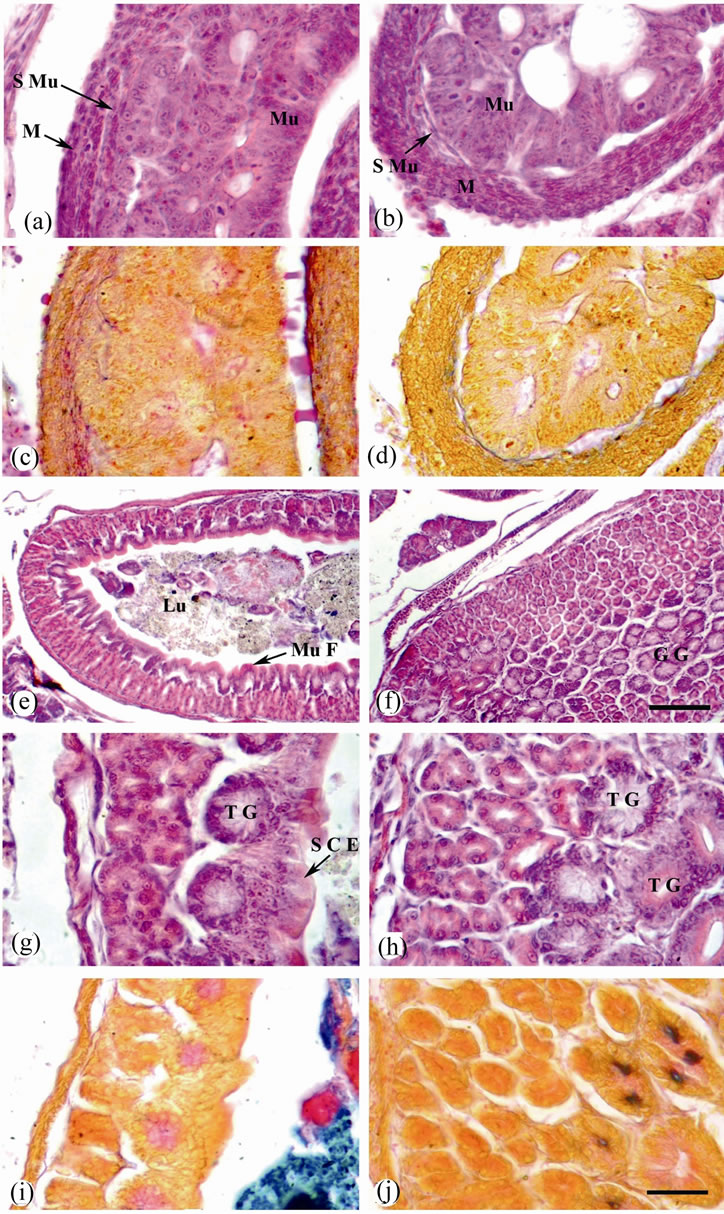
Figure 3. Sagittal sections of O. niloticus larvae stained with H & E (a), (b) and (e)-(h) or AB/PAS-OG (c), (d), (i) and (j). (a) and (c) Stomach of 4 dph. larvae from control female. (b) & (d) Stomach of 4 dph. larvae from treated female (with the high dose of thyroxine; 10 μg T4/g BW). (e) Stomach of 35 dph. larvae from control female. (f) Stomach of 35 dph. larvae from treated female. (g) and (i) A magnified portions of (e). (h) and (j) A magnified portions of (f). Mucosa (Mu); sub mucosa (S Mu); muscularis (M); mucosal fold (Mu F); lumen (Lu); gastric gland (G G); tubular gland (T G) and simple columnar epithelium (S C E). Scale bar = 25 µm (a)-(d) and (g)-(j) and 100 µm (e) and (f).
fibers is relatively thicker than the longitudinal one (Figures 4(e) and (i)). In the larvae obtained from treated female after 35 dph, the intestinal wall is thicker than that of control. Mucosal folds are long and numerous. Mucous cells are highly abundant with increase in size and contain NMPS (Figures 4(f) and (k)).
3.2.5. Accessory Glands (Liver & Pancreas)
After 4 dph, in the larvae obtained from control female, the liver appeared as a compact tissue of basophilic
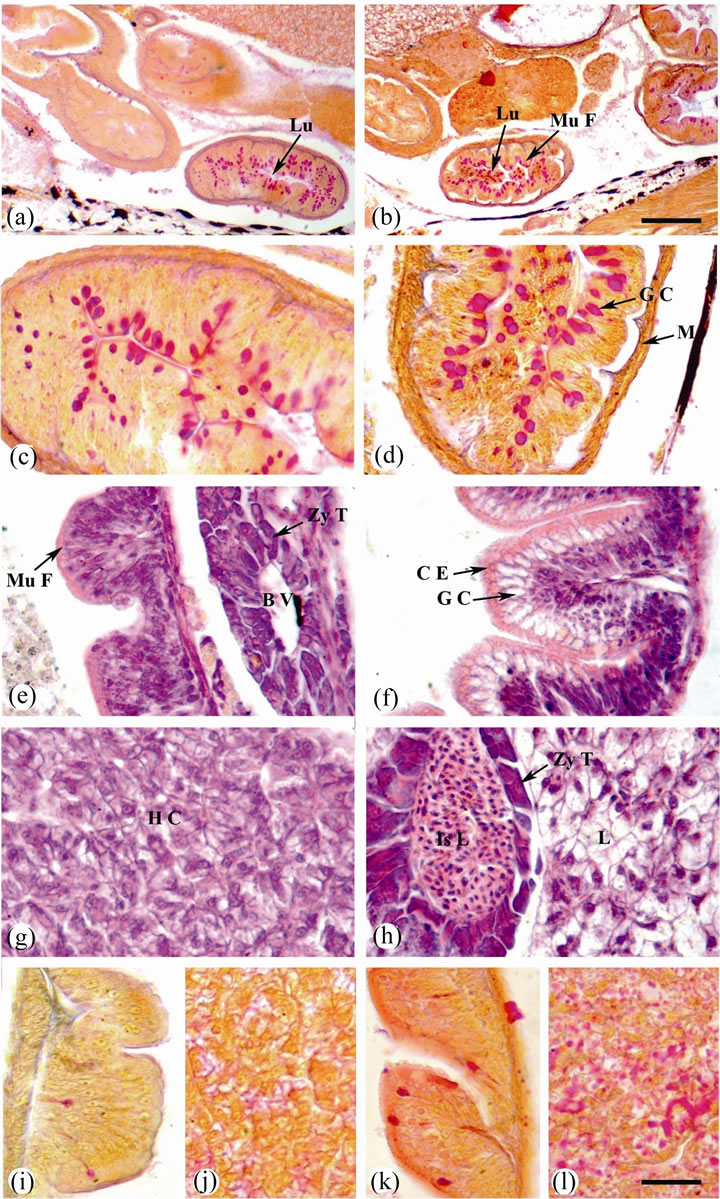
Figure 4. Sagittal sections of O. niloticus larvae stained with AB/PAS-OG (a)-(d) and (i)-(l) or H & E (e)-(h). (a) Intestine of 4 dph. larvae from control female. (b) Intestine of 4 dph. larvae from treated female. (c) A magnified portion of (a). (d) A magnified portion of (b). (e) and (i) Intestine of 35 dph. larvae from control female. (f) and (k) Intestine of 35 dph. larvae from treated female. (g) and (j) Liver of 35 dph. larvae from control female. (h) and (l) Liver of 35 dph. larvae from treated female. Lumen (Lu); mucosal fold (Mu F); goblet cell (G C); muscularis (M); blood vessel (B V); zymogenous tissue (Zy T); columnar epithelium (C E); hepatic cells (H C); Islet of Langerhans (Is L) and lipid (L). Scale bar = 100 µm (a) and (b) and 25 µm (c)-(l).
hepatocytes arranged around hepatic sinusoids beside the yolk sac. Hepatocytes had centrally located nuclei, reduced cytoplasm and lipid vacuoles (Figure 1(g)). In the larvae obtained from treated female after 4 dph, the lipid storage was observed in the liver coinciding with the onset of feeding (Figure 1(h)).
In the larvae obtained from control female at 35 dph, the liver tissue appears as a continuous mass of hepatocytes arranged in cords like around the veins and separated by narrow blood sinusoids. The hepatic cells are polygonal in shape with irregular ill-defined boundaries. Each hepatic cell has spherical large nucleus with mostly one distinct nucleolus and well defined nuclear membrane. The cytoplasm of the hepatocytes has a granular appearance and it is highly stainable with haematoxylin and eosin. It contains small vacuoles of different sizes. They attain faint red colour with PAS reaction, indicating the presence of glycogen (Figures 4(g) and (j)). The pancreas was extra-hepatic and the exocrine and endocrine regions were visible. Several clusters of basophilic pancreatic cells (exocrine pancreas) containing acidophilic zymogen granules were detected. The exocrine pancreatic cells were arranged in acini and pancreatic ducts were clearly distinguishable. The endocrine pancreas differentiated and endocrine cells arranged around many capillaries were grouped in islets of Langerhans (Figure 4(e)). In the larvae obtained from treated female after 35 dph, the hepatocytes are polygonal showed round nuclei with distinct nuclear membrane surrounded by small volume of cytoplasm. Lipid storage was observed in the liver. Hepatocytes are positive staining with PAS reaction indicating the presence of large amount of glycogen (Figures 4(h) and (l)). In the larvae obtained from treated female after 35 dph, pancreatic islets of Langerhans are surround a blood vessel and composed of a mass of polygonal, rounded, and nearly oval cells. Each cell is large with darkly stainable nucleus containing one nucleolus (Figure 4(h)).
4. Discussion
The present study examined the effects of exogenous treatment of O. niloticus females with L-thyroxine (T4) on the development of the digestive system during larval development, and its subsequent effect on larval growth and survival. The treatment with L-thyroxine (T4; 1 or 10 μg T4/g BW) accelerated larval growth, since a higher increase in both, length and weight of larvae occurred during the experimental period. This may be due to the direct effect of exogenous thyroxine, which might have been transferred from maternal circulation into oocytes and larvae, on the synthesis of activin bA [9]. Treated larvae gave also a significantly higher survival rate than that of control. Similar reports showed that thyroid hormones improve survival rates and accelerate growth and development in teleost larvae [9,10,19-23]. Improvement of larval development obtained by thyroid hormone treated female is dose-dependent. Larvae obtained from treated female with high dose of thyroxine (10 μg·T4/ g·BW) showed significantly higher increase in length, weight and survival rate than those of low dose of thyroxine (1 μg·T4/g·BW). Similar dose-dependent manner to thyroid hormone treatment was observed in milkfish Chanos chanos [24], Indian major carp Cirrhina mrigala [19], grouper Epinephelus coioides [25] and common carp Cyprinus carpio [22].
In the present study, no abnormalities were observed in larvae obtained from treated female with L-thyroxine (T4; 1 or 10 μg·T4/g·BW). The data suggest that a dose of 1 μg·T4/g·BW is appropriate for accelerating growth and improving survival during the experimental period. Similar observations were obtained in grouper Epinephelus coioides [25] and common carp Cyprinus carpio [22]. In contrast, [26] reported that treatment of striped bass Morone saxatilis larvae with 100 and 50 ng/ml resulted in retarded growth and lower survival compared with fish treated with 25 ng/ml of the hormone or with control. Growth and survival of sea bass, Lates calcarifer, larvae were also depressed after treatment with relatively low doses of thyroid hormones [27]. In addition, [28] found that, while T4 accelerated yolk sac absorption in tilapia Sarotherodon niloticus, high doses of the hormone did not improve growth and survival but instead caused abnormal shapes in the pectoral fins as well as lordosis and scoliosis.
The first few weeks of development become a critical period for fish larvae, particularly when the yolk-sac is reabsorbed and exogenous feeding starts. Larvae of many fish species show three phases of feeding: endogenous, mixed, and exclusively exogenous [29]. In the present study, during the endogenous period, the larvae treated with thyroxin start to exogenous feeding before their digestive tracts are fully developed. Most teleost fishes start exogenous feeding before their digestive tracts are fully developed [3,29].
In the present study, taste buds and mucus cells in the bucco-pharyngeal cavity of the fish treated with thyroxin are abundant. The abundance of taste buds in the buccopharyngeal cavity of the fish is rather to be correlated with the way in which the fish scores its food rather than with the nature of its diet. On the other hand, the concentration of mucus cells in bucco-pharynx shows that, this part is primarily concerned with mucus production, to facilitate swallowing of food and to entrap small food particles [30].
In the present study, the number of mucous cells on the fish treated with thyroxin are found in the anterior oesophagus and increased with larval development. The components of mucous cells in the anterior oesophagus were mainly sulphated mucosubstances, to protect the mucosa against chemicals, parasites, hypertonic media and acidity, as well as to provide lubrication for ingested food particles [14]. The presence of mucous cells in the esophagus on day 3 dph, previously reported for S. Senegalensis [31] and S. solea [32] and also the strong muscular layer, coincides with the establishment of exogenous feeding. In other species these mucous cells appear later during ontogeny [3]. Also, described acidic mucosubstances for S. solea. Increased epithelial stratification in correspondence with the greater number of goblet cells and the acid glycoproteins content have been related with a supportive function for the esophageal mucosa [33]. In the present study, mucus cells are abundant in the posterior oesophagus and contain neutral MPS. Mucus secreted in the digestive tract is the first protective barrier against mechanical, physical and chemical factors as well as against infections [34]. Besides the protection of mucosa against acidic secretory products [35]. Numerous PAS positive goblet cells appeared along the digestive tract. These cells play an important role in pregastric digestion in larvae [33].
The development of gastric glands in the stomach is a prerequisite for extracellular digestion, as they secrete HCL and digestive enzymes, mainly pepsin. The change from the intracellular digestion described above, to the extracellular luminal digestion is a prerequisite for complex digestive processes such as protein digestion, in order to assimilate more food to cover the increasing energy demand of the growing larvae. In the present study, the presumptive stomach of Nile tilapia, Oreochromis niloticus larvae could already be distinguished around 4 dph. The first gastric glands in the stomach appeared at thyroxin treatment is 2 days earlier than reported in a previous study [36]. In most examined teleost larvae, a completely differentiated stomach appears several weeks after the start of exogenous feeding [37], but in species such as the common sole [38], summer flounder [39], gilthead seabream [3] and yellowtail flounder [33] a rapid development from the undifferentiated, non-secretory gut at hatching to a morphologically differentiated state was observed by 12 dph. Also, thyroxine treatment accelerates stomach organogenesis in early metamorphosis of summer flounder Paralychthys dentatus, and inhibition of T4 synthesis with thiourea (Tu) delays stomach development [40].
In the present study, mucus cells were observed in the intestine after 1 dph in larvae treated with thyroxine and their number increased with larval development. The mucus cells contain neutral muccosubstances. The presence of neutral mucins in the digestive tract has been related to the absorption of easily digested substances, such as disaccharides and short chain fatty acids [41]. In most teleosts, the anterior intestine is involved in the absorption of lipids and it is characterized by lipid inclusions in the epithelial cells of the mucosa, which are considered as a temporary storage site [42]. In the present study, no lipid inclusions were observed in the anterior segment of the intestinal mucosa of the fish larvae, although, lipid vacuoles increased in the liver throughout development. These results are similar to those already reported in Hipoglossus hipoglossus [43] and P. californicus [8]. These authors suggested that the lack of such lipid vacuoles in the intestine of Artemia sp.-fed larvae could be interpreted as a sign of reduced intestinal lipid digestion due to the rapid passage of Artemia sp. through the alimentary canal of larvae [43], whereas [8] hypothesized that the lipid content of feed did not exceed the fatty acid absorption and exporting capacities of enterocytes, resulting in no accumulation of lipids in the intestinal mucosa.
The liver and pancreas in the larvae of Nile tilapia, Oreochromis niloticus differentiate early in development. Others have reported that, the pancreas and liver differentiate and function early in the development of several teleost species [44]. The first signs of nutrient absorption were observed 2 days after exogenous feeding started. The liver showed a PAS positive reaction as a sign of glycogen storage, as observed by [45] in several species. [32] considered the glycogen storage in the liver as a sign of the onset of hepatocyte functionality, which remains throughout the larval and juvenile stages. It has been demonstrated that, the digestive tract and digestion process undergoes major developmental changes during the first weeks of life fish. Pancreas secretion function constitutes the first step of maturation process of digestive function and the second is the onset of brush border membrane enzymes in the intestine [46].
In conclusion, the ontogeny of the digestive system in the larvae of Nile tilapia, Oreochromis niloticus are good pattern in the larvae treated with thyroxine. However, species specific differences in the appearance of functional goblet cells and gastric glands were noticed. Future work must be focused on the ontogeny of enzymatic secretions to provide precise information about the functionality of the digestive tract and to evaluate the effect of different feeding and weaning strategies on digestive tract maturation.
REFERENCES
- Calzada, A., Medina, A. and González de Canales, M.L. (1998) Fine structure of the intestine development in cultured sea bream larvae. Journal of Fish Biology, 53, 340-365. doi:10.1111/j.1095-8649.1998.tb00985.x
- Cataldi, E., Albao, C., Boglione, A., et al. (2002) Acipenser naccarii: Fine structure of the alimentary canal with references to its ontogenesis. Journal of Applied Ichthyology, 18, 329-337. doi:10.1046/j.1439-0426.2002.00383.x
- Sarasquete, M.C., Polo, A. and Yúfera, M. (1995) Histology and histochemistry of the development of the digestive system of larval gilthead seabream. Sparus aurata L. Aquaculture, 130, 79-92. doi:10.1016/0044-8486(94)00175-N
- Govoni, J.J., Boehlert, G.W. and Watanabe, Y. (1986) The physiology of digestion in fish larvae. Environmental Biology of Fishes, 16, 59-77. doi:10.1007/BF00005160
- Hamlin, H., Von Herbing, I.H. and Kling, L. (2000) Histological and morphological evaluations of the digestive tract and associated organs of haddock through-out post-hatching ontogeny. Journal of Fish Biology, 57, 716-732. doi:10.1111/j.1095-8649.2000.tb00270.x
- Mai, K., Yu, H., Ma, H., et al. (2005) A histological study on the development of the digestive system of Pseudosciaena crocea larvae and juveniles. Journal of Fish Biology, 67, 1094-1106. doi:10.1111/j.0022-1112.2005.00812.x
- Padrós, F., Minkoff, G., Sala, R., et al. (1993) Histological events throughout the development of turbot, Scophthalmus maximus larvae. Journal of Comparative Pathology, 109, 321-334. doi:10.1016/S0021-9975(08)80296-4
- Gisbert, E., Piedrahita, R.H. and Conklin, D.E. (2004) Ontogenetic development of the digestive system in California halibut, Paralichthys californicus with notes on feeding practices. Aquaculture, 232, 455-470. doi:10.1016/S0044-8486(03)00457-5
- Mousa, M.A. (2004) The effect of thyroxine on the activity of activin βA during the growth of the larvae of the Nile tilapia Oreochromis niloticus. Journal of Union of Arab Biologists Cairo, 22, 99-120.
- Yamano, K. (2005) The role of thyroid hormone in fish development with reference to aquaculture. Japan Agricultural Research Quarterly, 39, 161-168.
- Yen, P.M. and Chin, W.W. (1994) New advances in understanding the molecular mechanisms of thyroid hormone action. Trends in Endocrinology and Metabolism, 5, 65-72. doi:10.1016/1043-2760(94)90004-3
- Castro, L.F.C. and Wilson, J.M. (2011) Morphological diversity of the gastrointestinal tract in fishes. Fish Physiology, the Multifunctional Gut of Fish, 30, 1-55.
- Kurokawa, T., Iinuma, N., Unuma, T., et al. (2004) Development of endocrine system regulating exocrine pancreas and estimation of feeding and digestive ability in Japanese eel larvae. Aquaculture, 234, 513-525. doi:10.1016/j.aquaculture.2003.12.002
- Zaiss, M., Papadakis, I., Divanach, P., et al. (2006) Ontogeny of the digestive tract in shi drum, Umbrina cirrrosa reared using the mesocosm larval rearing system. Aquaculture, 260, 357-368. doi:10.1016/j.aquaculture.2006.06.001
- Hachero-Cruzado, I., Ortiz-Delgado, J.B., Borrega, B., et al. (2009) Larval organogenesis of flatfish brill Scophthalmus rhombus L: Histological and histochemical aspects. Aquaculture, 286, 138-149. doi:10.1016/j.aquaculture.2008.09.039
- Wegner, A., Ostaszewska, T. and Rożek, W. (2009) The ontogenetic development of the digestive tract and accessory glands of sterlet (Acipenser ruthenus L.) larvae during endogenous feeding. Reviews in Fish Biology and Fisheries, 19, 431-444. doi:10.1007/s11160-009-9111-8
- Conn, H.J. (1953) Biological stains. Williams and Wilkins Company, Baltimore.
- Bancroft, J.D. and Cook, H.C. (1984) Manual of histological techniques. Churchill Livingstone, New York.
- Ansal, M.D. and Kaur, K. (1998) Relative efficacy of dietary administration of 3,5,3’-triiodothyronine (T3) to different stages of an Indian major carp, Cirrhina mrigala (Hamilton): Growth and economics. Aquaculture Research, 29, 835-841. doi:10.1111/j.1365-2109.1998.tb01109.x
- Power, D.M., Llewellyn, L., Faustino, M., et al. (2001) Thyroid hormones in growth and development of fish. Comparative Biochemistry and Physiology—Part C: Toxicology & Pharmacology, 130, 447-459. doi:10.1016/S1532-0456(01)00271-X
- Gavlik, S., Albino, M. and Specker, J.L. (2002) Metamorphosis in summer flounder: Manipulation of thyroid status to synchronize settling behavior, growth, and development. Aquaculture, 203, 359-373. doi:10.1016/S0044-8486(01)00624-X
- Mousa, M.A., El-Gamal, A.S. and Khalil, M.-B.A. (2002) Effect of exogenous hormonal treatment on growth, survival and mucus cells activity during larval development of common carp, Cyprinus carpio. Journal of the Egyptian German Society of Zoology, 37, 193-207.
- Kang, D.-Y. and Chang, Y.J. (2004) Effects of maternal injection of 3,5,3’-triiodo-L-thyronine (T3) on growth of newborn offspring of rockfish, Sebastes schlegeli. Aquaculture, 234, 641-655. doi:10.1016/j.aquaculture.2004.01.011
- Lam, T.G., Juario, J.V. and Banno, J. (1985) Effect of thyroxine on growth and development in post-yolk sac larvae of milkfish, Chanos chanos. Aquaculture, 64, 179- 184. doi:10.1016/0044-8486(85)90203-0
- De Jesus, E.G., Toledo, J.D. and Simpas, M.S. (1998) Thyroid hormones promote early metamorphosis in grouper (Epinephelus coioides) larvae. General and Comparative Endocrinology, 112, 10-16. doi:10.1006/gcen.1998.7103
- Huang, L., Specker, J.L. and Bengtson, D.A. (1996) Effect of triiodothyronine on the growth and survival of striped bass (Morone saxatilis). Fish Physiology and Biochemistry, 15, 57-64. doi:10.1007/BF01874838
- Nugegoda, D., Walford, J. and Lam, T.J. (1994) Thyroid hormones in the early development of seabass (Lates calcarifer) larvae. Journal of Aquaculture in the Tropics, 9, 279-290.
- Nacario, J.F. (1983) The effect of thyroxine on the larvae and fry of Sarotherodon niloticus L. (Tilapia nilotica). Aquaculture, 34, 73-83. doi:10.1016/0044-8486(83)90292-2
- Ostaszewska, T., Wegner, A. and Wegiel, M. (2003) Development of the digestive tract of ide, Leuciscus idus (L.) during the larval stage. Archives of Polish Fisheries, 11, 181-195.
- Shehata, S.M.A. (1999) Studies on the anatomy and histochemistry of the digestive tract in the fingerling stages of the grass carp, Ctenopharyngdon idella (Valenciennes), in relation to food and feeding habits. Egyptian Journal of Aquatic Biology and Fisheries, 13, 113-145.
- Sarasquete, C., De Canales, M.L.G., Arellano, J.M., et al. (1996) Histochemical aspects of the yolk-sac and digestive tract of larvae of the Senegal sole, Solea senegalensis (Kaup, 1858). Histology and Histopathology, 11, 881-888.
- Boulhic, M. and Gabaudan, J. (1992) Histolgical study of the organogenesis of the digestive system and swim bladder of the Dover sole, Solea solea (Linneaus 1758). Aquaculture, 102, 373-396. doi:10.1016/0044-8486(92)90190-V
- Baglole, C.J., Murray, H.M., Goff, G.P., et al. (1997) Ontogeny of the digestive tract during larval development of the Yellow tail flounder: A light microscopy and mucous histochemical study. Journal of Fish Biology, 51, 120-134. doi:10.1111/j.1095-8649.1997.tb02518.x
- Yashpal, M., Kumari, U., Mittal, S., et al. (2007) Histochemical characterization of glycoproteins in the buccal epithelium of the catfish, Rita rita. Acta Histochemica, 109, 285-303. doi:10.1016/j.acthis.2007.03.002
- Morrison, C.M. and Wright, J.R. (1999) A study of the histology of the digestive tract of the Nile tilapia. Journal of Fish Biology, 54, 597-606. doi:10.1111/j.1095-8649.1999.tb00638.x
- Cardellini, P., Zanella, S., Franceson, A., et al. (1998) Differentiation of the digestive tract in the shi drum, Umbrina cirrosa (L.), a new fish recently reared in Mediterranean aquaculture. In: Enne, G. and Greppi, G.F. (Eds.), New Species for Mediterranean Aquaculture, Elsevier, Amsterdam, 183-196.
- Ribeiro, L., Sarasquete, C. and Dinis, M.T. (1999) Histological and histochemical development of the digestive system of Solea senegalensis (Kaup, 1858) larvae. Aquaculture, 171, 293-308. doi:10.1016/S0044-8486(98)00496-7
- Veggetti, A., Rowlerson, A., Radaelli, G., et al. (1999) Post-hatching development of the gut and larval muscle in the sole. Journal of Fish Biology, 55, 44-65. doi:10.1111/j.1095-8649.1999.tb01045.x
- Bisbal, G.A. and Bengston, D.A. (1995) Development of the digestive tract in larval summer flounder. Journal of Fish Biology, 47, 277-291. doi:10.1111/j.1095-8649.1995.tb01895.x
- Soffientino, B. and Specker, J.L. (2000) Age-dependent response of developing gastric mucosa to thyroxine (T4) in summer flounder. 4th International Symposium on Fish Endocrinology (Abstracts), ISFE W-461, 31 July-3 August 2000, University of Washington and Northwest Fisheries Science Center, Seattle.
- Osman, A.H.K. and Caceci, T. (1991) Histology of the stomach of Tilapia nilotica_Linnaeus, 1758. From the river Nile. Journal of Fish Biology, 38, 221-223. doi:10.1111/j.1095-8649.1991.tb03107.x
- Kjørsvik, E., Van der Meeren, T., Kryvi, H., et al. (1991) Early development of the digestive tract of cod larvae, Gadus morhua during start-feeding and starvation. Journal of Fish Biology, 38, 1-15. doi:10.1111/j.1095-8649.1991.tb03086.x
- Luizi, F.S., Gara, B., Shields, R.J., et al. (1999) Further description of the development of the digestive organs in Atlantic halibut, Hippoglossus hippoglossus larvae, with notes on differential absorption of copepod and Artemia prey. Aquaculture, 176, 101-116. doi:10.1016/S0044-8486(99)00054-X
- Kurokawa, T. and Suzuki, T. (1996), Formation of the diffuse pancreas and the development of digestive enzyme synthesis in the larvae of the Japanese flounder Paralichthys olivaceus. Aquaculture, 141, 267-276. doi:10.1016/0044-8486(95)01237-0
- Tanaka, M. (1973) Studies on the structure and function of the digestive system of teleost larvae. Ph.D. Thesis. Kyoto University, Kyoto, 136.
- Ma, H.M., Cahu, C., Zambonino I.J.L., et al. (2005) Activities of selected digestive enzymes during larval development of large yellow croaker (Pseudosciaena crocea). Aquaculture, 245, 239-248. doi:10.1016/j.aquaculture.2004.11.032