Advances in Enzyme Research
Vol.04 No.01(2016), Article ID:64432,13 pages
10.4236/aer.2016.41002
Characterization of an Exopolygalacturonase from Leucoagaricus gongylophorus, the Symbiotic Fungus of Atta sexdens
Paulo R. Adalberto, Camilla C. Golfeto, Ariele C. Moreira, Fernando G. Almeida, Douglas Ferreira, Quezia B. Cass, Dulce H. F. Souza*
Chemistry Department, Federal University of Sao Carlos, São Carlos, Brazil

Copyright © 2016 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 5 February 2016; accepted 8 March 2016; published 11 March 2016
ABSTRACT
The present study aimed to purify and characterize one polygalacturonase from L. gongylophorus (PGaseLg), the symbiotic fungus of Atta sexdens. The enzyme was isolated by salting out of crude extract followed by two chromatographic steps. PGaseLG was identified with MS analysis and molecular exclusion chromatography revealed the monomeric nature of a protein with an estimated molecular weight of about 39 kDa. PGaseLg has an optimum temperature of 60˚C and optimum pH activity at 5.0. Using polygalacturonate as a substrate, the calculations of KM, Vmax and kcat were 0.65 mg∙mL−1, 1800 μmol∙min−1∙mg−1 and 35.97 s−1, respectively. The enzyme was stable for more than 3 h at 50˚C at pH 5.0; otherwise, at lower or higher pH values, the PGaseLg was less stable. The influence of several metals, EDTA and β-mercaptoethanol on enzyme activity was also determined. Thin layer chromatography (TLC) analyses indicated that PGaseLg is an exopolygalacturonase.
Keywords:
Polygalacturonase, L. gongylophorus, A. sexdens, Plague Control

1. Introduction
The symbiotic relationship of the leaf-cutter ant Atta sexdens with basidiomycete Leucoagaricus gongylophorus is an important Neotropical herbivorous system [1] -[3] . The ecological efficiency of symbiosis resulted in an agricultural pest responsible for major crop losses on Brazilian savannah [4] [5] . The ants cultivate an underground fungus-garden in specialized nest compartments [6] . Workers cut, transport and fragment fresh leaves on which they place fecal droplets, inoculating them with the symbiotic fungus [3] [7] . The mature fungi have an essential nutritional role for the ant colonies, providing specialized hyphae known as gongylidium [1] [4] . These gongylidia feed larval and alated forms, meanwhile complementing the workers’ diet [1] [7] [8] .
Biochemical issues are relevant in the intricate interdependence of mutualism [2] [7] . One of the key factors of the maintenance of the symbiosis is the mushroom supply of ants with hydrolytic enzymes and easily assimilated nutrients [4] [9] . One of the questions that had remained unknown is about the metabolism of polysaccharides by the symbiotic fungus [2] [5] . The cellulase was firstly thought to be the main fungal enzyme in the interaction [5] [7] [10] . Nevertheless, this enzyme is less produced, and cellulose remains intact in the cultivated garden [7] [11] . Otherwise results regarding hydrolytic enzyme profiles have revealed, among others, enzymes such as xilanases [12] , proteinases [13] , amylases [5] and broad production of pectinases [2] [3] [8] by the fungus. Presently, the fungal origin of proteinases and pectinases found in the fecal fluid of adult workers is well established [3] [7] [13] .
Pectin is a major structural component of plant cell walls, forming a gel-like matrix that is particularly abundant at cell wall interfaces in the middle lamella region of leaves, where it regulates intercellular adhesion [7] . A highly heterogeneous branched and esterified polymer of plant cell walls requires a large range of enzymes for complete degradation [14] . Pectinase is a generic term for heterogeneous enzyme complexes from pectin metabolism [15] . Beyond the role in plant physiology, several industrial applications are reported for plant materials treatment [16] [17] . Pectinases are classified as desterifying and depolymerizing. Pectin methyl-esterases (EC 3.1.1.11) and pectin acetyl-esterases (EC 3.1.1.6) are responsible for transesterification of esterified residues with the consequent liberation of acid to the environment [18] . The depolymerizing enzymes cleave the glycosidic linkage of adjacent residues by two distinct mechanisms. Pectin lyases (EC 4.2.2.10), or pectin transeliminases, digest esterified pectin by ß-elimination, producing oligogalacturonides 4,5-insaturated in a non-reducing position [19] . Polygalacturonases refer specifically to pectin hydrolases whose substrate is unsubstituted 1,4- polygalacturonic acid [20] . There are two classes of hydrolases, referred to as endopolygalacturonase (EC 3.2.1.15) and exopolygalacturonase (EC 3.2.1.67). Endo-PG hydrolyses the soft pectin randomly, producing a large range of oligomers [21] , and exo-PG acts in terminal residues, resulting in polygalacturonic acid monomers [18] . Several other glycosidases are involved in the complex as auxiliary enzymes [16] . Fungi [17] [22] [23] , bacteria [14] [24] and even insects [25] are known as sources of pectinases, among them, the symbiotic mold [1] [2] [7] [11] . Moreover, the symbiotic relation seems to be dependent on pectinases that are over expressed in gongylidia as an adaptation for nutritional interactions [1] [7] . The degradation of the pectic network would be necessary to access protein and starch inside the plant cells, rather than pectin being an important nutrient source in its own right [11] [26] .
As parts of our efforts to better understand the pectinases expressed by L. gongylophorus, we identified a bifunctional polygalacturonase/xylanase [12] , and we have constructed a three-dimensional model and analyzed the possible mechanism of the enzyme action [27] . Furthermore, a pectinolytic extract from L. gongylophorus has been partially characterized in respect to anchorage on magnetic support [28] . Since these results were obtained from an unpurified enzyme in terms of total activity and apparent kinetic parameters, the purification of the enzyme generated the chance for a settlement in these parameters. The present study aimed to characterize optimal temperature and pH, heat, chemical stability and kinetics of a homogeneous purified exopolygalacturonase from L. gongylophorus (PGaseLg). The mechanism of action and the effect of different metals and chemicals on the activity of the purified enzyme were also evaluated.
2. Material and Methods
2.1. Chemicals
All chemicals were of analytical grade; Milli-Q water was used throughout the experiments. Citric acid, formic acid, K3[Fe(CN)6], Na2S2O3, NH4HCO3, (NH4)2SO4, Na2HPO4, KCl, NH4Cl, MgCl2, CaCl2, FeCl3, MnCl2, CoCl2, CuSO4, NiCl2, ZnSO4, HgCl2, PbCl2, EDTA, β-mercaptoethanol, MES (2-[N-morpholino]ethanesulfonic acid), dithiothreitol (DTT), iodoacetamide (IAA), yeast nitrogen base (YNB), 3,5-dinitrosalicylic acid (DNS) and polygalacturonic acid were purchased from Sigma (St. Louis, MO). A Bradford method kit was purchased from Bio-Rad Laboratories (Hercules, CA) and trypsin gold, MS grade, from Promega Corporation (Madison, WI). A McIlvaine’s buffer solutions system (Citrate/phosphate, pH 5.0) was applied as a working buffer. The pH values were obtained by mixing Na2HPO4 0.2 mol∙L−1 and citric acid 0.1 mol∙L−1 at a suitable ratio.
2.2. Inductive Culture of L. gongylophorus for Polygalacturonase Production
The CCTI strain of L. gongylophorus (isolated from an A. sexdens nest) was donated by the Center of Studies on Social Insects (UNESP, Rio Claro, Brazil). The mold was cultivated at room temperature in stationary liquid cultures of an inductive medium (YNB 6.7 mg∙mL−1 and polygalacturonic acid 5.0 mg∙mL−1 in working buffer) [5] . The inoculums consisted of mycelial mass collected from solid cultures. After 30 days, the culture (1 liter) containing the secreted enzymes was filtered through a 0.45 μm-pore-sized membrane, and the volume was reduced 10 times by lyophilization. This solution was identified as a crude enzyme extract.
2.3. Enzyme Activity Assay
Polygalacturonase activity was evaluated according to the Miller method for reducing sugar determination [29] by mixing the enzyme with a solution containing polygalacturonic acid in final 5.00 mg∙mL−1 in a working buffer. The reaction mixture was kept at 60˚C for zero, 30 and 60 min., after which 100 μL aliquot was added to 400 μL of DNS reagent and heated to 80˚C for 5 min. The reaction vials were cooled centrifuged (16,000 g for 5 min.) and then added to 500 μL of H2O. The values of absorbance at 540 nm were determined. Product formation was quantified in reference to a standard curve prepared with 0 to 100 mmol∙L−1 galacturonicacid. Protein content was evaluated according to the Bradford protocol [30] . The enzymatic activity was expressed in units of activity, U. One U corresponds to the production of 1 μmol of reducing sugars per minute per mg of total protein. The activity values were obtained from three independent experiments and were expressed as weighed mean ± standard deviation.
2.4. Enzyme Purification
Polygalacturonase (PGaseLg) purification chromatographic procedures were carried out in the AKTA-FPLC system (GE Healthcare). The centrifugation through the purification procedures was conducted at 12,000 ×g for 20 min. at 4˚C. Dialyses, unless stated, were carried out against the working buffer at 4˚C.
(NH4)2SO4 salting out: prior to the chromatography columns, the stepwise precipitation with (NH4)2SO4 was useful in clarifying the crude extract. In the first step, ammonium sulfate was added to the crude enzyme solution up to 30% of saturation, and the insoluble contents were then removed by centrifugation and discharged. The supernatant received new additions of salt until the saturation reached 70%. The solution reposed overnight and was centrifuged to produce the precipitant within the interval from 30% to 70% of salt saturation. This pellet was solubilized in a 10 mL working buffer and dialyzed.
Gel filtrationchromatography: 2 mL of the salted-out 30% - 70% active fractions were loaded onto a Superdex S-75 (GE Healthcare) column (16 × 600 mm) pre-equilibrated with a working buffer with NaCl 100 mmol∙L−1. Elution was carried out with the same buffer at a flow rate of 1.0 mL∙min−1. The PGaseLg activity- eluted fractions were pooled and dialyzed against a 20 mmol∙L−1 acetate buffer, pH 5.0.
Cation exchange chromatography: 1 mL of the Superdex S-75 active fractions pool in a 20 mmol∙L−1 acetate buffer was applied onto a SPFF-Sepharose column (GE healthcare) pre-equilibrated with the same acetate buffer. The column was washed with 10 mL of the acetate buffer at a flow rate of 1 mL∙min−1, and the elution was with a NaCl linear gradient from 0 to 290 mmol∙L−1 in the same buffer. The PGaseLg activity was determined in desalted fractions, and the active peak was pooled for characterization.
2.5. Gel Electrophoresis
Each step of enzyme purification was followed by electrophoresis in 15% (v/v) polyacrylamide gel in denaturing conditions (SDS-PAGE), as described by Laemmli [31] . After the run, the gel was submitted to a silver stain, and the single band, from SPFF chromatography, was set out for IAA alkylation and trypsin digestion before MS analysis. Molecular weights of the standard proteins were β-galactosidase, 116 kDa; bovine serum albumin, 66.2 kDa; ovalbumin, 45.0 kDa; lactate dehydrogenase, 35.0 kDa; REase Bsp98I, 25.0 kDa; β-lactoglobulin, 18.4 kDa; and lysozyme, 14.4 kDa.
2.6. Characterization of Purified Enzymes
2.6.1. Influence of the Temperature on Purified Enzyme
The temperature effect on enzyme activity was determined at 30˚C, 40˚C, 45˚C, 50˚C, 55˚C, 60˚C, 65˚C, 75˚C and 80˚C using polygalacturonic acid as the substrate. Experiments were expressed in enzyme activity (U) versus temperature plot. Based on the temperature assay, it was possible to calculate activation energy.
2.6.2. The Thermal Stability
Thermal inactivation was evaluated by incubating the purified PGaseLg at 30˚C, 40˚C and 50˚C. Reaction vials were kept, at pH 5.0, under mild agitation for 10 hours in the experimented temperature. At each 30 min., 50 μL aliquots were gathered and submitted to ice bath, and the enzyme residual activity (U%) was determined at 60˚C and pH 5.0. The residual activity was plotted as a function of the incubation temperature.
2.6.3. Influence of the pH
The optimal pH value of PGaseLg activity was determined by assaying the purified enzyme at 60˚C in the working buffer at pH values from 2.5 to 6.5 (slope of 0.5). Activities (U) were plotted against pH values.
2.6.4. The Chemical Stability
To evaluate the influence of pH on the stability of the enzyme, it was kept for six hours at 50˚C in the working buffer at pH values of 3.0, 4.0, 5.0, 6.0 and 7.0 or the Tris/glycine buffer for pH 8.0. The residual activities of 50 μL aliquots were determined at optimal activity conditions. The determined half-lives of each experiment were plotted versus pH treatment.
2.6.5. Michaelis-Menten Constant
The PGaseLg kineticconstant (KM) was determined with ten substrate concentrations ranging from 0.10 to 5.0 mg∙mL−1 of galacturonic acid or 74% esterified pectin. KM and Vmax values were calculated using the double reciprocal Lineweaver-Burk plots.
2.6.6. Effect of Cations and Other Substances on PGaseLg Activity
The effect of a number of metal ions and other reagents on enzyme activity in the assay medium was tested. Pure PGaseLg was assayed in the presence of KCl, NH4Cl, MgCl2, CaCl2, FeCl3, MnCl2, CuSO4, CoCl2, NiCl2, ZnSO4, PbCl2, HgCl2, EDTA and β-mercaptoethanol at 5.0 mmol∙L−1. The assays were conducted with the 50 mmol∙L−1 MES buffer, pH 5.5. Before the assay, the enzyme solution was dialyzed against this same buffer. Residual activity was determined in triplicate at each treatment.
2.6.7. Determination of the PGaseLg Oligomerization State
The SDS-PAGE running was used for molecular mass determination, and the oligomerization state determination of PGaseLg was performed by loading the enzyme on a Superose 12 HR column (10 × 30 cm) calibrated with bovine serum albumin (BSA) as molecular mass standard.
2.7. Thin Layer Chromatography
Thin layer chromatography (TLC) analyses of polygalacturonic acid and 74% esterified pectin hydrolysis products were performed on heat-inactivated samples from overnight digestion, at 40˚C in the working buffer. Aliquots of respective pectic hydrolysate were 10 times concentrated and spotted on 8 × 10 cm silicagel 60G aluminum sheets (Merck, Germany). Mono-, di-, tri- and polygalacturonic acid were applied as standard. The chromatography was performed using the ascending method; the mobile phase consisted of a 5:3:2 mixture of n-butanol: H2O: acetic acid. For visualization of the spots, the dried plate was sprayed with 10% sulfuric acid in methanol followed by heating at 105˚C for 5 min.
2.8. LC-MS/MS Analysis
Sliced and washed SDS-PAGE slabs were destained using freshly prepared K3[Fe(CN)6] 30 mmol∙L−1 and Na2S2O3 100 mmol∙L−1 mixed with a 1:1 ratio and added to cover the gel pieces. Afterward, the gels were washed until they were clear by ammonium bicarbonate 100 mmol∙L−1. The gels were emerged in acetonitrile and dried in a vacuum centrifuge. After the reduction with dithiothreitol 10 mmol∙L−1 in ammonium bicarbonate 50 mmol∙L−1, the samples were alkylated by iodoacetamide 55 mmol∙L−1 in NH4HCO3 50 mmol∙L−1. Alkylated peptides were digested by trypsin gold, mass spectrometry grade, in an ammonium bicarbonate buffer (25 mmol∙L−1, pH 8.0) and extracted by acetonitrile/formic acid 0.1% (40:60). Samples were desalted by ZipTip® and kept at 20˚C until the LC-MS/MS analysis.
Tryptic digested peptides were analyzed by online nanoflow LC-MS on an EASY-nLC II system (Thermo Scientific) connected to an LTQ-OrbitrapVelos instrument (Thermo Scientific) via a Proxeonnanoelectrospray ion source. Peptides were separated with a linear gradient from 0% to 60% acetonitrile (0.1% formic acid) on an analytical EASY-Column (10 cm, ID75 µm, 3 µm, C18-Thermo Scientific, 300 nL∙min−1) previously trapped in a pre-column EASY-Column (2 cm, ID100 µm, 5 µm, C18-Thermo Scientific). An LTQ-OrbitrapVelos mass spectrometer was operated using DDA (data-dependent acquisition) in positive ion mode. The 20th most intense precursor ions were selected for CID fragmentation. Full MS scans were performed with 60,000 full-width half-maximum (FWHM) nominal resolution settings (m/z range 400 - 1200, collision energy 35 eV, activation Qz of 0.250, activation time 10 ms). The minimum signal threshold was 15,000 counts, and for dynamic exclusion, 1 repeat count was considered with a duration of 30 s. The instrument was calibrated externally according to manufacturer’s instructions.
3. Results and Discussions
3.1. Purification of PGaseLg
Polygalacturonic acid, added to the culture broth, was efficient in the PGaseLg induction whose purification was carried out as summarized in Table 1. The purification to homogeneity was completed after ammonium sulfate salting out and two chromatographic steps. The activity in ammonium sulfate salting out (30% - 70% saturation) was 82% recovered in the precipitated proteins with a purification fold of 3.2. Superdex S-75 gel filtration, Figure 1(a), provided a purification in the order of 12.9-fold with a yield of 61.2%. SPFF, Figure 1(b), cation exchange chromatography promoted the homogeneous purity. Along the sodium chloride gradient (0 - 280

Table 1. Summary of the purification of polygalacturonase from L. gongylophorus.

Figure 1. Elution profiles for (a) Gel filtration chromatography of active fraction of PGaseLg (NH4)2SO4 clarified extract on Superdex S-75 column; (b) Cation exchange chromatography (SPFF Sepharose) of pooled active eluate from Superdex-75.
mmol∙L−1), three isolated bands were eluted, and PGaseLg activity was detected in the first, corresponding to 85 mmol∙L−1 of salt concentration. An overall purification of up to 30.8-fold with a 48.8% recovery was achieved (Table 1).
The homogeneity of the purified PGaseLg was demonstrated by the presence of one single protein band on SDS-PAGE stained with silver salts (Figure 2).
To calculate the molecular weight of the purified PGaseLg, the electrophoretic mobility against the logarithm molecular weights of known polypeptides was plotted and compared to the electrophoretic mobility of the PGaseLg. The calculated molecular weight of the PGaseLg was approximately 39.0 kDa. This molecular mass is in agreement with previously reported PGase from other sources, such as Rhizomucorpusillus [18] and Tricodermaharzianum [32] (about 31 kDa), Acrophialophoranainiana (31 to 35.5 kDa) [14] , Saccharomyces cerevisiae strain IM1-8b (36 kDa) [33] , Neurosporacrassa (37 kDa) [23] or Rhizopusoryzae (37.44 kDa) [34] . Higher molecular mass PGase is known, such as from E. carotovora (43 kDa) [24] and A. giganteus (69.9 kDa) [35] . Size exclusion chromatography enables the separation of impurities from the target protein, and the specific elution profile provides an estimate of its molecular weight/size; it also furnishes information of its primary oligomeric state [36] . The lower than 66 kDa PGaseLg apparent molecular weight determined by Superose 12 HR gel filtration chromatography is indicative that the enzyme was isolated as a monomer. These results are in agreement with published results for polygalacturonases from Burkholderiaglumae [37] and T. harzianum [32] .
3.2. PGaseLg Identification by MS/MS Database Search
The PGaseLg was identified in the SDS-PAGE gel; the band was excised from the gel and treated with trypsin, and the peptides were analyzed by online LC-MS nanoflow. The MS spectra were searched for on different databases with two different search engines and in-house Proteome Discoverer 1.4 software (Thermo, USA). Databases with different numbers of sequences were used to increase the protein identification confidence. The databases were downloaded by typing “Leucoagaricus” (202 protein sequences) as a keyword on both NCBI and Uniprot sites using the SEQUEST search engine (Proteome Discovery 1.4). The “Fungi_NCBI” (2,204,168 protein sequences) database was used directly from the MASCOT 2.2.4 search engine with NCBInr filtered by fungi taxonomy. The identified peptide sequences with significant sequence coverage with a polygalacturonase sequence of the L. gongylophorus are presented in Table 2.
The database search of mass spectrometry analysis identified the PGaseLg with an annotated polygalacturonase at NCBI (Accession number 317468146) with a theoretical molecular weight of 37.034 kDa. Instead, in SDS-PAGE, a PGaseLg molecular weight of 39.04 kDa was observed. This fact may be explained by glycosylation, which is not annotated in the deposited L. gongylophorus polygalacturonase sequence, which is commonly assigned to other glycosidases.
The amino acid sequence similarity search in the Swissprot database indicated that peptides were similar to polygalacturonase sequences in glycosyl hydrolase family 28. The four amino acid groups (NTD, DD, HG and RIK), presumably involved in catalysis, are conserved in these polygalacturonases in GH family 28 and are situated at AA 177 - 179, 199 - 200, 221 - 222 and 252 - 254, respectively (Figure 3).

Figure 2. SDS-PAGE analysis of each purification step. MW: molecular mass marker (116.0, 66.2, 45.0, 35.0, 25.0, 18.4 and 14.4 kDa); Lane 1: salted-out precipitant; Lane 2: Superdex S-75 gel filtration eluted protein (pooled 60-69 fraction); Lane 3: SPFF column product (purified enzyme).

Figure 3. Polygalacturonase sequence from L. gongylophorus (GenBank: ADV30326.1). In bold are the peptide sequences identified by mass spectrometry for PGaseLg. The four amino acid groups (NTD, DD, HG and RIK) involved in the catalytic mechanisms of the family 28 polygalacturonases are indicated in underline. The seven cysteine residues are in bold and in undeline.
Table 2. Peptide coverage sequences of first hit to enzyme identified in gel band.
3.3. Physicochemical Characterization
Using polygalacturonic acid as a substrate, the optimum temperature for activity of the enzyme was 60˚C, Figure 4(a). The pattern of polygalacturonase activity from PGaseLg enzyme showed a typical asymmetric thermal profile. Determinations of optimal temperature for pectinases from different sources are closely related, for instance 60˚C for Acrophialophora nainiana [14] or 65˚C for Paenibacillus sp [38] and Paecilomyces variotii [39] . Optimal temperature of polygalacturonases from Rhizomucor pusillus [18] , Penicillium viridicatum [15] , Thermoascus aurantiacus [20] , Pycnoporus sanguineus [40] and A. giganteus [35] ranged from 55˚C to 65˚C. Other optimal temperatures are registered as 45˚C for Neurospora crassa [23] , 40˚C for T. harzianum [32] and E. carotovora [24] . The activation energy was determined by using the Arrhenius equation from the slope of the plot of the natural logarithm of the activation rate constants of purified PGaseLg versus the reciprocal absolute temperature (˚K), and it was estimated at 45.6 kJ∙mol−1 (10.90 kcal∙mol−1). This result was similar to the 68.86 kJ∙mol−1 observed in polygalacturonase from E. carotovora [24] . This activation energy is lower than a purified polygalacturonase from tomatoes (270.6 kJ∙mol−1) [41] , but higher than commercial preparations (free 18.9 kJ∙mol−1 or immobilized 20.7 kJ∙mol−1) [42] .
The effect of pH on the purified PGaseLg activity toward polygalacturonic acid was examined at 60˚C. As shown in Figure 4(b), the enzyme showed an optimum pH for hydrolase activity at 5.0 in the working buffer. The enzyme activity decreased to 50% at pH levels lower than 3.5 or higher than 6.0. A similar pH profile was reported for PGs from T. harzianum [32] , E. carotovora [24] and Fusarium moniliforme [43] . The loss of activity at neutral and basic pH values is due to the ionization state of amino acidic residues involved in substrate binding and in catalysis, and it has been reported for other glycoside hydrolases [21] . Moreover, the substrate pKa of polygalacturonic acid is 3.7, which requires acidic pH values. However, the optimal pH of pectinases from Streptomyces lydicus [44] and Neurospora crassa [23] was determined at 6.0 and from Acrophialophora nainiana [14] at 8.0. Acidic endopolygalacturonases are produced by Aspergillus kawachii [22] , active at pH 2.0

Figure 4. Effect of (a) temperature and (b) pH on enzyme activity. Assay conditions as described in experimental section.
in the presence of stabilizers, and by phytopathogen Burkholderia cepacia, with an optimum pH of 3.5 [21] .
The stability of an enzyme is an important parameter since maintenance of activity over a long period is important in designing pectinases reactors and also for reproducibility of data acquisition. With respect to temperature, in the absence of a substrate, PGaseLg showed >75% of the original activity at 30˚C and 40˚C, for 10 hours. At 50˚C, the enzyme lost 50% of its initial activity ~240 minutes (Figure 5(a)). Concerning the effect of the temperature on the enzymatic activity, PGaseLg stability is in agreement with the data already reported for polygalacturonase from bacterial [24] or fungal [21] sources. Stabilization mechanisms (immobilization [16] , salts [15] and stabilizers [22] ) have been identified using mesophilic and thermophilic enzymes.
Thermal stability contrasts with optimal activity; in the presence of a substrate, at 60˚C, the enzyme completely loses the activity in less than 15 minutes. The effect of substrate protection is demonstrated by the fast deactivation of the enzyme at its optimal temperature of reaction. The results showed that assay conditions of pH and temperature are not the best conditions for stability, which is in agreement with the fact that catalytic performance (activity) and stability of pectinases are quite different aspects [44] . These observations typify the PGaseLg as a mesophilic enzyme.
In another experiment, PGaseLg was maintained at 50˚C at different pH values (3.0 to 8.0) for 360 minutes. Samples were taken at each 15 minutes until 120 minutes and then at every 30 minutes as exhibited in Figure 5(b). At lower and neutral pH values, the recovery of enzyme activity decayed rapidly (≤ 1 h); on the other hand, at optimal pH condition (pH 5.0), the enzyme preserved 50% of its initial activity for 240 min. The half-life time at 50˚C of PGaseLg, experimentally determined, was plotted as a function of pH of treatment, Figure 5(c).
The resulting profile, in Figure 5(c), reveals the higher stability at pH 5.0 and the lower stability when the conditions change. The herein reported results are in agreement with those reported for P. variotii [39] or for Kluyveromyces marxianus [45] . This is a typical characteristic of fungal polygalacturonase acid [18] .
3.4. Kinetic Characterization
A typical Michaelian kinetic was observed for the hydrolysis of polygalacturonic acid and also for 74% esterified pectin at pH 5.0 and 60˚C by PGaseLg. When polygalacturonic acid was the substrate, the KM, Vmax and Kcat values were 0.65 mg∙mL−1, 1800 μmol∙min−1∙mg−1 and 35.97 s−1, respectively, while in the hydrolysis of 74% esterified pectin, the values of KM, Vmax and Kcat were 1.18 mg∙mL−1, 310 μmol∙min−1∙mg−1 and 6.19 s−1, respectively. From these results, it can be reported that PGaseLg has higher affinity toward polygalacturonic acid than esterified pectin.
The KM values of polygalacturonase from A. nainiana [14] , R. pusillus [18] , T. harzianum [32] , A. giganteus [35] , S. cerevisiae [33] , N. crassa [23] and S. lydicus [44] for polygalacturonic acid at different buffers range from 0.22 to 5.0 mg∙mL−1, in agreement with the observed PGaseLg KM value. It should be said that a wide

Figure 5. Stabilities of purified PGaseLg. (a) Thermal deactivation of the enzyme at 30˚C (□), 40˚C (○) and 50˚C (Δ); (b) Chemical effect on stability of the enzyme at 50˚C under 3.0 (■), 4.0 (□), 5.0 (●), 6.0 (○), 7.0 (▲) and 8.0 (Δ) pH values; (c) Profile of experimental half-lives of PGaseLg in function of pH incubation.
range of kinetic parameter values has been reported for polygalacturonases from various sources of microorganisms. This may be attributed to differences in assay procedures [14] .
3.5. Effect of Metals and Chemicals on PGaseLg Activity
The susceptibility of PGaseLg to several cations, EDTA and β-mercaptoethanol at 1.0 and 5.0 mmol∙L−1 was investigated. Table 3 points out the activation of the enzyme in both concentrations of K+, 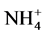 and the Mg2+ cations. In the case of K+ 5 mmol∙L−1, the enzyme activity increased fivefold.
and the Mg2+ cations. In the case of K+ 5 mmol∙L−1, the enzyme activity increased fivefold.
The PGaseLg inactivation observed in the presence of Hg2+ and Ni2+ and the inhibitory effect of other tested cations are indicative of active site blockage by metal complexation; the recovery activity for these cations is ordered in the following sequence: Ca2+ < Cu2+ < Co2+ < Pb2+ < Fe3+ < Zn2+ < Ni2+ ≈ Hg2+.
Manganese promoted 34% of enzyme activation at low concentration, but at 5.0 mmol∙L−1, the activity dropped to about 11%, as is observed for the PGase of A. giganteus(35). Taken together, apart from the specific responses, these results are in consonance with the literature which characteristically exhibits the inhibition by ordinary metals and the deactivation by heavy metals [15] [32] [39] .
EDTA poorly affected the PGaseLg activity at lower experimental concentrations but the effect was amplified at higher concentration, indicating metal complexation in the catalysis process. β-mercaptoetanol promoted a similar effect on enzyme, which is an expected result since the deduced amino acid sequence of the enzyme (Figure 3) predicted one free cysteine, this residue would perform a critical role in the catalysis [35] .
3.6. TLC Analysis of Hydrolyzates
The TLC technique was applied to investigate the PGaseLg mechanism using the hydrolysis products from enzyme action on polygalacturonic acid and 74% esterified pectin. It was observed that from polygalacturonic acid digestion, the only soluble product released was monogalacturonic acid (Figure 6). Based on this result, the enzyme can be classified as exopolygalacturonase (EC 3.2.1.67). In contrast with the wide distribution of endopolygalacturonase, the occurrence of exopolygalacturonases is less frequent. Exopolygalacturonases are classified as a fungal type, which mainly releases monogalacturonic acid, and a bacterial type, which mainly produces digalacturonic acid. The isolated polygalacturonase from L. gongylophorus is a typical fungal exopolygalacturonase.
Even upon prolonged incubation, the action of the enzyme on 74% esterified pectin did not exhibit the same accumulation of the final products. This observation is due to interruption of exopolygalacturonase action on esterified residues of the substrate since methyl esters limit an exopolygalacturonase action.
4. Conclusions
This paper is the first report of the isolation to electrophoretic homogeneity of an acidic exopolygalacturonase
Table 3. Influence of different metals and chemicals, at 1.0 and 5.0 mmol∙L−1, on PGaseLg activity.
 1 2 3 P A B
1 2 3 P A B
Figure 6. Reaction products hydrolyzed from pectin and polygalacturonic acid by the PGaseLg. The reaction mixture contained 5.0 mg∙mL−1 of polygalacturonic acid (A) and 74% esterified pectin (B) in McIlvaine’s buffer (pH 5.0); 1, monogalacturonic acid; 2, digalacturonic acid; 3, trigalacturonic acid; and P, polygalacturonic acid. The arrow points to galacturonic acid as the main hydrolysis product of PGaseLg on polygalacturonic acid.
secreted by L. gongylophorus. The 37 kDa enzyme was biochemically and biophysically characterized as a typical fungal mesophilic exopolygalacturonase.
Polygalacturonases have been listed as an important factor in maintaining leaf-cutting ants/fungus symbiosis and, thus, inhibiting their activities can lead to the development of cutting ants control systems. In order to contribute to the study of these enzymes as targets for inhibition, we have described a xylanase/polygalacturonase bifunctional enzyme [12] [27] and we have also immobilized the partially purified PGase from L. gongylophorus in magnetic particles [28] . The purification and characterization of the polygalacturonase have increased the knowledge of the nature and properties of this enzyme, which is essential for understanding its structure, mechanisms of action, stability and inhibition studies.
Acknowledgements
This work was supported by a grant 2011/21955-3 from the Sao Paulo Research Foundation (FAPESP). Paulo R. Adalberto is supported by a grant from the Brazilian National Research Council (CNPq 167524/2013-5). We thank to Prof. Fernando C. Pagnocca (UNESP, Rio Claro) for provided L. gongylophorus.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
Cite this paper
Paulo R.Adalberto,Camilla C.Golfeto,Fernando G.Almeida,DouglasFerreira,Quezia B.Cass,Dulce H. F.Souza,Ariele C.Moreira, (2016) Characterization of an Exopolygalacturonase from Leucoagaricus gongylophorus, the Symbiotic Fungus of Atta sexdens. Advances in Enzyme Research,04,7-19. doi: 10.4236/aer.2016.41002
References
- 1. De Fine Licht, H.H., Boomsma, J.J. and Tunlid, A. (2014) Symbiotic Adaptations in the Fungal Cultivar of Leaf- Cutting Ants. Nature Communications, 5, 5675. http://dx.doi.org/10.1038/ncomms6675
- 2. Erthal, M., Silva, C.P., Cooper, R.M. and Samuels, R.I. (2009) Hydrolytic Enzymes of Leaf-Cutting ant Fungi. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 152, 54-59. http://dx.doi.org/10.1016/j.cbpb.2008.09.086
- 3. Schiøtt, M., Rogowska-Wrzesinska, A., Roepstorff, P. and Boomsma, J. J. (2010) Leaf-Cutting ant Fungi Produce Cell Wall Degrading Pectinase Complexes Reminiscent of Phytopathogenic Fungi. BMC Biology, 8, 156. http://dx.doi.org/10.1186/1741-7007-8-156
- 4. Sternberg, L.S., Pinzon, M.C., Moreira, M.Z., Moutinho, P., Rojas, E.I. and Herre, E.A. (2007) Plants Use Macronutrients Accumulated in Leaf-Cutting ant Nests. Proceedings of the Royal Society B: Biological Sciences, 274, 315-321. http://dx.doi.org/10.1098/rspb.2006.3746
- 5. Silva, A., Bacci, M., Pagnocca, F.C., Bueno, O.C. and Hebling, M.J.A. (2006) Production of Polysaccharidases in Different Carbon Sources by Leucoagaricus gongylophorus Möller (Singer), the Symbiotic Fungus of the Leaf-Cutting Ant Atta sexdens Linnaeus. Current Microbiology, 53, 68-71. http://dx.doi.org/10.1007/s00284-005-0431-1
- 6. Mikheyev, A.S., Mueller, U.G. and Abbot, P. (2006) Cryptic Sex and Many-to-One Coevolution in the Fungus-Growing ant Symbiosis. Proceedings of the National Academy of Sciences of the United States of America, 103, 10702-10706. http://dx.doi.org/10.1073/pnas.0601441103
- 7. Moller, I.E., De Fine Licht, H.H., Harholt, J., Willats, W.G.T. and Boomsma, J.J. (2011) The Dynamics of Plant Cell- Wall Polysaccharide Decomposition in Leaf-Cutting ant Fungus Gardens. PLoS ONE, 6, e17506. http://dx.doi.org/10.1371/journal.pone.0017506
- 8. Rønhede, S., Boomsma, J.J. and Rosendahl, S. (2004) Fungal Enzymes Transferred by Leaf-Cutting Ants in Their Fungus Gardens. Mycological Research, 108, 101-106. http://dx.doi.org/10.1017/S0953756203008931
- 9. Schiøtt, M., Licht, H.H.D.F., Lange, L. and Boomsma, J.J. (2008) Towards a Molecular Understanding of Symbiont Function: Identification of a Fungal Gene for the Degradation of Xylan in the Fungus Gardens of Leaf-Cutting Ants. BMC Microbiology, 8, 40. http://dx.doi.org/10.1186/1471-2180-8-40
- 10. Bacci, M., Bueno, O.C., Rodrigues, A., Pagnocca, F.C., Somera, A.F. and Silva, A. (2013) A Metabolic Pathway Assembled by Enzyme Selection May Support Herbivory of Leaf-Cutter Ants on Plant Starch. Journal of Insect Physiology, 59, 525-531. http://dx.doi.org/10.1016/j.jinsphys.2013.02.007
- 11. Siqueira, C.G.D., Bacci, M., Pagnocca, F.C., Bueno, O.C. and Hebling, M. (1998) Metabolism of Plant Polysaccharides by Leucoagaricus gongylophorus, the Symbiotic Fungus of the Leaf-Cutting Ant Atta sexdens L. Journal of Applied & Environmental Microbiology, 64, 4820-4822.
- 12. Moreira, A.C., Ferreira, D., Almeida, F.G., Rodrigues, E., Fernandes, J.B., Silva, M.F., Vieira, P.C., Pagnocca, F.C. and Souza, D.H.F. (2014) Molecular and Kinetic Characterization of Two Extracellular Xylanases Isolated from Leucoagaricus gongylophorus. Applied Biochemistry and Biotechnology, 173, 694-704.
- 13. Kooij, P.W., Rogowska-Wrzesinska, A., Hoffmann, D., Roepstorff, P., Boomsma, J.J. and Schiøtt, M. (2014) Leucoagaricus gongylophorus Uses Leaf-Cutting Ants to Vector Proteolytic Enzymes towards New Plant Substrate. The International Society for Microbial Ecology Journal, 8, 1032-1040. http://dx.doi.org/10.1038/ismej.2013.231
- 14. Celestino, S.M.C., Freitas, S.M., Javier Medrano, F., Valle de Sousa, M. and Ferreira, E.X. (2006) Purification and Characterization of a Novel Pectinase from Acrophialophora nainiana with Emphasis on Its Physicochemical Properties. Journal of Biotechnology, 123, 33-42. http://dx.doi.org/10.1016/j.jbiotec.2005.10.024
- 15. Gomes, E., Leite, R.S.R., da Silva, R. and Silva, D. (2009) Purification of an Exopolygalacturonase from Penicillium viridicatum RFC3 Produced in Submerged Fermentation. International Journal of Microbiology International Journal of Microbiology, 2009, Article ID: 631942.
- 16. Lara-Márquez, A., Zavala-Páramo, M.G., López-Romero, E. and Camacho, H.C. (2011) Biotechnological Potential of Pectinolytic Complexes of Fungi. Biotechnology Letters, 33, 859-868. http://dx.doi.org/10.1007/s10529-011-0520-0
- 17. Benoit, I., Coutinho, P.M., Schols, H.A., Gerlach, J.P., Henrissat, B. and de Vries R.P. (2012) Degradation of Different Pectins by Fungi: Correlations and Contrasts between the Pectinolytic Enzyme Sets Identified in Genomes and the Growth on Pectins of Different Origin. BMC Genomics, 13, 321-332. http://dx.doi.org/10.1186/1471-2164-13-321
- 18. Siddiqui, M. A., Pande, V. and Arif, M. (2012) Production, Purification, and Characterization of Polygalacturonase from Rhizomucor pusillus Isolated from Decomposting Orange Peels. Enzyme Research, 2012, Article ID: 138634.
- 19. Gummadi, S.N. and Kumar, D.S. (2005) Microbial Pectic Transeliminases. Biotechnology Letters, 27, 451-458. http://dx.doi.org/10.1007/s10529-005-2197-8
- 20. Martins, E.S., Silva, D., Leite, R.S.R. and Gomes, E. (2007) Purification and Characterization of Polygalacturonase Produced by Thermophilic Thermoascus aurantiacus CBMAI-756 in Submerged Fermentation. Antonie van Leeuwenhoek, 91, 291-299. http://dx.doi.org/10.1007/s10482-006-9114-6
- 21. Massa, C., Degrassi, G., Devescovi, G., Venturi, V. and Lamba, D. (2007) Isolation, Heterologous Expression and Characterization of an Endo-Polygalacturonase Produced by the Phytopathogen Burkholderia capacia. Protein Expression and Purification, 54, 300-308. http://dx.doi.org/10.1016/j.pep.2007.03.019
- 22. Contreras Esquivel, J.C. and Voget, C.E. (2004) Purification and Partial Characterization of an Acidic Polygalacturonase from Aspergillus kawachii. Journal of Biotechnology, 110, 21-28. http://dx.doi.org/10.1016/j.jbiotec.2004.01.010
- 23. Polizeli, M.L.T.M., Jorge, J.A. and Terenzi, H.F. (1991) Pectinase Production by Neurospora crassa: Purification and Biochemical Characterization of Extracellular Polygalacturonase Activity. Journal of General Microbiology, 137, 1815-1823. http://dx.doi.org/10.1099/00221287-137-8-1815
- 24. Maisuria, V.B., Patel, V.A. and Nerurkar, A.S. (2010) Biochemical and Thermal Stabilization Parameters of Polygalacturonase from Erwinia carotovora subsp. carotovora BR1. Journal of Microbiology and Biotechnology, 20, 1077-1085. http://dx.doi.org/10.4014/jmb.0908.08008
- 25. Celorio-Mancera, M.P., Carl Greve, L., Teuber, L.R. and Labavitch, J.M. (2009) Identification of Endo- and Exo-Polygalacturonase Activity in Lygus hesperus (Knight) Salivary Glands. Archives of Insect Biochemistry and Physiology, 70, 122-135. http://dx.doi.org/10.1002/arch.20282
- 26. Aylward, F., Burnum-Johnson, K.E., Tringe, S.G, Teiling C., Tremmel, D.M., Moeller, J.A., Scott, J.J., Barry, K.W., Piehowski, P.D., Nicora, C.D., Malfatti, S.A., Monroe, M.E., Purvine, S.O., Goodwin, L.A., Smith, R.D., Weinstock, G.M., Gerardo, N.M., Suen, G., Lipton, M.S. and Currie, C.R. (2013) Leucoagaricus gongylophorus Produces Diverse Enzymes for the Degradation of Recalcitrant Plant Polymers in Leaf-Cutter Ant Fungus Gardens. Applied and Environmental Microbiology, 79, 3770-3778. http://dx.doi.org/10.1128/AEM.03833-12
- 27. Dos Santos, A.M. and Souza, D.H.F. (2015) Técnicas de Bioinformática-aplicadas ao estudo de poligalaturonases de fungos. Novas Edições Acadêmicas, São Paulo
- 28. Adalberto, P.R., Santos, F.J., Golfeto, C.C., Iemma M.R.C., Souza, D.H.F. and Cass, Q.B. (2012) Immobilization of Pectinase from Leucoagaricus gongylophorus on Magnetic Particles. Analyst, 137, 4855-4859. http://dx.doi.org/10.1039/c2an35682a
- 29. Miller, G.L. (1959) Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Analytical Chemistry, 31, 426-428. http://dx.doi.org/10.1021/ac60147a030
- 30. Bradford, M.M. (1976) A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Analytical Biochemistry, 72, 248-254. http://dx.doi.org/10.1016/0003-2697(76)90527-3
- 31. Laemmli, U.K. (1970) Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature, 227, 680-685. http://dx.doi.org/10.1038/227680a0
- 32. Mohamed, S.A., Farid, N.M., Hossiny, E.N. and Bassuiny, R.I. (2006) Biochemical Characterization of an Extracellular Polygalacturonase from Trichoderma harzianum. Journal of Biotechnology, 127, 54-64. http://dx.doi.org/10.1016/j.jbiotec.2006.06.009
- 33. Blanco, P., Sieiro, C., Reboredo, N.M. and Villa, T.G. (1997) Genetic Determination of Polygalacturonase Production in Wild-Type and Laboratory Strains of Saccharomyces cerevisiae. Archives of Microbiology, 167, 284-288. http://dx.doi.org/10.1007/s002030050445
- 34. Zhang, J., Henriksson, H., Szabo, I.J., Henriksson, G. and Johansson, G. (2005) The Active Component in the Flax- Retting System of the Zygomycete Rhizopus oryzae sb Is a Family 28 Polygalacturonase. Journal of Industrial Microbiology and Biotechnology, 32, 431-438. http://dx.doi.org/10.1007/s10295-005-0014-y
- 35. Pedrolli, D.B. and Carmona, E.C. (2010) Purification and Characterization of the Exopolygalacturonase Produced by Aspergillus giganteus in Submerged Cultures. Journal of Industrial Microbiology & Biotechnology, 37, 567-573. http://dx.doi.org/10.1007/s10295-010-0702-0
- 36. Irvine, G. B. (2003) High-Performance Size-Exclusion Chromatography of Peptides. Journal of Biochemical and Biophysical Methods, 56, 233-242. http://dx.doi.org/10.1016/S0165-022X(03)00061-7
- 37. Degrassi, G., Devescovi, G., Kim, J., Hwang, I. and Venturi, V. (2008) Identification, Characterization and Regulation of Two Secreted Polygalacturonases of the Emerging Rice Pathogen Burkholderia glumae. FEMS Microbiology Eco- logy, 65, 251-262. http://dx.doi.org/10.1111/j.1574-6941.2008.00516.x
- 38. Soriano, M., Diaz, P. and Pastor, F.I.J. (2005) Pectinolytic Systems of Two Aerobic Sporogenous Bacterial Strains with High Activity on Pectin. Current Microbiology, 50, 114-118. http://dx.doi.org/10.1007/s00284-004-4382-8
- 39. Damásio, A.R.L., Silva T.M., Maller, A., Jorge, J.A., Terenzi, H.F. and Polizeli, M.L. (2009) Purification and Partial Characterization of an Exo-Polygalacturonase from Paecilomyces variotii Liquid Cultures. Applied Biochemistry and Biotechnology, 160, 1496-1507. http://dx.doi.org/10.1007/s12010-009-8682-0
- 40. Quiroga, E.N., Sgariglia, M.A., Molina, C.F., Sampietro, D.A., Soberón, J.R. and Vattuone, M.A. (2009) Purification and Characterization of an Exo-Polygalacturonase from Pycnoporus sanguineus. Mycological Research, 113, 1404- 1410. http://dx.doi.org/10.1016/j.mycres.2009.09.007
- 41. Fachin, D., Smout, C., Verlent, I., Ly Nguyen, B., Van Loey, A.M. and Hendrickx, M.E. (2004) Inactivation Kinetics of Purified Tomato Polygalacturonase by Thermal and High-Pressure Processing. Journal of Agricultural and Food Chemistry, 52, 2697-2703. http://dx.doi.org/10.1021/jf030531h
- 42. Busto, M.D., García-Tramontín, K.E., Ortega, N. and Perez-Mateos, M. (2006) Preparation and Properties of an Immobilized Pectinlyase for the Treatment of Fruit Juices. Bioresource Technology, 97, 1477-1483. http://dx.doi.org/10.1016/j.biortech.2005.06.013
- 43. Niture, S.K. and Pant, A. (2004) Purification and Biochemical Characterization of Polygalacturonase II Produced in Semi-Solid Medium by a Strain of Fusarium moniliforme. Microbiological Research, 159, 305-314. http://dx.doi.org/10.1016/j.micres.2004.06.002
- 44. Jacob, N., Asha Poorna, C. and Prema, P. (2008) Purification and Partial Characterization of Polygalacturonase from Streptomyces lydicus. Bioresource Technology, 99, 6697-6701. http://dx.doi.org/10.1016/j.biortech.2007.10.002
- 45. Silva, E.G., Borges, M.F., Medina, C., Piccoli, R.H. and Schwan, R.F. (2005) Pectinolytic Enzymes Secreted by Yeasts from Tropical Fruits. FEMS Yeast Research, 5, 859-865. http://dx.doi.org/10.1016/j.femsyr.2005.02.006
NOTES

*Corresponding author.