Agricultural Sciences
Vol.07 No.11(2016), Article ID:72017,22 pages
10.4236/as.2016.711072
Effect of Combined Application of Subsurface Drainage and Mineral Fertilization on Iron-Reducing Bacterial Populations’ Developments and Fe2+ Uptake by Two Rice Varieties in an Iron Toxic Paddy Soil of Burkina Faso (West Africa)
Cécile Harmonie Otoidobiga1*, Honoré Kam2, Adama Bagayogo2, Aboubacar Savadogo3, Joseph B. Sawadogo1, Sayouba Sawadogo4, Adama Sawadogo1, Yapi Sinaré2, Ibrahima Ouédraogo2, Prosper Zombré4, Susumu Asakawa5, Alfred S. Traoré1, Dayéri Dianou3
1Research Center for Biological, Alimentary and Nutritional Sciences, Research and Training Unit, Life and Earth Sciences, University of Ouagadougou, Ouagadougou, Burkina Faso
2Institute of Environment and Agricultural Research, Bobo-Dioulasso, Burkina Faso
3National Center for Sciences and Technology Research, Ouagadougou, Burkina Faso
4Laboratory of Soil-Materials and Environment, Research and Training Unit, Life and Earth Sciences, University of Ouagadougou, Ouagadougou, Burkina Faso
5Soil Biology and Chemistry, Graduate School of Bioagri Cultural Sciences, Nagoya University, Nagoya, Japan

Copyright © 2016 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: September 19, 2016; Accepted: November 13, 2016; Published: November 16, 2016
ABSTRACT
Rice is one of the staple crops in Burkina Faso. However, the local production covers only 47% of the population demands. One of the main reasons of the poor productivity in Burkina Faso is iron toxicity which is related mainly to the activity of Iron Reducing Bacteria in the rice field’s ecosystems. In order to control the harmful effects of Iron Reducing Bacterial populations and to improve rice productivity, a pots experiment was conducted at the experimental site of the University Ouaga I Pr. Joseph KI-ZERBO. An iron toxic soil from Kou Valley (West of Burkina Faso) and two rice varieties, BOUAKE-189 and ROK-5, sensitive and tolerant to iron toxicity, respectively, were used for the experiment. The pots were drained for 14 days (D2) and amended with chemical fertilizers (NPK + Urea and NPK + Urea + Ca + Mg + Zn complexes). Control pots without drainage and fertilization (D0/NF) were prepared similarly. The kinetics of Iron Reducing Bacterial populations and ferrous iron content in soil near rice roots were monitored throughout the cultural cycle using MPN and colorimetric methods, respectively. The total iron content was evaluated in rice plant using a spectrometric method. Data obtained were analyzed in relation to drainage and fertilization mode, rice growth stage and rice yield using the Student’s t-test and XLSTAT 2014 statistical software. The experiment showed that the combined application of subsurface drainage and NPK + Urea + Ca + Mg + Zn fertilization, reduced significantly the number of IRB in the soil near rice roots for both rice varieties (p = 0.050 and p = 0.020) increased the leaf tissue tolerance to excess amounts of Fe, and rice yield.
Keywords:
Iron Reducing Bacteria, Rice Variety, Iron Uptake, Subsurface Drainage, Fertilization

1. Introduction
Rice is the fourth staple crop after sorghum, millet and maize in Burkina Faso [1] . However, the local production of rice covers only 47% of the population demands. To meet the increasing requirements of rice, the Government of Burkina Faso imports about 260,000 tonnes of rice per year, with more than 30 milliards of CFA francs of currency losses [2] . Sikirou et al. [3] reported that one reason for the poor productivity in wetlands in West Africa is the prevalence of biotic and abiotic stresses. The abiotic stresses include drought, submergence and iron toxicity [4] [5] [6] . The iron toxicity is recognized as one of the most widespread nutritional disorders and one of the major edaphic constraints of lowlands in West Africa that affects crop growth, especially rice growth [7] [8] [9] [10] . Haefele et al. [11] estimated that 19% of the total rice area in Africa has a potential risk of Fe toxicity [3] . Chérif et al. [5] reported also that about 55% of the rice area is affected by Fe toxicity in three West African countries (Guinea, Ivory Coast and Ghana), with about 10% of rice cultivation area abandoned due to severe iron toxicity.
In Burkina Faso, many lowland crop fields were even abandoned due to iron toxicity [12] . Iron toxicity affects many agricultural plains in Burkina Faso like Moussodougou, Tiéfora and Kou Valley [13] [14] . For the latter, since 1986, 300 ha of fields were aban- doned because of ferrous intoxication [12] , and most among these intoxicated fields remained uncultivated up to date [14] .
Iron toxicity is a major nutritional disorder affecting rice production in irrigated and rainfed lowland soils and occurs as a consequence of the reduction of insoluble Fe3+ into soluble Fe2+ under both anaerobic and low pH conditions [10] [15] [16] . Indeed, most mineral soils are rich in Fe, but under aerobic conditions, Fe3+ biodisponibility remains low. In contrast, in flooded soils, Fe2+ is quickly absorbed by plant roots and can lead to cellular Fe overload, inducing numerous metabolic disorders [17] .
Iron reduction into ferrous form is largely controlled by microbial processes [18] . In fact, in natural systems, Fe (III) minerals can be reduced by strictly anaerobic or facultative Fe-reducing bacteria using a wide range of organic compounds or H2 as electron donors [19] [20] . As soon as a soil becomes flooded, the dissolved oxygen is consumed by aerobic bacteria and chemical oxidation reactions. Oxygen is depleted fast in most regions of the soil and alternative electron acceptors are used [7] . In anoxic conditions, Fe (III)-compounds are reduced and it results in ferrous ions production. Thus, the IRB population produces Fe2+ by coupling the Fe(III) reduction to the oxidation of substrates to support growth [21] [22] [23] [24] [25] .
Therefore, iron toxicity is a condition caused by the microbial reduction under flooded conditions of insoluble iron-III into soluble iron-II, which can be taken up by rice plants in excess amounts [18] .
Iron toxicity occurs when the rice plant accumulates a toxic concentration of Fe in the leaves [26] [27] [28] . High concentrations of Fe in soil solution also decrease the absorption by the rice plant of other plant nutrients, especially Phosphorus and Potassium [10] [29] . Excessive Fe uptake results in increased polyphenol oxidase activity, leading to the production of oxidized polyphenols, the cause of leaf bronzing. Large amounts of Fe in plants can give rise to the formation of oxygen radicals, which are phytotoxic and responsible for protein degradation and peroxidation of membrane lipids [30] [31] .
The symptoms of iron toxicity vary with rice cultivar [30] . The excess Fe is accumulated as brown dots in plant cells, causing bronzing symptoms of leaves, blackening of roots and damage to cellular membranes [32] [33] [34] [35] .
Benckiser et al. [36] and Becker and Asch [10] reported that iron-induced yield is frequently associated with a poor nutrient status of the soil. Kosaki and Juo [37] underlined also that the poor level of development of lowlands in West Africa, epitomized with bad water management which generates favourable conditions for the occurrence of iron toxicity [38] .
The present experiment was developed in the sensitive site of Kou Valley, to determine the effects of chemical fertilization and subsurface drainage on microbiological and chemical parameters sustaining iron toxicity in paddy fields and on rice yield. Therefore, plastics pots were filled with a sensitive soil, amended by chemicals fertilizers and drained periodically during rice cultivation. The Iron Reducing Bacterial (IRB) populations’ density, Fe2+ content in the paddy soil and iron accumulation in rice plant, were recorded during the cultural cycle of BOUAKE-189 and ROK-5 rice varieties (sensitive and tolerant to iron toxicity, respectively).
2. Materials and Methods
2.1. Soil Sampling and Experiments Sites Location
The soil used for the experiments was collected at Kou Valley, a site located at the West of Burkina Faso (11˚23'12 N and 4˚23'25'' W) (Figure 1). The physical and chemical properties of the soil used in the experiments were described in our previous report [39] .
The soil collected was carried out to the experimental site of the University Ouaga I Pr. Joseph KI-ZERBO (12˚22'45.7'' N and 1˚29’52.5'' W, Figure 1). The experiments were performed from June to October 2014.
2.2. Climatic Characteristics of the Experimental Site Area
The experiments site was located in the central plateau of Burkina Faso consisting of a rainy season (June-October) with peaks from July to September, and a dry season (November-May). During the experiments period (June-October 2014), sixty-two rainy days were recorded, with an average rainfall of 743.5 mm (Figure 2). The average daily temperature was 29.03˚C with minimum at 27.6˚C and maximum at 30.7˚C (Figure 3). The daily relative hygrometry (RH), varied from 45% to 77% (Figure 3), with a sunniness average of 7.26 hours/day.
2.3. Plant Material
Plant materials used for experiments were BOUAKE-189 and ROK-5 rice varieties, originating from the Asian species Oryza sativa L (Indica varietal group), respectively [40] . Native from Indonesia, BOUAKE-189 rice variety, is adapted to the irrigated rice, and is issued from the breeding selection of the National Agricultural Research Center (CNRA, Ivory Coast). This rice has an average yield estimated to 4.5 tons/ha, with a cy-
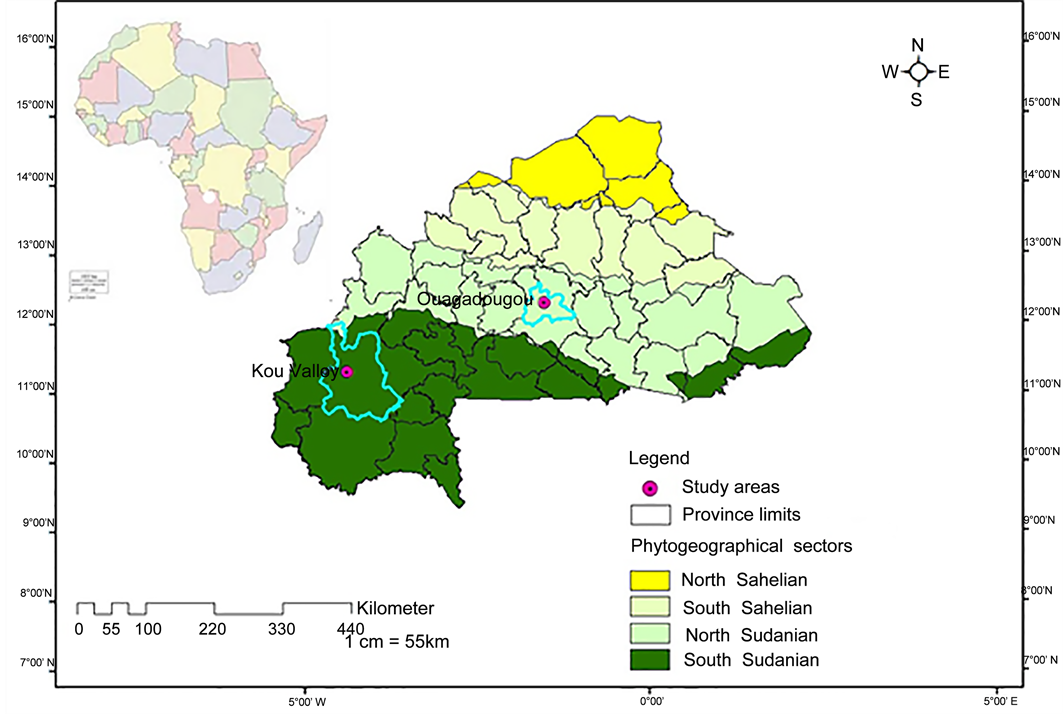
Figure 1. Location of sampling and experimental sites.
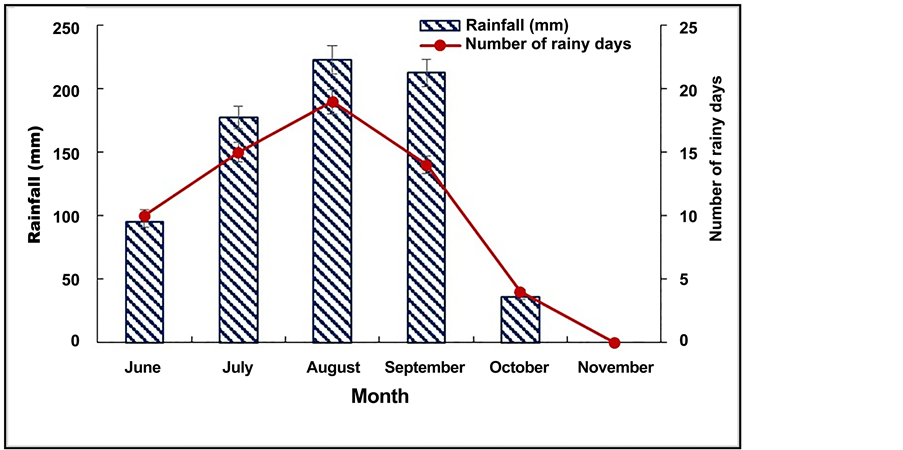
Figure 2. Monthly rainfall and monthly number of rainy days from June to November 2014.
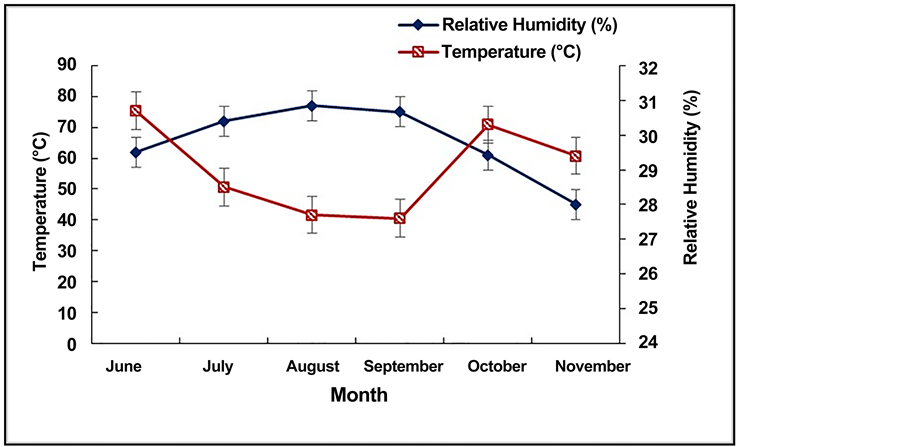
Figure 3. Monthly temperatures and monthly relative humidity from June to November 2014.
cle of seedling-maturity of 125 to 130 days. With an average yield estimated to 5 tons/ ha, the ROK-5 rice variety is an inbred rice issued from the West Africa Rice Development Association (WARDA, Rokupr, Sierra Leone) and adapted to mangrove rice.
2.4. Pots Experiments
Experiments were carried out as previously described by Otoidobiga et al. [39] . Seventy two plastics pots with 25 cm3 of bulk were used in 3 replications throughout the study. At the bottom of each pot, an external tap was installed to sub-drain the soil. After 2 weeks of flooding, 15 day-old rice plants were transplanted. Two rice varieties, BOUAKE-189 [38] and ROK-5 [41] , sensitive and resistant to iron toxicity, respectively were used. The soil was continuously flooded until rice maturity and harvest (120 days after flooding). Three replications and three modes of fertilization were performed throughout the study: without fertilization, NPK + Urea and NPK + Urea + Zn + Ca + Mg, respectively. The doses of N-P-K (14 - 23 - 14), CaCO3, ZnO and MgCl2 application in pots were in the ratio of 720:50:22.4:20 mg/kg of dry soil according to the recommended doses of 300 kg/ha for N-P-K, 10 kg/ha for ZnO, 250 kg/ha for CaCO3 and 8.92 kg/ha of MgCl2 at the rice transplanting [1] [36] [38] - [43] . The Urea fertilizer (240 mg/kg of dry soil) was applied in two further dressings (at rice transplanting, and 60 days after transplanting, respectively) according to the recommended dose of 100 kg/ha [1] . Two modes of drainage were applied during the study: without drainage (D0) and drainage for 14 days (D2), respectively by regulating the drained water flow from the bottom [18] as recommended by our previous results [18] . Control pots, without drainage and fertilization (D0/NF) were prepared similarly.
2.5. Iron Reducing Bacterial Populations Monitoring
The concentration of Iron Reducing Bacteria (IRB) in the soil were determined by the most-probable-number (MPN) method, and a culture medium adapted from Hammann and Ottow [44] consisting of Glucose (20 g), Sodium acetate (5 g), MgSO4· 7H2O (0.05 g), K2HPO4 (0.25 g), KH2PO4 (0.25 g), NaCl (0.05 g), Na2- MoO4·2H2O (0.005 g), CaCO3 (5 g), Fe2O3 (1 g) per liter of distilled water. The pH was adjusted to 7.2 by addition of NaOH. The medium (9 ml) was dispensed into 16 ml Hungate tubes and autoclaved at 121˚C for 15 min. One milliliter of the 10−1 to 10−9 fold of diluted soil suspension was inoculated to 9 ml of the basal medium for enumeration in Hungate tubes. Tubes were incubated at 30˚C for five days. Formation of reddish coloration after addition of a reagent containing 0.2% ortho-phenan- troline and 10% acetic acid, sustaining the reduction of Fe3+ (Fe2O3) into Fe2+ (reddish coloration) was used for detection of positive tubes after incubation period. The most probable numbers of IRB were calculated from a table of MPN for three tubes. The enumeration of bacteria was performed before flooding when the soil was dried, on transplanting day (two weeks after flooding) and during the rice growth stages until harvest near rice roots [39] .
2.6. Determination of Ferrous Iron Concentration in Soil
From the soil sample (2 g) for bacterial enumeration and at the same periods during the rice cultural cycle, the ferrous iron was extracted using extraction medium (AlCl3 0.5%), according to the method of Vizier and Blanch [45] . The extracted ferrous iron solution (80 µl) was removed by micropipette and rapidly introduced into microplate containing 20 µl of a reagent containing 0.2% ortho-phenantroline and 10% acetic acid. Immediately after mixing (for 5 s), the iron content of the soil was determinated by measuring the absorbance at 510 nm using a Spectronic 61 photometer.
2.7. Determination of Fe Content in Rice Plant
The Total Fe was analysed in the leaves and roots of the two rice varieties described above. Young leaves were taken from each pot during the cultural cycle of rice. At harvest, the aerial biomass and the roots of each plant were also collected. Completely dried plant parts were digested in a diacid mixture of nitric acid (HNO3) and perchloric acid (HClO4) (15:2) [46] . The dried plant parts (500 mg) were immersed overnight in 15 mL concentrated HNO3 in a conical flask. On the following day, 2 mL HClO4 was added and the sample was digested on a hot plate at 60˚C for 2 h, followed by further digestion at 90˚C until white fumes of HClO4 effervesced out. The leftover liquid was transferred to a 50 mL volumetric flask and diluted with doubly distilled water. The solution was filtered through Whatman paper No 41 filter paper and the total Fe content was determined by Atomic Absorption Spectrometry [47] .
2.8. Statistical Analysis
Data obtained were subjected to analysis of variance (ANOVA) with regard to the IRB populations’ development and activity, drainage and fertilization modes and rice yield using XLSTAT 2015 software. Mean variables were compared using the Fishers’ test at probability level p = 5% [39] .
3. Results and Discussion
3.1. Iron Reducing Bacterial Populations’ Dynamics in Soil near Rice Roots
The variance of the numbers of IRB in soil near rice roots, in relation to subsurface drainage and combined application of subsurface drainage and fertilization is presented in Table 1. The Fishers’ test revealed that the number of bacteria in the soil near BOU- AKE-189 and ROK-5 rice roots wasn’t significantly related to the subsurface drainage (p = 0.891 and p = 0.941, respectively). These results can be explained by the survival of IRB in drainage condition [18] . Thus, Ouattara [12] , Jacq et al. [48] and Liesack et al. [49] reported that som IRB can survive and grow in aerobic compartments in the presence of low pressure of oxygen where a surplus of oxygen is released by healthy roots. The IRB number was significantly related to the combined both factors (p = 0.050 and p = 0.020, Table 1). From the experiment, the D2 drainage in the NPK + Urea + Ca +

Table 1. Variance of IRB number in soil near rice roots in relation to subsurface drainage and fertilization, during the cultural cycle of BOUAKE-189 and ROK-5 rice varieties.
df = degree of freedom; F = Fisher F; *significant p < 0.05; **significant p < 0.01; ns: not significant p> 0.05.
Zn + Mg amended pots showed the lowest average number of IRB population in soil near rice roots, during the cultural cycle of both rice varieties (2 ×107 and 2.6 ×107 cells/g dry soil, for BOUAKE-189 and ROK-5 rice variety, respectively).
The present results are in agreement with those obtained in our previous study on Kamboinse paddy soils [18] , which showed that subsurface drainage combined to mineral amendment (D2 + NPK + Urea) reduced significantly the number of IRB in microplots (p = 0.050 and p = 0.020, respectively for the two rice varieties). According to our previous study on the same paddy soil the mean density of IRB population in the soil decreased in the NPK + Urea + Ca + Zn + Mg amended pots for BOUAKE-189 and ROK-5 rice varieties, respectively [39] . Furthermore, Benckiser et al. [36] and Jacq et al. [48] reported that the number of IRB decreased with increased supply of K, Ca, and Mg for IR22 and IR42 rice varieties (susceptible and tolerant to iron toxicity, respectively). Trolldenier [50] showed also that a sufficient mineral nutrition of potassium was important in maintaining the oxidising power of rice roots and in the reducing of IRB populations in rice fields.
Therefore, combined application of subsurface drainage and fertilization can lead to a significant reduction of IRB number in rice field.
The experiment showed also that the number of IRB in the soil near rice roots increased after two weeks of flooding in all pots for BOUAKE-189 and ROK-5 rice varieties (Figure 4, Figure 5). Soon as a soil is flooded or submerged by stagnant water, the reductive processes start [51] . Ethan and Odunze [6] reported that flooding affects electrochemical and chemical processes which in turn, affect soil fertility in a dynamic manner [12] . Thus, the growth of Iron Reducing Bacteria is stimulated by flooding [18] [39] [48] [52] .
It appeared also, that the number of IRB in soil near rice roots increased gradually with fluctuations from transplanting day to rice flowering and maturity stages in all the paddy pots (Figure 4, Figure 5). The highest densities of IRB, in most pots, were recorded from rice tillering and flowering to maturity stages (108 to 1010 cells/g dry soil). Berthelin et al. [48] observed a same evolution of IRB population during rice cultural cycle in a Senegal paddy soil. Our previous results obtained on Kamboinse and Kou Valley paddy soils [18] [39] reported the same pattern. Indeed, Dobermann and Fairhurst [30] reported that the periods of intense metabolic activity of rice plant (e.g., tillering, flowering and maturity), result in an increase of rhizoflora population, which in turn leads to an increased demand for electron acceptors. Thus, the highest level of reduced soil condition corresponds at these stages of growth at which rice plant enhance the exudation of carbohydrates and other metabolites sustaining IRB population growth in soil [18] [21] [48] . These results indicate that in spite of drainage and fertilization mode, rice plant remains the main factor that modulates the dynamic of IRB during rice cultural cycle, as reported by Jacq et al. [48] .
3.2. IRB Activity in Soil near Rice Roots
The variance of ferrous iron content in soil near rice roots in relation to subsurface drainage and combined application of subsurface drainage and fertilization for
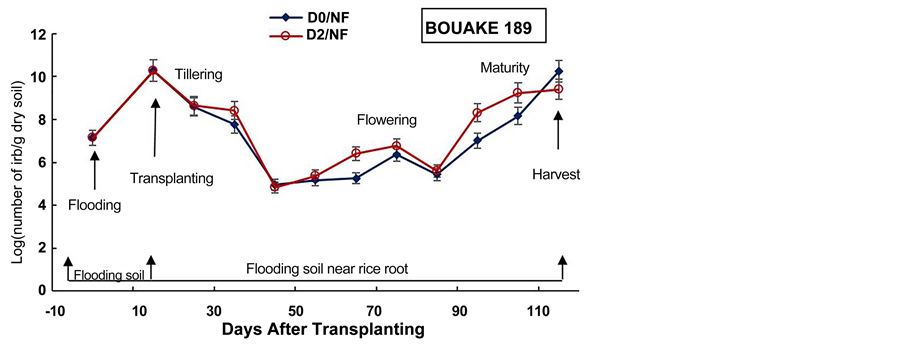
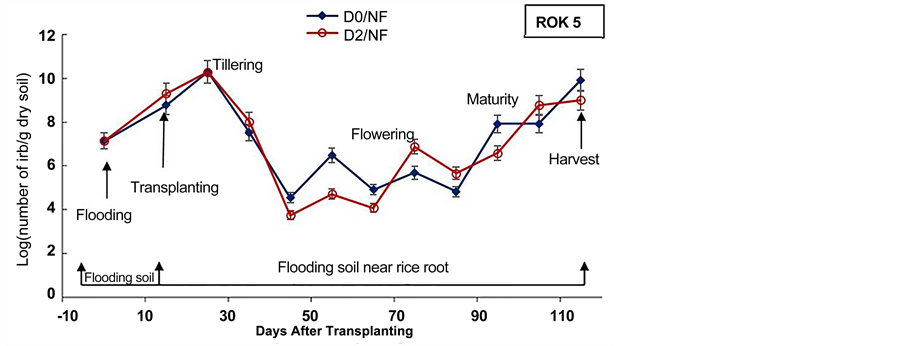
Figure 4. Densities of Iron Reducing Bacteria in soil before flooding, at trans- planting day and in soil near rice roots during the cultural cycle of BOUAKE-189 and ROK-5 rice varieties in pots without drainage and fertilization (D0/NF) and drained for 14 days (D2/NF), respectively (means of 3 replicates).
BOUAKE-189 and ROK-5 rice varieties are reported in Table 2. From the study, no significant effect of subsurface drainage and combined application of both factors was observed on the ferrous iron content in soil near rice roots, for BOUAKE-189 rice variety (p = 0.676 and p = 0.940, respectively), and for ROK-5 rice variety (p = 0.746 and p = 0.750, respectively), (Table 2).
From the present experiment, and in agreement with our previous study [18] , the application of subsurface drainage alone, doesn’t reduce significantly ferrous iron content in soil near rice roots for both varieties (Figure 6). These results can be explained by the production of ferrous iron in the rhizosphere, by surviving facultative anaerobic or aerobic Iron Reducing Bacteria [48] . As reported by Otoidobiga et al. [18] , ferrous iron production in drained microplots could be also ascribed by chemical reduction of Iron III in the soil [53] . The heterogeneous distribution of oxygen through the drainage can also create anoxic compartments where ferrous iron can be reduced by surviving anaerobic IRB population [26] [49] .
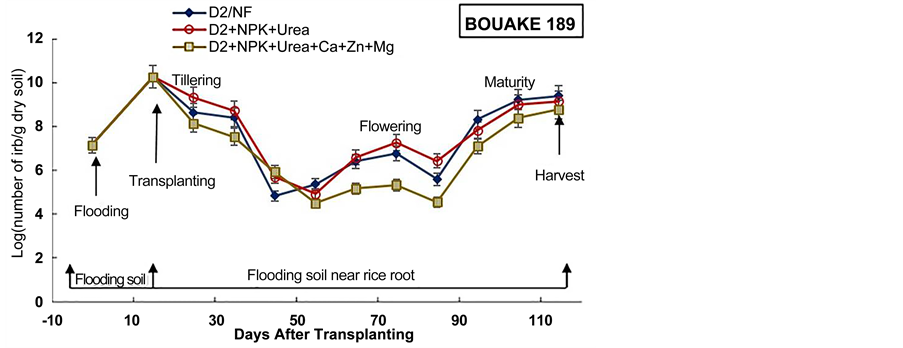
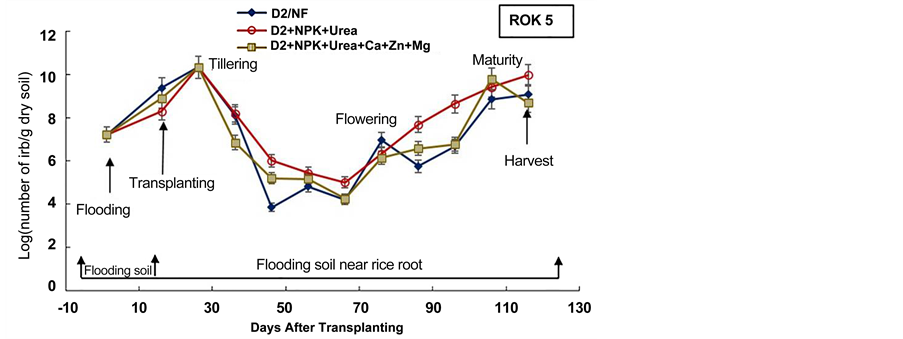
Figure 5. Densities of Iron Reducing Bacteria in soil before flooding, at transplanting day and in soil near rice roots during the cultural cycle of BOUAKE-189 and ROK-5 rice varieties in drained pots for 14 days without fertilization (D2/NF) and combined application of D2 drainage, NPK + Urea and NPK + Urea + Ca + Zn + Mg fertilization (D2 + NPK + Urea and D2 + NPK + Urea + Ca + Zn + Mg), respectively (means of 3 replicates).
Table 2. Variance of ferrous iron content in soil near rice roots in relation to subsurface drainage and fertilization, during the cultural cycle of BOUAKE-189 and ROK-5 rice varieties.
df = degree of freedom; F = Fisher F; *significant p < 0.05; **significant p < 0.01; ns: not significant p > 0.05.
The average of the ferrous iron content in the soil near rice roots was low in D2 + NPK + Urea amended pots, relatively to the control pots, for the two rice varieties (Figure 7). Indeed, Trolldenier [50] , revealed that the nutritional status of rice plant influences bacterial activity and the oxidation-reduction conditions around the roots. Moreover, Jacq et al. [48] mentioned that, as long as P and K uptake by roots remains effective and roots are not damaged, the oxygen flow from aerial parts of the rice plant is sufficient to oxidize small amounts of Fe (II), and Fe (III)-oxides may precipitate on the root-soil interface (rhizoplane). Thus, we can deduct that the D2 subsurface drainage combined to NPK + Urea amendment may contribute to the decrement of iron reduction in rice paddy soil.
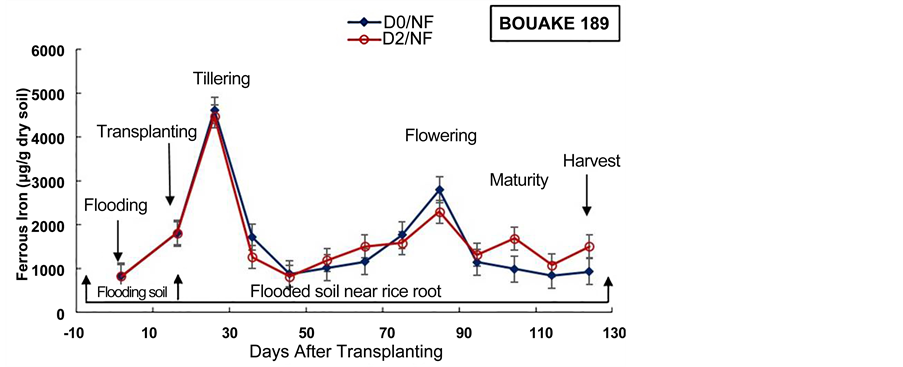
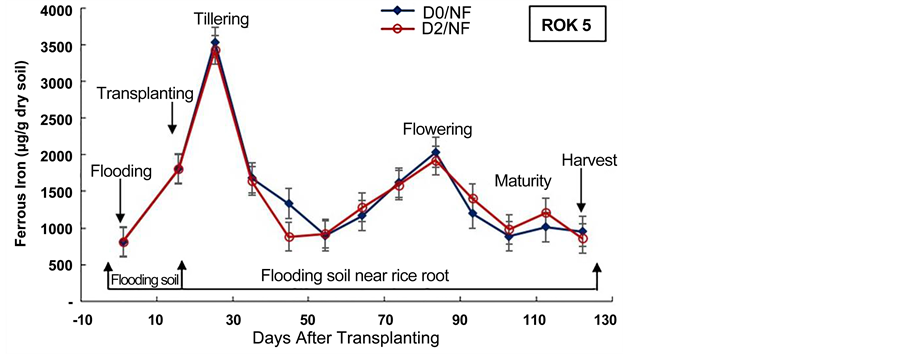
Figure 6. Evolution of soil ferrous iron content during the cultural cycle of BOU- AKE-189 and ROK-5 rice varieties in pots without drainage (D0) and drained for 14 days (D2), respectively (means of 3 replicates).
Furthermore, the experiment evidenced the highest ferrous iron content in the soil of D2 + NPK + Urea + Ca + Zn + Mg pots (Figure 7), for the two rice varieties. Indeed, the role of Ca, Mg, and Zn fertilizers is the regulation of ferrous iron absorption in the rice plant, both as competing ion and by increasing the plant tolerance to iron toxicity [54] [55] . Therefore, in the present experiment, D2 + NPK + Urea + Ca + Zn + Mg application doesn’t reduce ferrous iron production in rice fields, however, NPK + Urea + Ca + Zn + Mg application permits to support rice plant tolerance to the high content of toxic iron in soil as previous underlined by Otoidobiga et al. [39] .
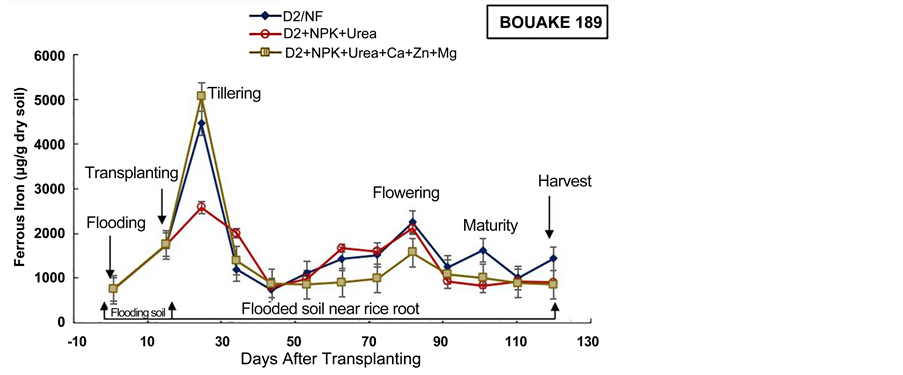
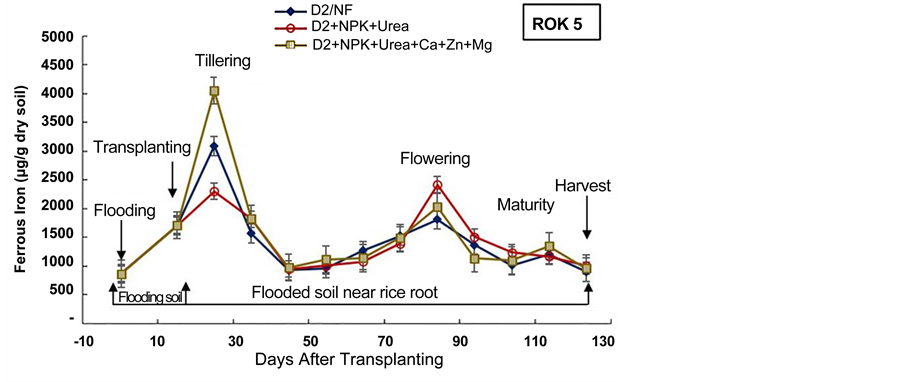
Figure 7. Evolution of soil ferrous iron content during the cultural cycle of BOUAKE-189 and ROK-5 rice varieties in drained pots for 14 days without fertilization (D2/NF), and combined application of D2 drainage NPK+Urea and NPK + Urea + Ca + Zn + Mg fertilization (D2 + NPK + Urea, and D2 + NPK + Urea + Ca + Zn + Mg), respectively (means of 3 replicates).
The concentration of ferrous iron increased after two weeks of soil flooding for all the treatments (Figure 6, Figure 7). This result is in agreement with our previous results obtained in Kamboinse paddy soils [18] . Jacq et al. [48] and Betremieux [52] reported also the same increment of ferrous iron content in soil after the flooding of a Senegal paddy soil. Indeed, Becker and Asch [10] and Shahid et al. [56] , underlined that iron toxicity occurs only in flooded soils and affects primarily the production of lowland rice. Thus, the reducing conditions of waterlogged lowland soils boost iron toxicity through solubilization of almost all iron into its ferrous form (Fe2+) [10] [56] . Moreover, Jacq et al. [48] indicated also that a major part of rice crop losses in southern Senegal, were ascribed to primary iron toxicity because of transplanting of seedlings immediately after flooding by most farmers.
In most pots, the highest content of ferrous iron in soil near rice roots was recorded from rice tillering and flowering to maturity stages (103 to 5 × 103 µg/g dry soil) (Figure 6, Figure 7). These results can be explained by enhanced iron microbial reduction in the rizosphere due to intensive exudation during the physiological active phase between heading and flowering [18] [21] [48] [57] . These results are in agreement with those obtained by Dobermann and Fairhurst [30] which showed that in these periods of intense metabolic activity, facultative and obligate anaerobic bacteria reduce Fe3+ to Fe2+. In fact, Ethan and Odunze [6] recorded that in reductive condition, the number of Iron Reducing Bacteria in soil increased almost parallel to the decrease in redox potential and to the increase in iron reducing power [49] [50] [58] . Jacq et al. [48] also revealed that despite the aeration mechanism of the roots, iron-reduction pro- cesses in the bulk soil are stimulated by the physiological activity and by the growth of rice roots. Moreover, many studies revealed also that redox potential was higher in soils near the plants than in soils away from the plants [50] [55] . These results highlighted that the nutritional status of the rice plant essentially influences the activity of bacteria and the oxidation-reduction conditions around the roots. Furthermore, Trolldenier [50] and Prade et al. [57] reported that root debris and/or exudation of carbohydrates at these stages of rice growth can decrease redox potential and stimulate microbial activity and anaerobic respiration (denitrification and/or ferric iron reduction).
3.3. Effect of Combined Application Subsurface Drainage and Fertilization on BOUAKE-189 and ROK-5 Rice Plants Total Iron Content
The effects of drainage and fertilization on iron content in BOUAKE-189 and ROK-5 rice plants were recorded during the study (Table 3, Table 4). The analysis of variance
Table 3. Variance of total iron content in rice roots in relation to subsurface drainage and ferti- lization, during the cultural cycle of BOUAKE-189 and ROK-5 rice varieties.
df = degree of freedom; F = Fisher F; *significant p < 0.05; **significant p < 0.01; ns: not significant p > 0.05.
Table 4. Variance of total iron content in rice plant in relation to subsurface drainage and fer- tilization, during the cultural cycle of BOUAKE-189 and ROK-5 rice varieties.
df = degree of freedom; F = Fisher F; *significant p < 0.05; **significant p < 0.01; ns: not significant p > 0.05.
showed that the total iron content in the roots of both rice varieties wasn’t significantly related to subsurface drainage (p = 0.075 and p = 0.182, respectively) and combined application of fertilization and drainage (p = 0.092 and p = 0.191, respectively) (Table 3). It appeared also that the drained pots (D2, D2 + NPK + Urea and D2 + NPK + Urea + Ca + Zn + Mg) showed the highest total iron contents in roots (Figure 8). These results are in agreement with those of Mullilab [59] and Ethan and Odunze [6] who reported that the drainage of a waterlogged soil oxidizes the soil and precipitates Fe2+ into Fe3+ forms which are insoluble at the root surface.
The total iron accumulation in BOUAKE-189 and ROK-5 rice aerial biomass was significantly related to subsurface drainage (p = 0.011, Table 4). However, no significant effect of combined application of both factors was observed for the two rice varieties (p = 0.056, Table 4).
Moreover, the total iron contents of aerial biomass in the drained (D2) pots were significantly high comparatively to the controls (D0) pots (p = 0.011) (Figure 9). Panda et al. [60] , explaining the physiological and biochemical mechanisms of Fe uptake by rice plant indicated that rice plants release into the rhizosphere siderophores which bind to Fe3+ in the form of a ligand. The ligand complex enters into the cell, and Fe3+ is reduced into Fe2+ inside the cytoplasm. Therefore, drainage may optimize iron uptake by rice plant by increasing the availability of Fe3+ through the oxidation of ferrous iron.
The experiment revealed also that the total iron content in aerial biomass of pots which received combined application of subsurface drainage and fertilization (D2 +
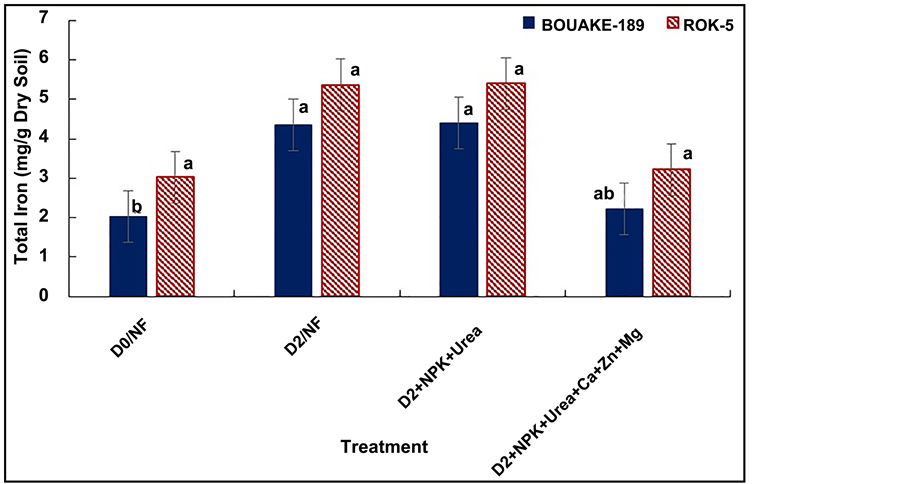
Figure 8. Roots total iron content of BOUAKE-189 and ROK-5 rice varieties in drained for 14 days without fertilization (D2/NF) and combined application of D2 drainage, NPK+Urea and NPK + Urea + Ca + Zn + Mg fertilization (D2 + NPK + Urea and D2 + NPK + Urea + Ca + Zn + Mg), respectively (means of 3 replicates). Yields sharing the same letter are not significantly different according to Fishers’ test p > 0.05.
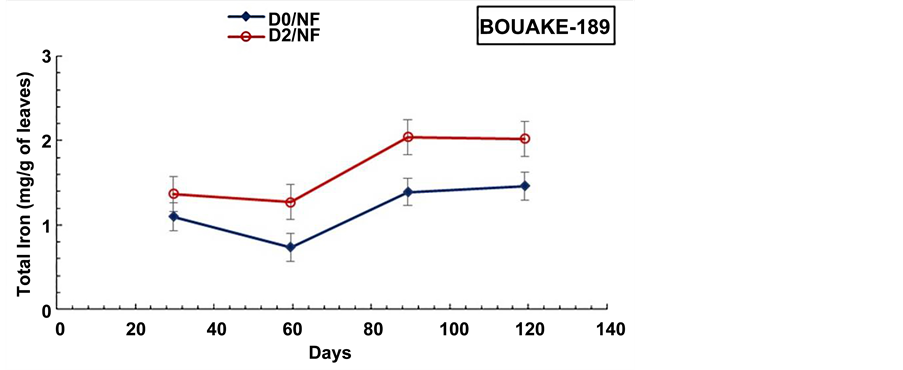
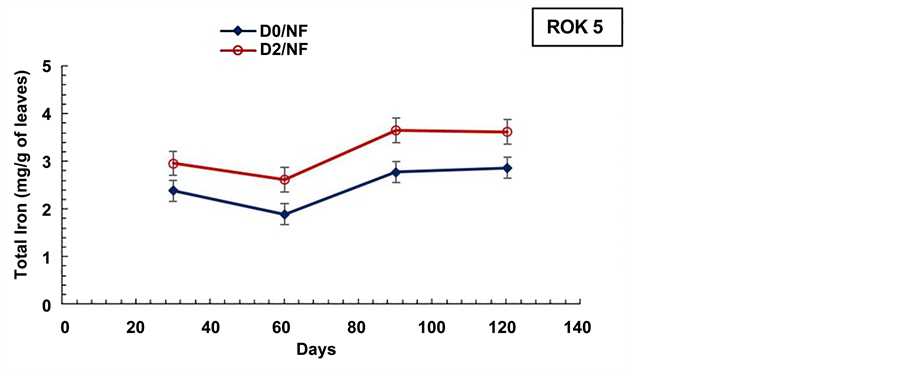
Figure 9. Evolution of the aerial biomass total iron content during the cultural cycle of BOUAKE-189 and ROK-5 rice varieties in pots without drainage and fertilization (D0/NF) and in pots drained for 14 days without fertilization (D2/ NF) (means of 3 replicates).
NPK + Urea and D2 + NPK + Urea + Ca + Zn + Mg) were higher relatively to D2 drained and non-drained pots (D0) (Figure 9, Figure 10), for both rice varieties. The combined application of subsurface drainage and mineral fertilization seems to have a synergic effect on the rice plant Fe3+ absorption. Thus, in combined application of subsurface drainage and mineral fertilization condition, the drainage favours oxidation and precipitation of Fe2+ into Fe3+ forms, while the mineral fertilization (NPK + Urea + Ca + Zn + Mg) contributes to Fe3+ acquisition by enabling the plant to synthesize more photosynthetic assimilates (NADPH+, H+) [6] [59] .
3.4. Effect of Combined Application of Subsurface Drainage and Fertilization on BOUAKE-189 and ROK-5 Rice Varieties Yield
Throughout the study, a significant difference was observed on rice biomass yield for subsurface drainage and combined effects of both factors (p = 0.001 and p = 0.000, respectively) for ROK-5 rice varieties (Table 5). Furthermore, the experiment revealed that the biomass yield of BOUAKE-189 rice variety was related to subsurface drainage (p = 0.007, Table 5), while combined effects of both factors showed no significant difference (p = 0.108, Table 5). However, it appeared that D2 + NPK + Urea + Ca + Zn + Mg treatments showed the highest rice biomass yield, followed by D2 + NPK + Urea ones, for the two rice varieties, respectively (Figure 11). Becker and Asch [10] indicated that various water, crop, and nutrient management options could alleviate the negative effects of Fe toxicity on lowland-rice performance. The findings of our studies are supported by the results of Ethan and Odunze [6] , who recorded an increase of rice yield during a study on an iron toxic soil amended with nitrogen fertilizers and subjected to periodical drainage. Our previous study in Kamboinse reported also that microplots, which received D2 subsurface drainage and NPK + Urea amendment showed the highest yield [18] .
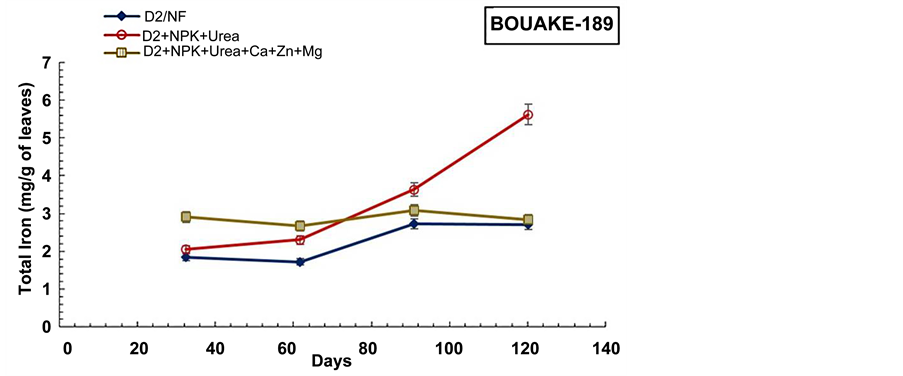
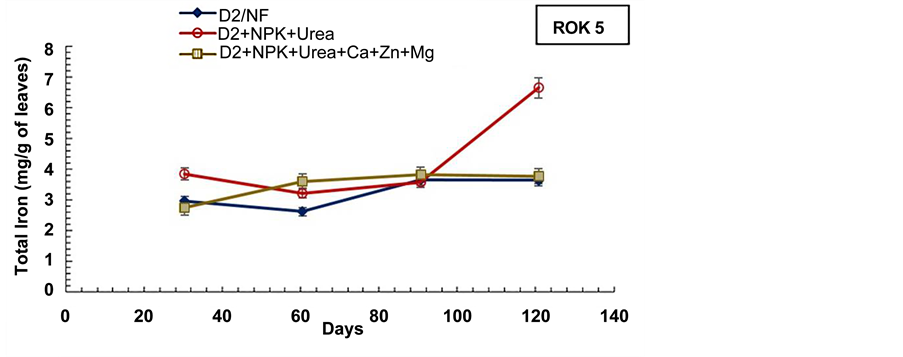
Figure 10. Evolution of the aerial biomass total iron content during the cultural cycle of BOUAKE-189 and ROK-5 rice varieties in drained pots for 14 days without fertilization (D2/NF), and in pots with combined application of D2 drainage NPK + Urea and NPK + Urea + Ca + Zn + Mg fertilization (D2 + NPK + Urea, and D2 + NPK + Urea + Ca + Zn + Mg), respectively (means of 3 replicates).
Table 5. Variance of total biomass yield in relation to subsurface drainage and fertilization, for BOUAKE-189 and ROK-5 rice varieties.
df = degree of freedom; F = Fisher F; *significant p < 0.05; **significant p < 0.01; ns: not significant p > 0.05.
Figure 11. Total biomass yield of BOUAKE-189 and ROK-5 rice varieties in drained (D2) in drained pots for 14 days without fertilization (D2/NF) and combined application of D2 drainage, NPK + Urea and NPK + Urea + Ca + Zn + Mg fertilization (D2 + NPK + Urea and D2 + NPK + Urea + Ca + Zn + Mg), respectively (means of 3 replicates). Yields sharing the same letter are not significantly different according to Fishers’ test p > 0.05.
Keita [61] and Ethan et al. [6] indicated that drainage in iron-toxic soils increased grain yield of lowland rice by the reduction of ferrous iron absorption of rice plant. Indeed, many studies showed that soil oxygenation through water drainage affects the iron profiles by inducing fast chemical and microbial oxidization reactions of the iron II in the zones where oxygen is present [49] [62] .
Our previous experiment, on the effect of fertilization on the rice yield in the same paddy soil, reported also that the pots which received NPK + Urea + Ca + Zn + Mg amendment, recorded the highest yield, relatively to NPK + Urea fertilization [39] . In fact, the application of plant essential nutrients counteracts negative effects of excess amounts of iron, by competing with Fe2+ uptake at the sites of ion adsorption on roots or by enhancing plants’ defence or tolerance mechanisms [10] [55] [63] .
However, the D2 + NPK + Urea + Ca + Zn + Mg treatment showed the highest yield (296.6 and 433.7 g/pot, respectively) relatively to solely application of NPK + Urea + Ca + Zn + Mg (250.017 and 362.87 g/pot, respectively), as previously reported in the same paddy soil, for BOUAKE-189 and ROK-5 rice varieties, respectively [39] . Thus, in the present study, the oxidation power of the subsurface drainage [6] combined to the mineral (Ca, Mg, Mn P, K and Zn) fertilization may optimize the acquisition of Fe by rice plant for an improved growth and yield [55] [63] [64] .
4. Conclusions
From the results obtained in this study, subsurface drainage combined with NPK + Urea + Ca + Zn + Mg fertilization decreases the number of IRB in an iron toxic paddy soil under BOUAKE-189 and ROK-5 rice varieties cultivation. Thus, oxygenation throughout drainage and mineral fertilization of the soil maintains the oxidising power of rice roots and reduces the IRB population’s number. Moreover, the low level of ferrous iron content in the soil near rice roots, for both rice varieties in the D2 + NPK + Urea pots, indicates that the combination of drainage and NPK + Urea fertilization may decrease iron reduction and/or optimize the absorption of Fe(III) complexes into the rice plant. However, the subsurface drainage combined to NPK + Urea + Ca + Zn + Mg complex amendment recorded high level of ferrous iron content in the soil near rice roots, high content of total iron in the aerial biomass and the highest mean of rice biomass yield for both rice varieties. Indeed, combined application of drainage and mineral fertilization seems to have a synergic effect on improving rice yield. The oxidation power of the subsurface drainage combined to the mineral (Ca, Mg, Mn P, K and Zn) fertilization may optimize the acquisition of Fe by rice plant for an improved growth and yield. In fact, for the two rice varieties, NPK, Ca, Mg, and Zn amendment seems to increase the leaf tissue tolerance to excess amounts of Fe and to optimize Fe3+ absorption, produced through the oxidation of ferrous iron during the subsurface drainage, at the root surface in the rice plant, for a better growth and better yield.
The experiment showed also that the rice plant remains the main factor which modulates the dynamic and activity of IRB during rice cultural cycle.
Acknowledgements
The authors would like to express profound gratitude to International Foundation for Science, CNS-FL/WAAP, FCN/WAAPP, CIOSPB, PACER-UEMOA/RABIOTECH, CNR- ST/IRSS, General Direction of Meteorology-Burkina Faso and CRSBAN-DBM/UFR- SVT/U. Ouaga I Pr. J. KI-ZERBO for financial and technical supports.
Cite this paper
Otoidobiga, C.H., Kam, H., Bagayogo, A., Savadogo, A., Sawadogo, J.B., Sawadogo, S., Sawadogo, A., Sinaré, Y., Ouédraogo, I., Zombré, P., Asakawa, S., Traoré, A.S. and Dianou, D. (2016) Effect of Combined Application of Subsurface Drainage and Mineral Fertilization on Iron-Reducing Bacterial Populations’ Developments and Fe2+ Uptake by Two Rice Varieties in an Iron Toxic Paddy Soil of Burkina Faso (West Africa). Agricultural Sciences, 7, 783-804. http://dx.doi.org/10.4236/as.2016.711072
References
- 1. INERA (2000) Descriptive Folders of Rice Cultivars. Institute of Environment and Agricultural Research, Ouagadougou.
- 2. MAHRH (2009) Evolution of Rice Areas, Yields and Productivities in Burkina Faso 1984-2009. Ministry of Agriculture, Water and Fisheries Resources, Ouagadougou.
- 3. Sikirou, M., Saito, K., Achigan-Dako, E.G., Dramé, K.N., Ahanchédé, A. and Venuprasad, R. (2015) Genetic Improvement of Iron Toxicity Tolerance in Rice-Progress, Challenges and Prospects in West Africa. Plant Production Science, 18, 423-434.
http://dx.doi.org/10.1626/pps.18.423 - 4. Brady, N.C. (1982) Rice Research Strategies for the Future. The International Rice Research Institute, Manila, 13-25.
- 5. Chérif, M., Audebert, A., Fofana, M. and Zouzou, M. (2009) Evaluation of Iron Toxicity on Lowland Irrigated Rice in West Africa. Tropicultura, 27, 88-92.
- 6. Ethan, S. and Odunze, S. (2011) Effect of Water Management and Nitrogen Rates on Iron Concentration and Yield in Lowland Rice. Agriculture and Biology Journal of North America, 2, 622-629.
http://dx.doi.org/10.5251/abjna.2011.2.4.622.629 - 7. Olaleye, A.O., Tabi, F.O., Ogunkunle, A.O., Singh, B.N. and Sahrawat, K.L. (2001) Effect of Toxic Iron Concentrations on the Growth of Lowlands Rice. Journal of Plant Nutrition, 24, 441-457.
http://dx.doi.org/10.1081/PLN-100104971 - 8. Wan, J.L., Zhai, H.Q. and Wan, J.M. (2005) Mapping of QTLs for Ferrous Iron Toxicity Tolerance in Rice (Oryza sativa L.). Journal of Genetics and Genomics, 32, 1156-1166.
- 9. Gridley, H.E., Efisue, A., Tolou, B. and Bakayako, T. (2006) Breeding for Tolerance to Iron Toxicity at WARDA. In: Audebert, A., Narteh, L.T., Kiepe, P., Millar, D. and Beks, B., Eds., Iron Toxicity in Rice-Based System in West Africa, WARDA, Cotonou, 96-111.
- 10. Becker, M. and Asch, F. (2005) Iron Toxicity in Rice—Conditions and Management Concepts. Journal of Plant Nutrition and Soil Science, 168, 558-573.
http://dx.doi.org/10.1002/jpln.200520504 - 11. Haefele, S.M., Nelson, A. and Hijmans, R.J. (2014) Soil Quality and Constraints in Global Rice Production. Geoderma, 235-236, 250-259.
http://dx.doi.org/10.1016/j.geoderma.2014.07.019 - 12. Ouattara, A.S. (1992) Contribution to the Study of Iron Reducing Bacteria and Sulfate in Paddy Soils of the Kou Valley (Burkina Faso). PhD Dissertation, University of Provence, Aix-Marseille.
- 13. Wellens, J. and Nitcheu, M.N. (2010) Appropriation and Popularization of Tools of Decision Support in the Co-Management of the Perimeter Irrigated of the Kou Valley (Southwest of Burkina Faso).
http://www.ge-eau.org/Documents/SIMIS_etude%20de%20cas.pdf - 14. Keita, A., Yacouba, H., Hayde, L.G. and Schultz, B. (2013) A Single-Season Irrigated Rice Soil Presents Higher Iron Toxicity Risk in Tropical Savannah Valley Bottoms. Open Journal of Soil Science, 3, 314-322.
http://dx.doi.org/10.4236/ojss.2013.37036 - 15. Quinet, M., Vromman, D., Clippe, A., Bertin, P., Lequeux, H., Dufey, I., Lutts, S. and Lefèvre, I. (2012) Combined Transcriptomic and Physiological Approaches Reveal Strong Differences between Short- and Long-Term Response of Rice (Oryza sativa) to Iron Toxicity. Plant, Cell and Environment, 35, 1837-1859.
http://dx.doi.org/10.1111/j.1365-3040.2012.02521.x - 16. Stein, R.J., Duarte, G.L., Spohr, M.G., Lopes, S.I.G. and Fett, J.P. (2009) Distinct Physiological Responses of Two Rice Cultivars Subjected to Iron Toxicity under Field Conditions. Annals of Applied Biology, 154, 269-277.
http://dx.doi.org/10.1111/j.1744-7348.2008.00293.x - 17. Briat, J.F. (1997) Iron Transport and Storage in Plants. Trends in Plant Science, 2, 187-193.
http://dx.doi.org/10.1016/S1360-1385(97)85225-9 - 18. Otoidobiga, C.H., Keita, A., Yacouba, H., Traore, A.S. and Dianou, D. (2015) Dynamics and Activity of Iron-Reducing Bacterial Populations in a West African Rice Paddy Soil under Subsurface Drainage: Case Study of Kamboinse in Burkina Faso. Agricultural Sciences, 6, 860-869.
http://dx.doi.org/10.4236/as.2015.68083 - 19. Ehrlich, H.L. and Newman, D.K. (2009) Geomicrobiology. CRC Press, Boca Raton, 606 p.
- 20. Luef, B., Fakra, S.C., Csencsits, R., Wrighton, K.C., Williams, K.H., Wilkins, M.J., Downing, K.H., Long, P.E., Comolli, L.R. and Banfield, J.F. (2013) Iron Reducing Bacteria Accumulate Ferric Oxyhydroxide Nanoparticle Aggregates That May Support Planktonic Growth. International Society for Microbial Ecology Journal, 7, 338-350.
- 21. Yi, W., Wang, B. and Qu, D. (2012) Diversity of Isolates Performing Fe(III) Reduction from Paddy Soil Fed by Different Organic Carbon Sources. African Journal of Biotechnology, 11, 4407-4417.
- 22. Hori, T., Aoyagi, T., Itoh, H., Narihiro, T., Oikawa, A., Suzuki, K., Ogata, A., Friedrich, M.W., Conrad, R. and Kamagata, Y. (2015) Isolation of Microorganisms Involved in Reduction of Crystalline Iron(III) Oxides in Natural Environments. Frontiers in Microbiology, 6, 386.
http://dx.doi.org/10.3389/fmicb.2015.00386 - 23. Borch, T., Kretzschmar, R., Kappler, A., Van Cappelen, P., Ginder-Vogel, M., Voegelin, A. and Campbell, K. (2010) Biogeochemical Redox Processes and Their Impact on Contaminant Dynamics. Environment Science Technology, 44, 15-23.
http://dx.doi.org/10.1021/es9026248 - 24. Lalonde, K., Mucci, A., Ouellet, A. and Gelinas, Y. (2012) Preservation of Organic Matter in Sediments Promoted by Iron. Nature, 483, 198-200.
http://dx.doi.org/10.1038/nature10855 - 25. Melton, E.D., Swanner, E.D., Behrens, S., Schmidt, C. and Kappler, A. (2014) The Interplay of Microbially Mediated and Abiotic Reactions in the Biogeo-Chemical Fe Cycle. Nature Reviews Microbiology, 12, 797-808.
http://dx.doi.org/10.1038/nrmicro3347 - 26. Ponnamperuma, F.N. (1972) The Chemistry of Submerged Soils. Advances in Agronomy, 24, 29-96.
http://dx.doi.org/10.1016/S0065-2113(08)60633-1 - 27. Yoshida, S. (1981) Fundamentals of Rice Crop Science. International Rice Research Institute, Manila.
- 28. Sahrawat, K.L. (2004) Iron Toxicity in Wetland Rice and the Role of Other Nutrients. Journal of Plant Nutrition, 27, 1471-1504.
http://dx.doi.org/10.1081/PLN-200025869 - 29. Ponnamperuma, F.N., Bradfield, R. and Peech, M. (1955) Physiological Disease of Rice attributable to Iron Toxicity. Nature, 175, 265-265.
http://dx.doi.org/10.1038/175265a0 - 30. Dobermann, A. and Fairhurst, T. (2000) Rice: Nutrient Disorders & Nutrient Management. International Rice Research Institute, Los Baños, 191.
- 31. Wu, L., Shhadi, M.Y., Gregorio, G., Matthus, E., Becker, M. and Frei, M. (2014) Genetic and Physiological Analysis of Tolerance to Acute Iron Toxicity in Rice. The Rice Journal, 7, 8.
http://dx.doi.org/10.1186/s12284-014-0008-3 - 32. Mengel, K. and Kirkby, E.A. (1987) Principles of Plant Nutrition. 4th Edition, International Potash Institute, Bern.
- 33. Bienfait, H.F., Van Den Briel, M.L. and Mesland-Mul, N.T. (1985) Free Space Iron Pools in Roots: Generation and Mobilization. Plant Physiology, 78, 596-600.
http://dx.doi.org/10.1104/pp.78.3.596 - 34. Vahl, L.C. (1991) Iron Toxicity in Rice Genotypes Irrigated by Flooding. PhD Thesis, Federal University of Rio Grande do Sul, Porto Alegre.
- 35. Schmidt, W. (1999) Review Mechanisms and Regulation of Reduction-Based Iron Uptake in Plants. New Phytologist, 141, 1-26.
http://dx.doi.org/10.1046/j.1469-8137.1999.00331.x - 36. Benckiser, G., Ottow, J.C.G., Watanabe, I. and Santiago, S. (1984) The Mechanism of Excessive Iron-Uptake (Iron Toxicity) of Wetland Rice. Journal of Plant Nutrition, 7, 177-185.
http://dx.doi.org/10.1080/01904168409363184 - 37. Kosaki, T. and Juo, A.S.R. (1986) Iron Toxicity in Inland Valleys: A Case Study from Nigeria. In: Juo, A.S.R. and Lowe, J.A., Eds., Wetlands and Rice in Sub-Saharan Africa Ibadan, IITA, Ibadan, 167-174.
- 38. Audebert, A. and Sahrawat, K.L. (2000) Mechanisms for Iron Toxicity Tolerance in Lowland Rice. Journal of Plant Nutrition, 23, 1877-1885.
http://dx.doi.org/10.1080/01904160009382150 - 39. Otoidobiga, C.H., Sawadogo, A., Sinaré, Y., Ouédraogo, I., Zombré, P., Asakawa, S., Traore, A.S. and Dianou, D. (2016) Effect of Fertilization on the Dynamics and Activity of Iron Reducing Bacterial Populations in a West African Rice Paddy Soil Planted with Two Rice Varieties: Case Study of Kou Valley in Burkina Faso. Journal of Environmental Protection, 7, 1119-1131.
http://dx.doi.org/10.4236/jep.2016.78101 - 40. ISRA (2012) Official Catalogue of Species and Varieties Cultivated in Senegal. Agricultural Research Institute of Senegal. Ministry of Agriculture and Rural Equipment, Dakar, Senegal, 212 p.
- 41. Singh, B.P., Das, M., Prasad, R.N. and Ram, M. (1992) Characteristics of Fe-Toxic Soils and Affected Plants and Their Correction in Acid Haplaquents of Meghalaya. International Rice Research Newsletter, 17, 18-19.
- 42. Sokona, M.E.B., Boro, A., Hema, A. and Katiella, B. (2010) Diagnostic Study of the Rice Irrigation Scheme of Tiefora, Province of Comoe, Region of the Cascades. Field Report, 2iE, Ouagadougou.
- 43. Jutras, G. (2006) Guide of Interpretation of Soil Analysis.
http://blog.ac-versailles.fr/formationcapa/public/MP2/Interpretation_AnalyseSol_.pdf - 44. Hammann, R. and Ottow, J.C.G. (1974) Reductive Dissolution of Fe2O3 by Saccharolytic Clostridia and Bacillus Poly-Myxa under Anaerobic Conditions. Journal of Plant Nutrition and Soil Science, 137, 108-115.
- 45. Vizier, J.F. (1969) Selection and Development of a Method for Iron Ferrous Dosage in Waterlogged Soils. ORSTOM Books, Pedology Serial, Montpellier.
- 46. Chapman, H.D. and Pratt, P.F. (1961) Methods of Analysis for Soils, Plants and Waters. University of California, Berkeley.
- 47. Murphy, J. and Riley, J.P. (1962) A Modified Single Solution Method for Determination of Phosphorus in Natural Waters. Analytica Chimica Acta, 27, 31-36.
http://dx.doi.org/10.1016/S0003-2670(00)88444-5 - 48. Jacq, V.A., Prade, K. and Ottow, J.G. (1991) Iron Sulphide Accumulation in the Rhizosphere of Wetland Rice (Oriza sativa L.) as the Result of Microbial Activities. In: Fyfe, W.S., Ed., Developments in Geochemistry, Vol. 6, Diversity of Environmental Biogeochemistry, Elsevier, Amsterdam, 453-468.
- 49. Liesack, W., Schnell, S. and Revsbech, N.P. (2000) Microbiology of Flooded Rice Paddies. FEMS Microbiology Reviews, 24, 625-645.
http://dx.doi.org/10.1111/j.1574-6976.2000.tb00563.x - 50. Trolldenier, G. (1977) Mineral Nutrition and Reduction Processes in the Rhizosphere of Rice. Plant and Soil, 47, 193-202.
http://dx.doi.org/10.1007/BF00010379 - 51. Conrad, R. (2007) Microbial Ecology of Methanogens and Methanotrophs. Advances in Agronomy, 96, 1-63.
http://dx.doi.org/10.1016/S0065-2113(07)96005-8 - 52. Betremieux, R. (1951) Experimental Study of Iron and Manganese in the Soils. Annales Agronomiques, 10, 193-295.
- 53. Ehrlich, H.L. (1996) Geomicrobiology. 3rd Edition, Marcel Dekker, New York.
- 54. Sahrawat, K.L. (2010) Reducing Iron Toxicity in Lowland Rice with Tolerant Genotypes and Plant Nutrition. Plant Stress, 4, 70–75.
- 55. Fageria, N.K., Santos, A.B., Barbosa Filho, M.P. and Guimarães, C.M. (2008) Iron Toxicity in Lowland Rice. Journal of Plant Nutrition, 31, 1676-1697.
http://dx.doi.org/10.1080/01904160802244902 - 56. Shahid, M., Nayak, K.A., Shukla, A.K., Tripathi, R., Kumar, A., Raja, R., Panda, B.B., Meher, J., Bhattacharyya, P. and Dash, D. (2014) Mitigation of Iron Toxicity and Iron, Zinc, and Manganese Nutrition of Wetland Rice Cultivars (Oryza sativa L.) Grown in Iron-Toxic Soil. Clean Soil, Air, Water, 42, 1604-1609.
http://dx.doi.org/10.1002/clen.201300178 - 57. Prade, K., Ottow, J.C.G. and Jacq, V.A. (1993) Excessive Iron Uptake (Iron Toxicity) by Wetland Rice (Oryza sativa L.) on an Acid Sulphate Soil in the Casamance/Senegal. International Institute for Land Reclamation and Improvement, Wageningen, the Netherlands. International Livestock Research Institute Publication, 44, 150-162.
- 58. Begg, C.B.M., Kirk, G.J.D., Mackenzie, A.F. and Neue, H.U. (1994) Root-Induced Iron Oxidation and pH Changes in Lowland Rice Rhizosphere. New Phytologist, 128, 469-477.
http://dx.doi.org/10.1111/j.1469-8137.1994.tb02993.x - 59. Mullilab, H. (2008) Manufacturers of Microbiology Products. New Delhi, India.
- 60. Panda, B.B., Sharma, S.G., Mohapatra, P.K. and Das, A. (2012) Iron Stress Induces Primary and Secondary Micronutrient Stresses in High Yielding Tropical Rice. Journal of Plant Nutrition, 35, 1359-1373.
http://dx.doi.org/10.1080/01904167.2012.684128 - 61. Keïta, A. (2015) Subsurface Drainage of Valley Bottom Irrigated Rice Schemes in Tropical Savannah: Case Studies of Tiefora and Moussodougou in Burkina Faso. PhD Thesis, Wageningen University, Delft.
- 62. Hauck, S., Benz, M., Brune, A. and Schink, B. (2001) Ferrous Iron Oxidation by Denitrifying Bacteria in Profundal Sediments of a Deep Lake (Lake Constance). FEMS Microbiology Ecology, 37, 127-134.
http://dx.doi.org/10.1111/j.1574-6941.2001.tb00860.x - 63. Panda, B.B., Sharma, S., Mohapatra, P.K. and Das, A. (2012). Application of Excess Nitrogen, Phosphorus, and Potassium Fertilizers Leads to Lowering of Grain Iron Content in High-Yielding Tropical Rice. Communications in Soil Science and Plant Analysis, 43, 2590-2602.
http://dx.doi.org/10.1080/00103624.2012.716122 - 64. Audebert, A. (2006) Diagnosis of Risk and Approaches to Iron Toxicity Management in Lowland Rice Farming. In: Audebert, A., Narteh, L.T., Kiepe, P., Millar, D. and Beks, B., Eds., Iron Toxicity in Rice-Based Systems in West Africa, ADRAO, Cotonou, 6-17.