Journal of Behavioral and Brain Science
Vol.4 No.9(2014), Article
ID:49632,10
pages
DOI:10.4236/jbbs.2014.49039
Invariance of Functional Characteristics to Task Difference at Low-Level and High-Level Areas in the Ventral Visual Pathway
Yul-Wan Sung1,2, Uk-Su Choi2, Seiji Ogawa1,2
1Kansei Fukushi Research Institute, Tohoku Fukushi University, Sendai, Japan
2Neuroscience Research Institute, Gachon University, Incheon, Korea
Email: sung@tfu-mail.tfu.ac.jp
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 4 July 2014; revised 21 August 2014; accepted 3 September 2014
ABSTRACT
Tasks with stimuli are often used for the examination of stimulus-related functional characteristics of brain areas. However, the task can affect the response to a stimulus. Repetition suppression is a phenomenon that can be used to probe neuronal properties using macroscale functional magnetic resonance imaging (fMRI). The use of repetition suppression as an investigative tool to assess functional characteristics warrants the investigation of the invariance of repetition suppression to a given task. In this study, we examined repetition suppression using images of faces during different tasks. We found that the task difference did not change the response patterns related to repetition suppression in high-level areas and the primary visual area while it changed amplitudes of fMRI response to the visual stimuli. The result suggests that the repetition-suppression phenomenon is robust compared with the amplitude of fMRI response, and functional characteristics can be examined using the repetition-suppression phenomenon even under the condition that fMRI response is varied by task difference.
Keywords:Task Difference, Repetition Suppression, FFA, Central Stimulation, Peripheral Stimulation, fMRI, Face Images

1. Introduction
The repetition suppression observed in functional MRI (fMRI), which is also known as refractory suppression or fMRI adaptation, is a phenomenon in which the response to the second presentation of a visual stimulus decreases when the stimulus is presented repeatedly at short intervals. Refractory suppression is a suppression phenomenon that occurs between two input signals that arise successively within a short interval (<1000 ms). The response of a pair of identical stimuli is ideally twice that of one of the stimuli in the pair. The response to a pair of identical stimuli is about 20% larger than that to one of the stimuli in the pair with a short interval of 200 ms. However, the suppression recovers almost completely at 1000 ms, i.e., the response to paired stimulation becomes twice as large as that of one of the two stimuli in the pair [1] [2] . fMRI adaptation is a reduction in the response to a stimulus that is presented repeatedly with a relatively long interval, even up to several tens of seconds, i.e., the recovery in response takes longer in the case of fMRI adaptation than in the case of refractory suppression [3] [4] . Therefore, the latter can be used to assess the temporal characteristics of fast neuronal processing. However, in the case of two different stimuli in a pair, pair suppression can be weak, even with a very short interval (less than hundreds of milliseconds), when the common components of the two stimuli in the pair are small. Therefore, the phenomenon can also be used to assess the spatial characteristics of the distribution of different neuronal populations in a given brain area. This phenomenon has been used to examine neuronal properties via macroscale measurements using fMRI. In the context of refractory suppression, it is understood that the input signals to an area stem in a feed-forward manner from the lower area, and that signal suppression occurs via an interaction between the signals that arise successively.
Signal processing in the feed-forward manner has been found in the ventral visual pathway oriented for object processing which has a set of interconnected cortical regions from primary visual cortex (V1) to the temporal end. However, the processing in visual object areas in the ventral visual pathway is not solely dependent on feed-forward inputs. Many previous studies have reported that neuronal responses at V1 and high-order areas for object processing are enhanced in the presence of spatial attention [6] -[12] and that the response enhancement differs depending on visual field on which stimuli are presented [13] . Among those, Reddy et al. also reported that higher-order areas such as fusiform face area (FFA) show that an fMRI response to a task-relevant stimulus is enhanced compared with task-irrelevant stimuli [10] . They interpreted the task-relevant effect as top-down attention in that top-down attention is non-spatial and bottom-up is spatial. The response modulations by spatial and non-spatial (or bottom-up and top-down) attention give a possibility that an area in the ventral visual pathway get modulatory inputs from lateral areas or higher-order areas and the inputs affect refractory suppression for the area, that is, if an fMRI response to some stimulus is modulated by attention or an assigned task and the modulation of response reflects some changes of neuronal activity then the change of neuronal activity may lead to some modulation in refractory suppression. Although direct evidence of modulation about refractory suppression by attention does not exist, one of the characteristics of refractory suppression supports the possibility. It is that refractory suppression is strong in case that two stimuli in a stimulus pair elicit strong responses but week in case that both stimuli elicit weak responses or the second stimulus of two successive stimuli in the pair is weak [14] .
In this study, we tested whether fMRI response modulation by task difference accompanies with refractory suppression in relation with visual field differences (visual eccentricity).
For the purpose, we used task-irrelevant and task-relevant conditions. Under the task-irrelevant condition, participants performed color discrimination of a cross-hair at the center of the visual field while target face images are given at the central or peripheral visual fields. Under the task-relevant condition, participants performed an associative task composed of the color discrimination of the cross-hair and a gender discrimination of faces (Figure 1).
2. Materials and Methods
2.1. Subjects
Twenty-six healthy volunteers participated in this study. Thirteen subjects participated in the low-load and highload tasks, respectively. The participants were interviewed and their health was evaluated on the basis of physical examinations performed in the last 6 months of the experiment. All participants had normal or corrected-tonormal vision with 20/20 acuity, no history of neurological disease, and no medical conditions that would prevent MRI screenings, such as pregnancy, presence of a cardiac pacemaker, or claustrophobia. Subjects provided written informed consent after they were given a complete description of the study. This study was approved by the Institutional Review Board of Tohoku Fukushi University.
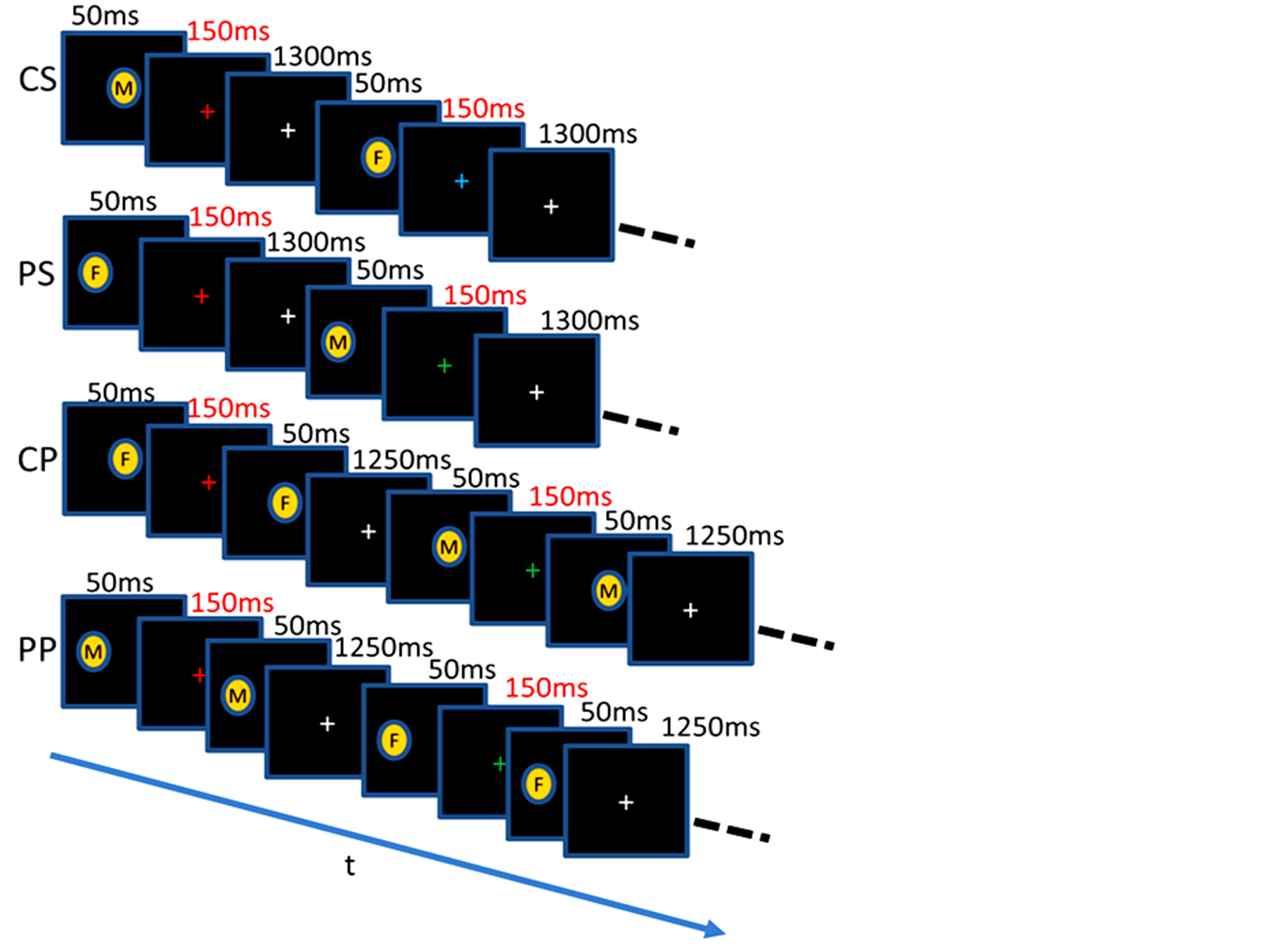
Figure 1. Stimulation conditions. There were 3 conditions for the central and peripheral stimulation. CS: Single central stimulation, CP: paired central stimulation, PS: single peripheral stimulation, PP: paired peripheral stimulation. F: female, M: male. Within the paired stimulation, faces were the same (SP) or different (DP). Each block included 8 stimuli with images female and male presented randomly. Color for the fixation task was changed randomly. The presentation time was 50 ms for each image and 150 ms for each colored crosshair. The same stimulation conditions were used for the lowand highload tasks. Under the task-irrelevant condition participants were required to detect color change of the crosshair by pressing button 1 for green and 2 for red. Under the task-relevant condition participants were required to identify the gender of the face presented in an image, in addition to the color change of the crosshair. Responses required participants to press button 1 for the combinations of gender and color, such as male and green or female and red, and button 2 for the other combinations.
2.2. Stimulation Procedure
The visual stimuli were images presented in the center of the visual field (central stimulation) or unilaterally in the left visual field (peripheral stimulation; Figure 1). There were 24 event blocks, with each block lasting 12 s. The 24 event blocks each consisted of 12 blocks of central and peripheral stimulation, respectively. The 12 blocks of central or peripheral stimulation comprised four blocks of single stimulation (S), paired stimulation with the same face (SP), and paired stimulation with different faces (DP) of the same but and different identities. The S, SP, and DP blocks were randomized at the intrasubject and intersubject levels. Therefore, there were three stimulus conditions for central and peripheral stimulation: S, SP, and DP (Figure 1). The experimental run started with a 12 s prestimulus state (for the removal of transient effects), and each stimulus block was followed by a 12 s control block; the total scan time was 600 s. The S block consisted of eight images of different faces that were presented for 50 ms and a 1.5 s repetition period interspersed with the control state. Each paired block (SP or DP) consisted of eight pairs of faces, with a 1.5 s interval between pairs. The interstimulus interval (ISI) of the SP and DP blocks was 150 ms. Images presented in the paired stimulation blocks were different between pairs, even in the SP block (Figure 1). During the scan, the colored crosshair was presented for 150 ms and the color was changed randomly to green or red with a 1.5 s interval. The onset of the colored crosshair occurred after the offset of the stimulus in the S block, and after that of the first stimulus in the SP and DP blocks. A gray crosshair was presented at the left period. The control block was the same as the stimulus block with the exception that there was no image presentation, i.e., the colored crosshair was presented for 150 ms and the gray crosshair period became longer, to match the period of face stimulation. The tasks in the stimulation state had two levels: task-irrelevant (T-I) and task-relevant (T-R). In the task-irrelevant condition, subjects were required to press buttons 1 or 2 according to the color of the crosshair (green or red, respectively). In the task-relevant condition, subjects were required to detect simultaneously the color changes of the crosshair and the gender of the individuals portrayed in the images, i.e., they had to press button 1 when the crosshair was green and the image showed a man, or when the crosshair was red and the image showed a woman, and press button 2 in any other case. The tasks in the control state were the same for both the T-I and T-R conditions and required detection of only a color change of the crosshair.
After this main experimental scan (refractory experiment), a localization scan (localization experiment) was performed to localize V1 and FFA in each subject. The localization scan included eight blocks. The duration of each block was 12 s. Out of these eight blocks, four blocks included images of faces, whereas four blocks included images of buildings that were interspersed with the 16-s control states and the 16 s poststimulus intervals. During the control state, a black crosshair was presented in the center of a gray background. Each event block consisted of eight different images. Each image in each block was presented for 1 s in the center of the visual field, and the same black crosshair was presented at an interimage interval of 0.5 s. The subjects performed a one-back task during the scan, in which they pressed button 1 when an image was repeated and pressed button 2 in all other scenarios.
2.3. Measurements
MRI experiments were performed using a Verio system (Siemens, Germany) with a standard, 12-channel head matrix coil operating at 3 Tesla. After the initial localization of images, T1-weighted anatomical images were obtained with an inverted recoveryand magnetization-prepared rapid acquisition using a gradient echo with a matrix size of 256 ´ 256 mm over a 256-mm field of view and a 1-mm slice thickness. For functional imaging, single-shot, gradient echo, planar imaging was performed using the following parameters: repetition time = 1000 ms, echo time = 25 ms, flip angle = 80˚, field of view = 220 mm, matrix size = 64 ´ 64 mm, and slice thickness = 3.4 mm, with no gap. Eighteen slices parallel to the anterior–posterior commissure line were acquired for each volume. The slice acquisition order was interleaved.
2.4. Visual Stimulation
The visual stimuli were grayscale images presented using a projector and displayed on a mirror mounted on the head coil (spatial resolution, 1024 ´ 768 pixels; refresh rate, 60 Hz; Panasonic, Japan). The images occupied 3.5˚ ´ 4˚ for the SP experiment, and the black crosshair was 0.34˚ ´ 0.34˚. The center of the images used for the peripheral stimulation was shifted 2˚ to the subject’s left. The reason parafovea was used for peripheral stimulation was to allow the discrimination of faces and facilitate fixation. The images were novel stimuli that were developed by the investigators. The images used for localization occupied 5˚ ´ 7˚ at the center of the visual field.
3. Results
The primary visual area V1 were identified near the posterior tip of the calcarine sulcus on activation map by a conjunction analysis of face and building stimulation in the localization experiments (Figure 2(a)). FFA was identified as a cluster of >200 voxels with a p-value of <0.001 around the fusiform gyrus by contrasting fMRI responses for the face and building stimulation in the localization experiment (Figure 2(b) and Figure 2(c)).
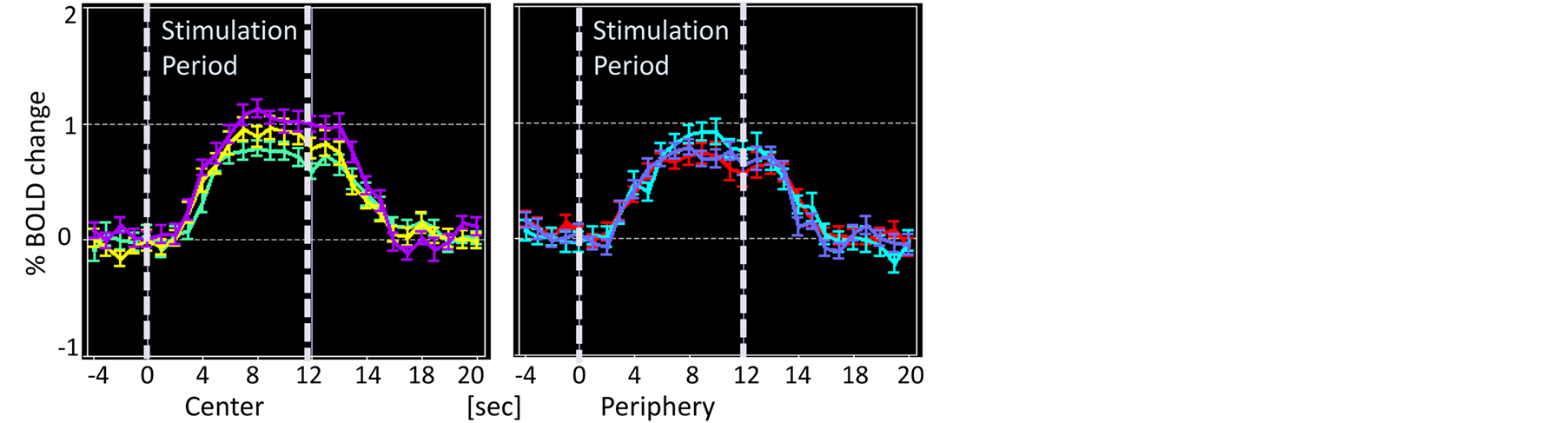
For analysis at each ROI, data (beta values) regarding fMRI signal changes were extracted from each V1 and FFA. FMRI signal time-courses at FFA were plotted for each task-irrelevant condition and task-relevant condition to see response profiles on the temporal axis (Figure 3). To examine the task effects on the stimulus, a



Figure 2. Activation maps by the localization experiment. (a) V1 (15, −95, −8) was identified by a conjunction analysis of the face and building conditions; (b) FFA (40, −49, −22) was identified by a contrast analysis of the face and building conditions; (c) FFA activation maps were seen at the z-direction to display the shape of activation site. The locations are based on Talairach coordinates. A, anterior; P, posterior; R, right; L, left. p < 0.001 (uncorrected).
 (a)
(a)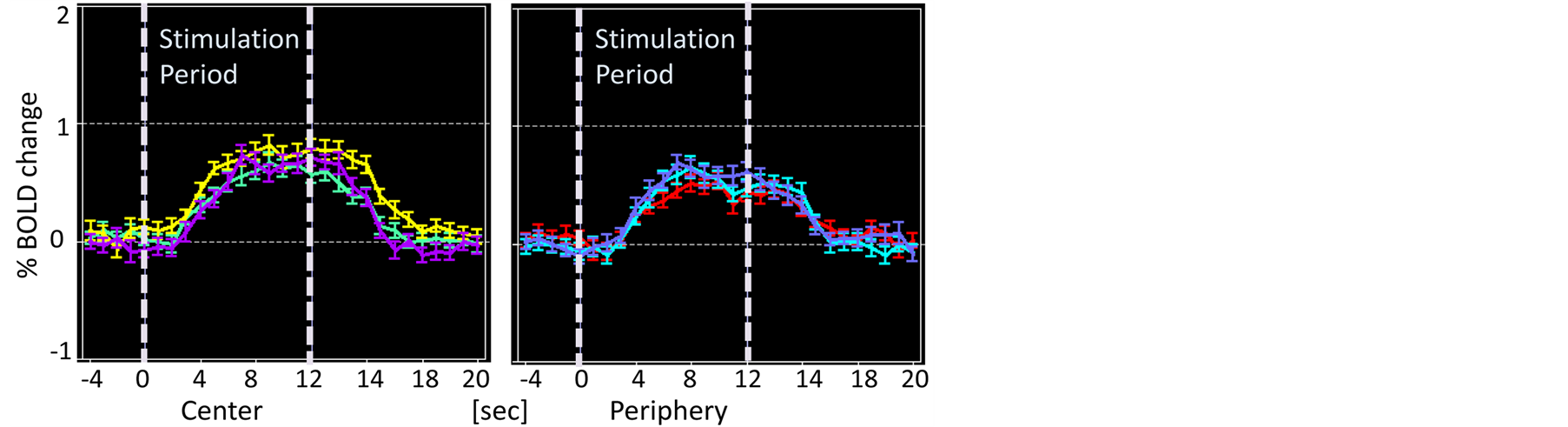 (b)
(b)
Figure 3. FMRI time-courses at FFA. (a) Task-irrelevant condition; (b) Task-relevant condition. The left panel of each figure is for “Central” stimulation and the right panel is for “Peripheral” stimulation. Green is for S, yellow is for SP and purple is for DP for “Central” stimulation, and red is for S, cyan is for SP and light purple is for DP for “Peripheral” stimulation.
four-way ANOVA was performed for the central stimulation using the factors of ROI (V1 and FFA), task (T-I and T-R), visual eccentricity (central and peripheral) and stimulus (S, SP, and DP) and the signal change as the dependent variable. This analysis revealed significant interactions between three factors of ROI, visual eccentricity and task [F(3, 228) = 11.33, p = 0.022], and significant main effect for the stimulus factor [F(2, 188) = 5.315, p = 0.001] (Figure 4). Therefore, two-way ANOVA was performed for each area (V1, FFA) with the factors of eccentricity and task.
The analysis at V1 revealed significant interaction between the eccentricity and task factors [F(1, 152) = 7.67, p = 0.006]. Further analysis regard to the task were not significant for the central stimulation [F(76) = 2.48, p = 0.119] but significant for the peripheral stimulation, that is, task-irrelevant (T-I) < task-relevant (T-R) [F(76) = 5.36, p = 0.023] (Figure 5). The analysis at FFA revealed no significant interaction between the visual eccentricity and task factors [F(1, 152) = 1.66, p = 0.200]. The main effect of the task factor was significantly different between T-I and T-R (T-I < T-R) [F(1, 152) = 10.11, p = 0.002] and the main effect of the visual eccentricity factor was significantly different between the central and peripheral stimulation (central stimulation > peripheral stimulation) [F(1, 152) = 22.35, p = 0.001] (Figure 6).

Figure 4. Averaged responses of three stimulus conditions of S, SP and DP. **: p < 0.001. The error bars represent S.E.M.

Figure 5. Responses of task-irrelevant and task-relevant conditions for central and peripheral stimulation (visual eccentricity factor) respectively. (a) Central stimulation; (b) Peripheral stimulation. *: p < 0.03. n.s.: not significant. The error bars represent S.E.M.
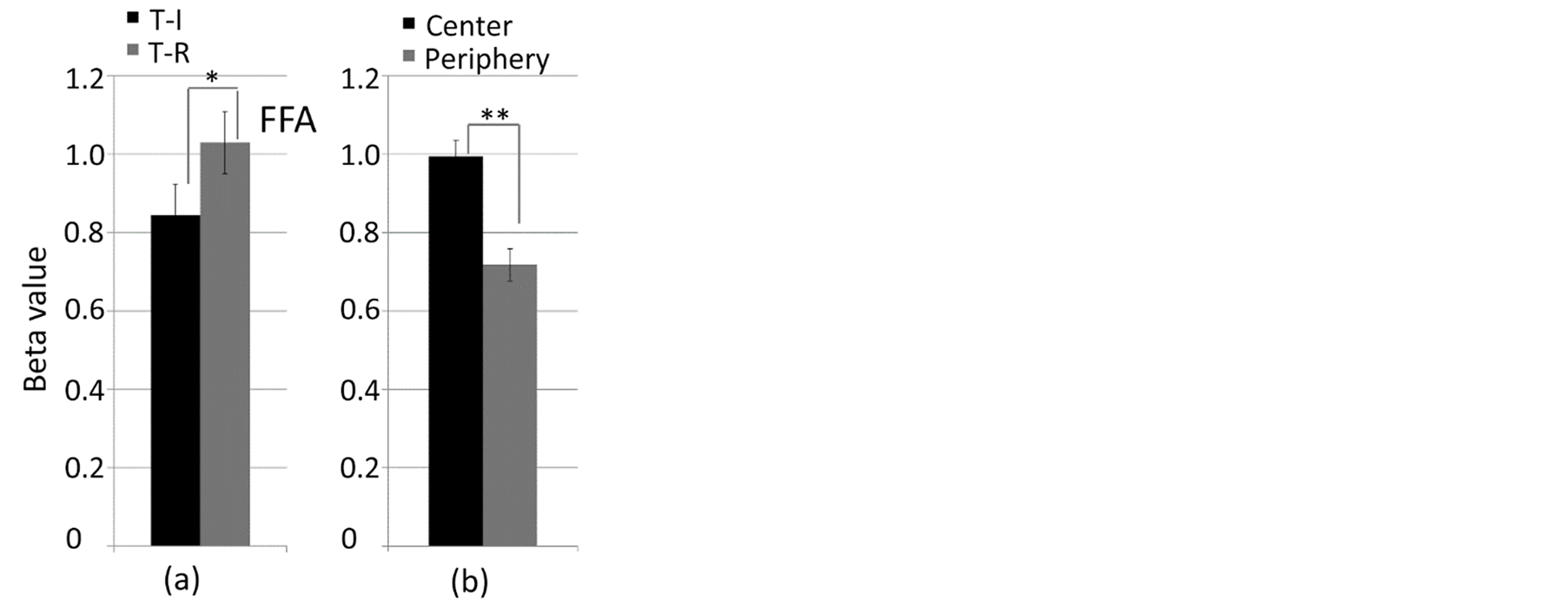
Figure 6. Responses for task-difference and visual eccentricity factors (a) task-irrelevant and task-relevant conditions (b) central and peripheral stimulation. *: p < 0.002, **: p < 0.001. The error bars represent S.E.M.
4. Discussion
The results that task difference was not reflected at V1 and FFA whether the stimuli were presented at the center or the periphery of the visual field, while task difference modulates fMRI response to the peripheral stimulation at V1 and to both central and peripheral stimulation at FFA, indicate that refractory suppression is more robust phenomenon than fMRI signal. Under the conditions of the present experiment, the results reject the hypothesis that fMRI response modulation by task difference accompanies with refractory suppression.
The larger response to task-relevant stimulation presented at peripheral visual field of V1 can be explained in relation with the distance from the fixation point to the location of target stimulus images. Amplitude of fMRI signal to unattended target stimulus images is known to decrease as the distance of the location of the stimulus images from the fixation point increases when attention is given to the fixation point [11] . This response decrease in task-irrelevant condition and a response increase in task-relevant condition by spatial attention might make significant difference between task-irrelevant and task-relevant conditions.
Enhancement in fMRI response at FFA for the task-relevant condition is consistent with the previous studies [6] [10] . The response enhancement at FFA can also be explained in relation with the studies of spatial attention because there would be partial increase in attention to the target stimulus face images in the task-relevant condition [7] -[9] [15] [16] . The response difference at FFA between the central and peripheral stimulation is consistent with the preference of the central vision of FFA [17] .
Object processing varies depending on not only stimulus-driven selection but also other factors such as visual eccentricity (or positon) and top-down selection [5] [18] -[20] . However, it is unclear yet to what extent those factors contribute to visual object processing. The present study may give some information about the effect of those factors on visual object processing; the effect by those factors to neuronal activity underlying the refractory suppression is not much as to affect an average activity in brain areas even if it exists.
In regard to signal processing modality, we used univariate analysis. Our results derived from evaluation of an average activity in a brain area rejected the hypothesis that refractory suppression would be modulated by task difference but it does not deny the possibility of acceptance of the hypothesis in case that multivariate analysis is applied [21] -[25] . However, most of the studies on repetition suppression is based on an average activity in brain areas and our objective in this study is also to examine whether the modulation occurs at the level of the average activity [1] [26] -[31] . Moreover, the experimental design in this study was not oriented to multivariate analysis, so that data processing by multivariate analysis is not appropriate. To examine refractory suppression by multivariate analysis will be our future study.
These, all together, suggest that the refractory suppression is robust to the task difference compared to the amplitude of fMRI signal.
Ideally it may be best to measure refractory suppression under a passive condition in which participants focus on the center of the visual field while stimulus images are given at some locations of the visual field because refractory suppression is known to reflect an interaction between feed-forward inputs. However, under the passive condition participants tends to lose their attention or fall asleep in the MRI scanner. Therefore, a task is needed to keep participants’ arousal state or attention constant, and the task should be exclusively independent on target stimulus. However, the task, even if it is independent, the response to the target stimulus can be affected [8] [10] [11] . Therefore, the finding in this study that the refractory suppression is robust to the task difference would be useful information in applying the refractory suppression to studies of functional characteristics of brain areas.
In regard to the refractory suppression, one hypothesis is that the response to a paired stimulus is ideally twice the response to one stimulus (single) in the pair when ISI is sufficiently long, i.e., around 1000 ms. According to this hypothesis, a response to a paired stimulus is regarded as suppressed if the response is less than twice of response to a single stimulus. Another hypothesis for refractory suppression is that the recovery from suppression (that is, for response to a pair to reach twice the response to a single stimulus) is faster for a paired stimulus when the paired stimulus consists of different types of stimuli. Previous studies on refractory suppression have not described explicitly the “twice” value in the evaluation of response suppression as the criterion for suppression. However, it remains uncertain whether the criterion can be affected by experimental conditions and the fully recovered response does not reach twice the value of the response to a single stimulus. In this context, a previous study showed that the refractory suppression at V1 by checker pattern stimulation had almost completely recovered after a 1000 ms interval [1] ; moreover, in our supplementary experiment using face images, we observed that the refractory suppression at FFA had recovered by >80% (i.e., a factor of 1.8 relative to the single stimulus), even with a short interval of 532 ms, whereas the recovery was about 40% for an interval of 132 ms when different faces were used in a pair. Therefore, it is reasonable to regard the double of the value of the single stimulus as the criterion for suppression. Those provide the validity to the implicit criterion for refractory suppression. Thus, it is reasonable to think that the responses to SP and DP at V1 and FFA, in which responses to SP or DP are less than twice that to the corresponding S, are regarded as suppressed. In the present study, the suppression for SP was weak compared with those reported by other studies that were conducted using similar ISIs [32] . One of the reasons for this discrepancy may be the switching of the location of stimulation between the center and the periphery in the same stimulus run, although the interval between the stimulus blocks was long [33] .
Next, we will describe the expected criticism regarding the intersubject design of and ROI identification in this study. One explanation for the use of the intersubject design to examine the effect of task condition was to avoid any possible interaction between task-irrelevant and task-relevant conditions when participants perform two tasks in the same experimental session, one of considered is that participants may perform the task under task-relevant condition implicitly while they are performing under task-irrelevant conditon explicitly. Another is that the scan time (600 s) and the performance of two tasks in one session can yield data with a worse quality. In fact, in our preliminary experiment in which two 600 s scans were performed, the data for the second scan contained much more physiological fluctuation and variation in response amplitudes than did the first scan.
For the localization of an ROI for FFA, the possibility that the ROI of FFA was shifted to an area preferring central stimulation should be considered, because face images were presented at the central visual field in the localization experiment. Even if there was a bias in FFA localization, which was shifted in the direction of the fovea due to the presentation of images at the center it does not defy the results because the effects of task condition and repetition suppression at FFA were evaluated separately for the central and periphery condition. Therefore, the definition of ROI based on the central presentation of images does not have an impact on our results.
5. Conclusion
The task difference did not affect the suppression phenomenon although fMRI response amplitudes are modulated by the task difference. This suggests that the refractory suppression is robust to the task difference than fMRI signal. This also suggests that task difference does not have to be considered, as long as univariate analysis is applied, when examining the suppression phenomenon, regardless of the presentation of the stimulation in the center or the periphery of the visual fields, and that the suppression phenomenon can be used as a tool for probing functional characteristics at both high-level and low-level areas.
Acknowledgements
This work was supported by JSPS KAKEN Grant Number 26350995 and 25330173, and MEXT-Supported program for the Strategic Research Foundation at Private Universtities, 2014-2018.
References
- Ogawa, S., Lee, T.-M., Stepnoski, R., Chen, W., Zhu, X.-H. and Ugurbil, K. (2000) An Approach to Probe Some Neural Systems Interaction by Functional MRI at Neural Time Scale down to Milliseconds. Proceedings of the National Academy of Sciences, 97, 11026-11031. http://dx.doi.org/10.1073/pnas.97.20.11026
- Waldeck, R., Pereda, A. and Faber, D.S. (2000) Properties and Plasticity of Paired-Pulse Depression at a Central Synapse. The Journal of Neuroscience, 20, 5321-5320.
- Henson, R.N. and Rugg, M.D. (2003) Neural Response Suppression, Haemodynamic Repetition Effects, and Behavioral Priming. Neuropsychologia, 41, 263-270. http://dx.doi.org/10.1016/S0028-3932(02)00159-8
- Grill-Spector, K., Henson, R. and Martin, A. (2006) Repetition and the Brain: Neural Models of Stimulus-Specific Effects. Trends in Cognitive Sciences, 10, 14-23. http://dx.doi.org/10.1016/j.tics.2005.11.006
- Kravitz, D.J., Vinson L.D. and Baker C.I. (2008) How Position Dependent Is Visual Object Recognition? Trends in Cognitive Sciences, 12, 114-122. http://dx.doi.org/10.1016/j.tics.2007.12.006
- Harel, A., Kravitz D.J. and Baker C.I. (2014) Task Context Impacts Visual Object Processing Differentially across the Cortex. Proceedings of the National Academy of Sciences, in press.
- Kanwisher N. and Wojciulik E. (2000) Visual Attention: Insights from Brain Imaging. Nature Reviews Neuroscience, 1, 91-100. http://dx.doi.org/10.1038/35039043
- O’Conner, D.H., Fukui, M.M., Pinsk, M.A. and Kastner, S. (2002) Attention Modulates Responses in the Human Lateral Geniculate Nucleus. Nature Neuroscience, 5, 1203-1209. http://dx.doi.org/10.1038/nn957
- O’Craven, K.M., Downing P.E. and Kanwisher N. (1999) fMRI Evidences for Objects as the Units of Attentional Selection. Nature, 401, 584-587. http://dx.doi.org/10.1038/44134
- Reddy, L., Moradi, F. and Koch C. (2007) Top-Down Biases Win against Focal Attention in the Fusiform Face Area. Neuroimage, 28, 730-739. http://dx.doi.org/10.1016/j.neuroimage.2007.08.006
- Sprague, T.C. and Serences, J.T. (2013) Attention Modulates Spatial Priority Maps in the Human Occipital, Parietal and Frontal Cortices. Nature Neuroscience, 16, 1879-1887. http://dx.doi.org/10.1038/nn.3574
- Yantis, S. (2008) The Neural Basis of Selective Attention. Current Directions in Psychological Science, 17, 86-90.http://dx.doi.org/10.1111/j.1467-8721.2008.00554.x
- Davidesco, I., Harel, M., Ramot, M., Kramer, U., Kipervasser, S., et al. (2013) Spatial and Object-Based Attention Modulates Broadband High-Frequency Responses across the Human Visual Cortical Hierarchy. The Journal of Neuroscience, 16, 1228-1240. http://dx.doi.org/10.1523/JNEUROSCI.3181-12.2013
- Ogawa, S., Sung, Y., Kamba, M. and Nagaoka, T. (2003) Probing the Temporal Dynamics of the Functional System Interaction in the Brain by Functional MRI. Advances in Neurological Sciences, 47, 902-908.
- Bressler, D., Fortenbaugh, F., Robertson, L. and Silver, M. (2013) Visual Spatial Attention Enhances the Amplitude of Positive and Negative fMRI Responses to Visual Stimulation in an Eccentricity-Dependent Manner. Vision Research, 85, 104-112. http://dx.doi.org/10.1016/j.visres.2013.03.009
- Bressler, D. and Silver, M. (2010) Spatial Attention Improves Reliability of fMRI Retinotopic Mapping Signals in Occipital and Parietal Cortex. Neuroimage, 53, 526-533. http://dx.doi.org/10.1016/j.neuroimage.2010.06.063
- Levy, I., Hasson, U., Avidan, G., Hendler, T. and Malach, R. (2001) Center-Periphery Organization of Human Object Areas. Nature Neuroscience, 4, 533-539.
- Hemond, C., Kanwisher, N. and Op de Beeck, H. (2007) A Preference for Contralateral Stimuli in Human Object-and Face-Selective Cortex. PLoS ONE, 2, e574. http://dx.doi.org/10.1371/journal.pone.0000574
- Pernet, C., Schyns, P. and Demonet, J.-F. (2007) Specific, Selective or Preferential: Comments on Category Specificity in Neuroimaging. Neuroimage, 35, 991-997. http://dx.doi.org/10.1016/j.neuroimage.2007.01.017
- Sowden, P.T. and Schynes, P.G. (2006) Channel Surfing in the Visual Brain. Trends in Cognitive Sciences, 10, 538-545. http://dx.doi.org/10.1016/j.tics.2006.10.007
- Haynes, J.D. and Rees, G. (2006) Decoding Mental States from Brain Activity in Humans. Nature Reviews Neuroscience, 7, 523-534. http://dx.doi.org/10.1038/nrn1931
- Mourao-Miranda, J., Bokde, A.L.W. and Born, C. (2005) Classifying Brain States and Determining the Discriminating Activation Patterns: Support Vector Machine on Functional MRI Data. Neuroimage, 28, 980-995.http://dx.doi.org/10.1016/j.neuroimage.2005.06.070
- Norman, K.A., Polyn, S.M., Detre, G.J. and Haxby, J.V. (2006) Beyond Mind-Reading: Multi-Voxel Pattern Analysis of fMRI Data. Trends in Cognitive Sciences, 10, 424-430. http://dx.doi.org/10.1016/j.tics.2006.07.005
- Johnson, J.D., McDuff, S.G.R., Rugg, M.D. and Norman, K.A. (2009) Recollection, Familiarity and Cortical Reinstatement: A Multivoxel Pattern Analysis. Neuron, 63, 697-708. http://dx.doi.org/10.1016/j.neuron.2009.08.011
- Schyns, P.G., Gosselin, F. and Smith, M.L. (2008) Information Processing Algorithms in the Brain. Trends in Cognitive Sciences, 13, 20-26. http://dx.doi.org/10.1016/j.tics.2008.09.008
- Eger, E., Henson, R.N., Driver, J. and Doaln, R.J. (2004) BOLD Repetition Decreases in Object-Responsive Ventral Visual Areas Depend on Spatial Attention. Journal of Neurophysiology, 92, 1241-1247. http://dx.doi.org/10.1152/jn.00206.2004
- Grill-Spector, K. and Malach, R. (2001) fMR-Adaptation: A Tool for Studying the Functional Properties of Human Cortical Neurons. Acta Psychologica, 107, 293-321. http://dx.doi.org/10.1016/S0001-6918(01)00019-1
- Henson, R.N., Shallice, T., Gorno-Tempini, M.L. and Dolan, R.J. (2002) Face Repetition Effects in Implicit and Explicit Memory Tests as Measured by fMRI. Cerebral Cortex, 12, 178-186. http://dx.doi.org/10.1093/cercor/12.2.178
- James, T.W. and Gauthier, I. (2007) Repetition-Induced Changes in BOLD Response Reflect Accumulation of Neural Activity. Human Brain Mapping, 27, 37-46. http://dx.doi.org/10.1002/hbm.20165
- Schweinberger, S.R., Huddy, V. and Burton, A.M. (2004) N250r: A Face-Selective Brain Response to Stimulus Repetitions. Neuroreport, 15, 1501-1505. http://dx.doi.org/10.1097/01.wnr.0000131675.00319.42
- Winston, J.S., Henson, R.N., Fine-Goulden, M.R. and Dolan, R.J. (2004) fMRI-Adaptation Reveals Dissociable Neural Representations of Identity and Expression in Face Perception. Journal of Neurophysiology, 92, 1830-1839.http://dx.doi.org/10.1152/jn.00155.2004
- Sung, Y., Someya, Y., Eriko, Y., Choi, S.-H., Cho, Z.-H., and Ogawa, S. (2011) Involvement of Low-Level Visual Areas in Hemispheric Superiority for Face Processing. Brain Research, 1366, 54-59. http://dx.doi.org/10.1016/j.brainres.2010.10.020
- Rauss, K., Pourtois, G., Vuilleumier, P. and Schwartz, S. (2009) Attentional Load Modifies Early Activity in Human Primary Visual Cortex. Human Brain Mapping, 30, 1723-1733. http://dx.doi.org/10.1002/hbm.20636