Open Journal of Medical Microbiology
Vol.2 No.1(2012), Article ID:18260,8 pages DOI:10.4236/ojmm.2012.21002
Emergence of Optochin Resistance among Streptococcus pneumoniae in Japan
1Division of Microbiology, Department of Pathology and Microbiology, Faculty of Medicine, Saga University, Saga, Japan
2Division of Infection Control and Prevention, Saga University Hospital, Saga, Japan
3Marketing Department, Siemens Healthcare Diagnostics Kabushiki Kaisha, Tokyo, Japan
4Department of Clinical Laboratory, Kyurin Medical Laboratory, Fukuoka, Japan
5Hibiki Research Group for Clinical Microbiology, Fukuoka, Japan
Email: *miyamoth@cc.saga-u.ac.jp
Received January 14, 2012; revised January 30, 2012; accepted February 14, 2012
Keywords: Optochin; Streptococcus pneumoniae; H+-ATPase; MALDI-TOF MS; 16S rRNA
ABSTRACT
The optochin susceptibility test is a key method for differentiating Streptococcus pneumoniae from other a-hemolytic streptococci; however, optochin-resistant (Optr) S. pneumoniae have been reported in the last two decades. In this study, we investigated the isolation frequency of Optr S. pneumoniae in the North Kyushu area of Japan, and biochemically and genetically characterized Optr S. pneumoniae clinical isolates. Seven (0.68%) out of 1032 S. pneumoniae isolates collected by the North Fukuoka Infectious Diseases Working Group were found to be Optr S. pneumoniae. Resistant strains had MICs of optochin 2- to 64- fold higher than susceptible strains, possessed different antimicrobial resistance profiles, and belonged to different serotypes. All the seven Optr isolates had mutations in the nucleotide sequence code for subunit c of F0F1 ATPase. Three isolates had mutations in codon 48 (deduced amino acid substitution of valine with phenylalanine) and two isolates had mutations in codon 49 (substitution of alanine with threonine or serine). Of the remaining two isolates, one had mutation in codon 50 (substitution of phenylalanine with leucine) and the other had mutation in codon 44 (substitution of methionine with isoleucine, which was a novel mutation in this position). From these results, we identified the mutation in the H+-ATPase subunit c gene (atpC) of S. pneumoniae, which was not recognized earlier, and determined that Opt resistance among Japanese pneumococcal isolates is not related to a specific pneumococcal serotype or antimicrobial resistance profile. Furthermore, the results indicate that when a-hemolytic streptococci resistant to optochin are isolated from patients with invasive infectious diseases, such as meningitis and pneumonia, we should perform additional examinations such as bile solubility tests or PCR assays before confirming isolates as viridans streptococci. This is the first report of the characterization of Optr S. pneumoniae in Japan.
1. Introduction
Streptococcus pneumoniae colonize the nasopharynx in 20% - 40% of children and 5% - 10% of adults at any time, and cause serious infectious diseases, such as pneumonia, septicemia, meningitis, and otitis media [1]. Five phenotypic characteristics are classically used in the clinical laboratory for the presumptive identification of S. pneumoniae: Gram stain morphology, colony morphology, type of hemolysis, optochin susceptibility, and agglutination with anti-pneumococcal polysaccharide capsule antibodies. Accurate identification is important for ensuring the correct diagnosis and treatment of patients because of the increasing frequency of resistance to penicillin and other antibiotic agents.
The optochin susceptibility test is one of the most important methods for differentiating S. pneumoniae from other a-hemolytic streptococci. Optochin (ethylhydrocupreine hydrochloride) is a quinine analog. It was introduced as a therapeutic agent for treatment of lobar pneumonia in early 20th century. However, its use resulted in severe side effects and a study reported that 4.5% of patients treated with optochin experienced loss of vision [2]. Thus, it was stopped being used as a therapeutic agent. Later, in 1915, optochin was found to be useful for differentiating S. pneumoniae from other a-hemolytic streptococci [3]. The optochin susceptibility test was found to be highly satisfactory and less timeconsuming for the identification of S. pneumoniae compared to bile solubility test; hence, it was adopted in clinical laboratories in 1955 [4]. In 1987, Kontiainen and Sivonen [5] first reported the identification of two clinical isolates of optochin-resistant (Optr) S. pneumoniae in blood samples taken from a 74-year-old man with pneumonia and liver cirrhosis and an 8-month-old child with sepsis from otitis media. Since then, the emergence of Optr S. pneumoniae has been reported from the United States [6-8], Israel [9], Portugal [10,11], Brazil [12], and Argentina [13]. In addition, mutations of the gene encoding subunit c of the Fo complex of transmembrane H+-ATPase were reported to be responsible for optochin resistance [14]. There have been no reports of Optr S. pneumoniae in Japan.
The purpose of this study is to investigate the isolation frequency of Optr S. pneumoniae in Japan, report the biochemical and genetic characteristics of the isolates, and alert clinical microbiologists of the presence of these strains in the community.
2. Materials and Methods
2.1. Bacterial Strains
A total of 1032 presumptive S. pneumoniae isolates were collected by the North Fukuoka Infectious Diseases Working Group (NFIDWG, Fukuoka, Japan). These clinical isolates were recovered mainly from the nasopharynx and sputum samples obtained from 138 medical clinics and hospitals participating in NFIDWG. Each isolate was confirmed to be S. pneumoniae based on its Gram stain morphology, colony morphology, type of hemolysis, optochin susceptibility, and agglutination with anti-pneumococcal polysaccharide capsule antibodies. In cases where optochin resistance was suspected, we conducted further examinations such as the bile solubility test, determination of the presence of the major autolysin gene (lytA) by PCR [15], determining bacterial profiles by matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS), and sequencing of the 16S rRNA genes (1480 to 1485 bases). S. pneumoniae R6 ATCC BAA-255 (uncapsulated derivate of D39) was used as the reference strain.
2.2. Optochin Susceptibility
Optochin disks (5 μg each; Eiken Co., Ltd., Tokyo, Japan) were placed on 5% sheep blood agar plates (Eiken) streaked with the test isolate. The diameter of inhibition zones around the disk was measured after 18 - 24 h incubation at 35˚C - 37˚C in a 5% CO2 atmosphere. Bacterial isolates with a diameter measuring ≥13 mm were tentatively identified as optochin-sensitive, while isolates with a diameter < 13 mm were identified as optochinresistant. The MICs of optochin (Sigma Co. LLC, St. Louis, MO, USA) for 1031 isolates were determined using the plate dilution method with Mueller-Hinton agar (Difco Laboratories, Detroit, MI, USA) containing 5% defibrinated sheep blood (Kohjin Bio Co., Ltd., Saitama, Japan) and an inoculum size of 104 CFU of bacteria. Cell growth was evaluated after incubation for 24 h at 37˚C in a 5% CO2 atmosphere. Bacterial isolates were considered optochin resistant when MICs were ≥4 μg/ml, based on the result of our present study (Figure 1) and a previous report by Pikis et al. [8].
2.3. Bile Solubility, Latex Agglutination, and Capsular Serotyping
Bile solubility and latex agglutination tests were performed using the Slidex Pneumo-Kit (Nippon bioMérieux Co., Ltd., Tokyo, Japan) based on the methods described by Whatmore et al. [16]. Capsular serotyping (the Neufeld Quellung test) was performed using each type or group serum (Statens Serum Institut, Copenhagen, Denmark) at the National Institute of Infectious Diseases, Tokyo, Japan.
2.4. Biotyping
Biotyping was performed using API 20 Strep V7.0 (Nippon bioMérieux) and an automated VITEK 2 Gram Positive Identification (GPI) Card (Nippon bioMérieux) at the Central and Clinical Laboratories in Saga University Hospital by medical technologists who are specialists in clinical microbiology. Cultures were grown anaerobically on 5% sheep blood agar (Eiken) at 35˚C - 37˚C for 22 - 26 h and suspensions were prepared for API 20 Strep V7.0 according to the manufacturer’s instructions. The result was determined by matching data with an API 20 Strep profile list. For preparing the GPI Card, colonies were picked and suspended in a 3.0-ml-sterile salt solution (pH 4.5 - 7.0), which was equivalent to a McFarland’s 0.50 - 0.63 standard according to the manufac-
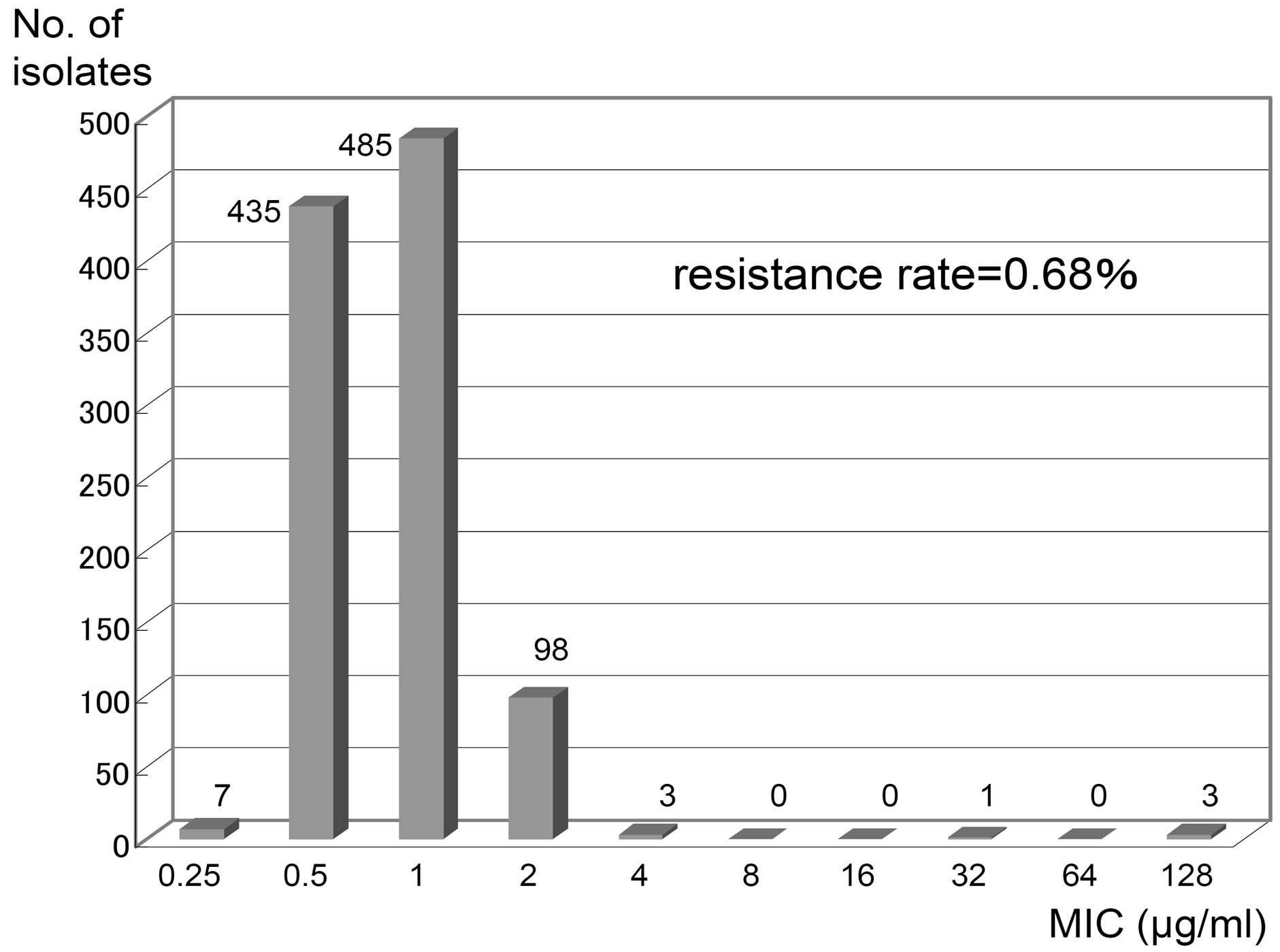
Figure 1. Distribution of MICs of Optochin in Streptococcus pneumoniae clinical isolates (1032 strains). The MICs are determined by the plate dilution method.
turer’s instructions.
2.5. PCR for the Major Autolysin Gene (lytA)
The lytA gene was detected by PCR using primers described by Ubukata et al. [15]. Using a Biometra T Gradient Thermocycler (Biometra GmbH, Goettingen, Germany), thirty cycles of DNA amplification were performed as follows: denaturation at 94˚C for 15 s, annealing at 50˚C for 30 s, and extension at 72˚C for 30 s. The presence of an amplified 273-bp sequence of the autolysin gene indicated the presence of the lytA gene. The DNA size marker was 100 Base-Pair Ladder DNA (Pharmacia Biotech Co., Ltd., Tokyo, Japan).
2.6. Matrix-Assisted Laser Desorption Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF MS) Identification
Identification of isolates by MALDI-TOF MS was performed on a Microflex LT instrument (Bruker Daltonics GmbH, Leipzig, Germany) with FlexControl (version 3.0) software (Bruker Daltonics) for the automatic acquisition of mass spectra in the linear positive mode within a range of 2 - 20 kDa. Colonies were examined by both direct deposition on MSP 96 target plates (Bruker Daltonics) and after a formic acid-acetonitrile extraction step according to the manufacturer’s instructions. According to the criteria proposed by the manufacturer, a result was considered valid (accurate identification to the species level) when the score was >2.0.
2.7. Antibiotic Susceptibility
The MICs to 12 antimicrobial agents were determined using the microdilution broth method following the Clinical and Laboratory Standards Institute (CLSI) guidelines. The antibiotics used in this test were as follows: benzylpenicillin (PCG), ampicillin (ABPC), cefazolin (CEZ), cefotiam (CTM), cefotaxime (CTX), cefpodoxime (CPDX), cefditoren (CDTR), panipenem (PAPM), minocycline (MINO), erythromycin (EM), clindamycin (CLDM), and levofloxacin (LVFX).
2.8. Sequencing of 16S rRNA Gene
The 16S rDNA (~1.5 kb) was amplified by PCR using the primers 8UA (5’-AGAGTTTGATCMTGGCTCAG- 3’) and 1485B (5’-TACGGTTACCTTGTTACGAC-3’) [17]. The purified PCR product was sequenced directly on both strands using a 3130xl Genetic Analyzer (Applied Biosystems, Foster City, CA, USA) and a BigDye Terminator Cycle Sequencing Kit v3.1 (Applied Biosystems). The primers 519A (5’-CAGCMGCCGCGGTAA- 3’), 519B (5’-ATTACCGCGGCRGCTG-3’), 774A (5’- GTAGTCCACGCTGTAAACGATG-3’), 774B (5’-CATCGTTTACAGCGTGGACTAC-3’), and 907B (5’-CCGTCAATTCMTTTRAGTTT-3’) were used as internal primers for sequencing. Homology search with the 16S rRNA gene sequences were performed against sequences registered in GenBank/EMBL/DDBJ using a basic local alignment search tool (BLAST).
2.9. Cloning and DNA Sequence Analysis of H+-ATPase Subunit c Gene
The H+-ATPase subunit c gene (atpC) of S. pneumoniae was amplified by PCR using primers (sense primer: 136 5’-TAGCGGTTAAAAGTTGACAA-3’; antisense primer: 437 5’-CCCTTTTCTTCTCGTTCC-3’) described by Cogné et al. [18]. After initial denaturation at 95˚C for 2 min, 25 cycles of DNA amplification were performed as follows: denaturation at 95˚C for 1 min, annealing at 54˚C for 2 min, extension at 72˚C for 2 min 30 s, and a final extension at 72˚C for 7 min 30 s. The expected 302-bp fragment was purified using PCR purification columns (GenElute Minus EtBr Spin Columns; Sigma Chemical Co., St Louis, MO, USA) and cloned using a TOPO TA Cloning Kit (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Transformation was performed using competent E. coli TOP10 cells provided by the manufacturer. A total of three to five white colonies were randomly selected from each clone library for sequence analysis. To prepare a template for sequence analysis, a partial fragment of the cloning vector (PCR II) containing an inserted PCR product was amplified using M13Forward (5’-GTAAAACGACGGCCAG-3’), M13Reverse (5’-CAGGAAACAGCTATGAC-3’), and AmpliTaq Gold DNA polymerase. Primers and dNTP were eliminated from the PCR mixture using an ExoSAP-IT Kit (USB, Cleveland, OH, USA) according to the manufacturer’s instructions and a 1 ml aliquot was used as a template for the sequencing reaction. Sequencing reactions were performed using the M13 primers and a BigDye Terminator Cycle Sequencing Kit v3.1 (Applied Biosystems). The nucleic acid sequences were determined using a 3130xl Genetic Analyzer (Applied Biosystems).
2.10. Genetic Transformation
Transformation in S. pneumoniae R6 was performed as described by Muñoz et al. [19]. S. pneumoniae R6 was grown in C medium plus yeast extract (C + Y) [20] to the late exponential phase and frozen at −80˚C after the addition of glycerol to 15%. Frozen stock (200 µl) was added to 4 ml of C + Y medium and incubated for about 2 h to reach competence for transformation. The competent S. pneumoniae R6 cells were diluted 10-fold in C + Y medium and a cloned PCR fragment (atpC gene) of S. pneumoniae SPJ661 was added at final concentrations of 0.1 mg/ml to 500 ml of the diluted cultures. These were then incubated at 30˚C for 1 h and at 37˚C for at least 2 h to allow expression of optochin resistance. Samples were then plated in 100 ml volumes on Mueller-Hinton agar (Difco Laboratories) containing 5% defibrinated sheep blood (Kohjin Bio) and 2 mg/ml optochin (Sigma).
2.11. Nucleotide Sequence Accession Numbers
Nucleotide sequence data of partial sequences of atpC genes were deposited in the DDBJ database under the accession numbers AB569578 to AB569584.
3. Results
3.1. Isolation Frequency and Biochemical Characteristics of Optochin-Resistant S. pneumoniae
A total of 1032 strains conclusive or presumptive as S. pneumoniae were collected by NFIDWG from patients with invasive diseases, such as pneumonia, septicemia, meningitis, and otitis media. Of these, 1025 isolates (99.32%) exhibited an optochin MIC of ≤2 mg/ml and seven isolates (0.68%) exhibited an MIC of ≥4 mg/ml (three isolates, 4 mg/ml; 1 isolate, 32 mg/ml; three isolates, ≥128 mg/ml) using the plate dilution method (Figure 1, Table 1). Table 1 shows that three of the seven optochin-resistant strains were isolated from the nasal discharges of children, while four strains were isolated from the sputum of elderly individual persons. As shown in Table 2, all isolates presented typical Gram staining characteristics, a-hemolysis on blood agar plates, bile solubility, and agglutination reactions with sera targeting pneumococcal capsular polysaccharides. In addition, all isolates contained the lytA gene and exhibited autolysis.
However, they were not confirmed as S. pneumoniae using API Strep 20 V.7.0 because they had biochemical alterations with regards to optochin resistance, a loss of inulin and starch utilization, and a loss of arginine hydrolysis and pyrrolidone arylamidase (Table 3). Table 2 shows that they were identified as S. pneumoniae using the extraction method based on MALDI-TOF MS analysis. Mass spectra were not obtained for strain SPJ1298 using the direct method because it produced mucoid colonies. Mass spectra were also not obtained for another three isolates probably because of autolysis, when the colonies were analyzed on day 1 after being cultured on blood agar plates (Table 2). Isolates belonged to different serotypes (Table 2) and exhibited different antimicrobial resistance profiles (Table 1). Although all Optr isolates were susceptible to PCG and LVFX, six of the seven isolates were resistant to EM (Table 1). In addition, five isolates were CPDX intermediates or CPDX resistant, and four isolates were resistant to CLDM (Table 1). Finally, all isolates were confirmed to be S. pneumoniae by the 16S rRNA gene analysis (Table 2).
3.2. Genetic Characteristics of Optochin-Resistant S. pneumoniae
Table 4 shows that all the seven Optr isolates had mutations in the nucleotide sequence coding for subunit c of F0F1 ATPase. Three isolates (SPJ48, 492, and 1298) had mutations in codon 48 (GTT to TTT, deduced amino acid substitution of valine with phenylalanine) and two isolates (SPJ743 and 1331) had mutations in codon 49 (GCC to ACC or TCC, substitution of alanine with threonine or serine). Of the remaining two isolates, one (SPJ246) had mutation in codon 50 (TTT to CTT, substitution of phenylalanine with leucine) and the other
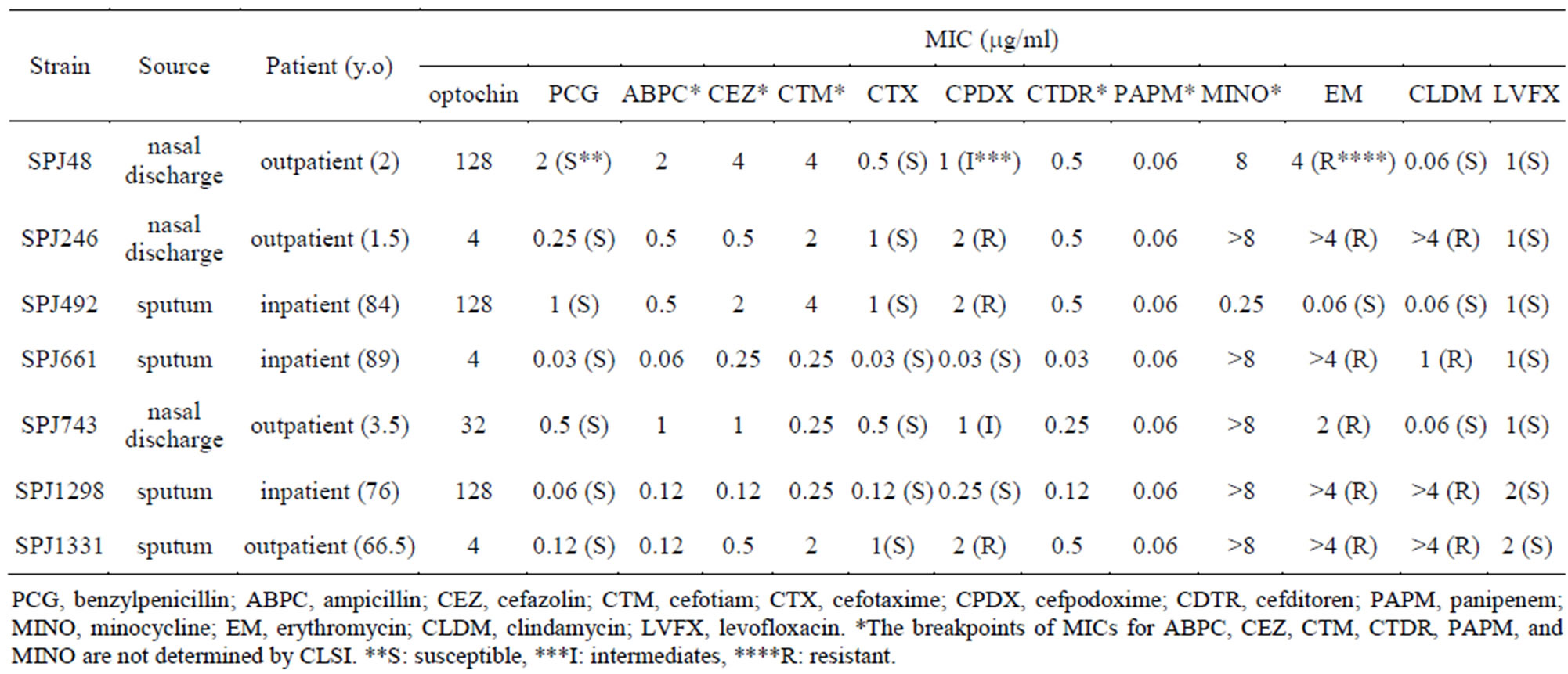
Table 1. Characteristics of optochin-resistant Streptococcus pneumoniae clinical isolates.
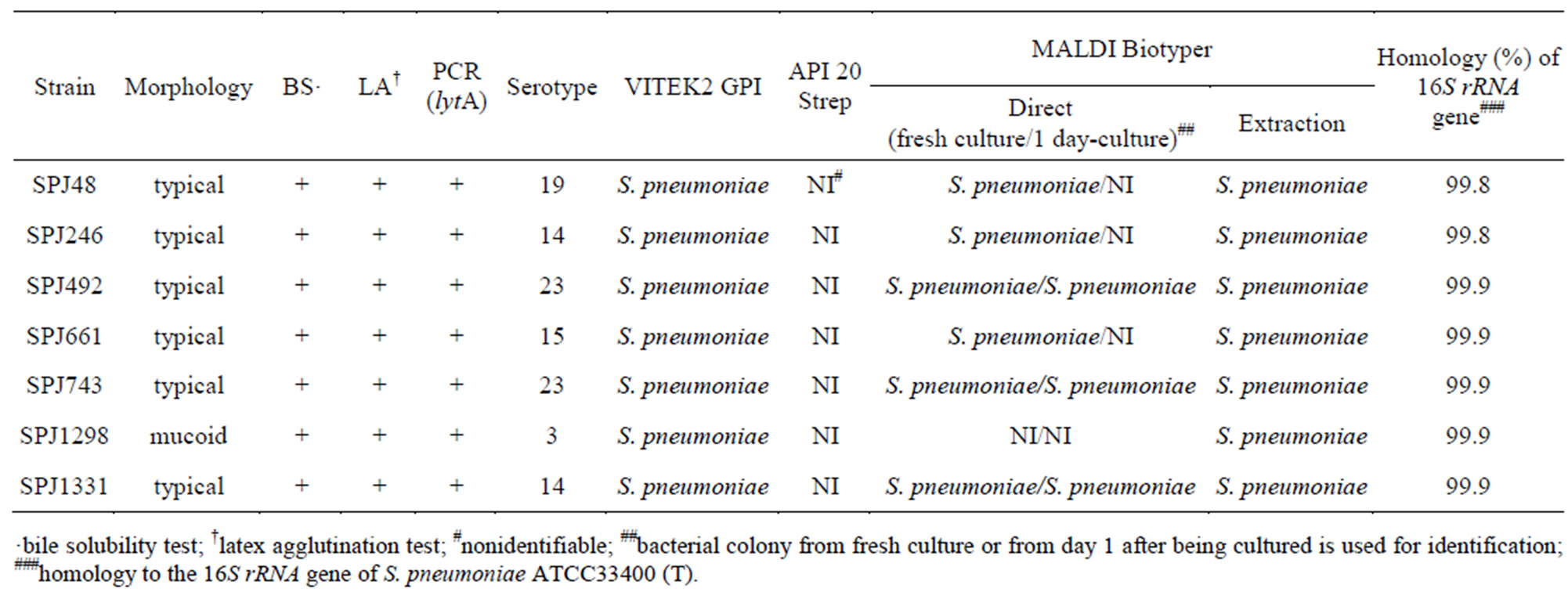
Table 2. Characteristics and identification of optochin-resistant Streptococcus pneumoniae clinical isolates.
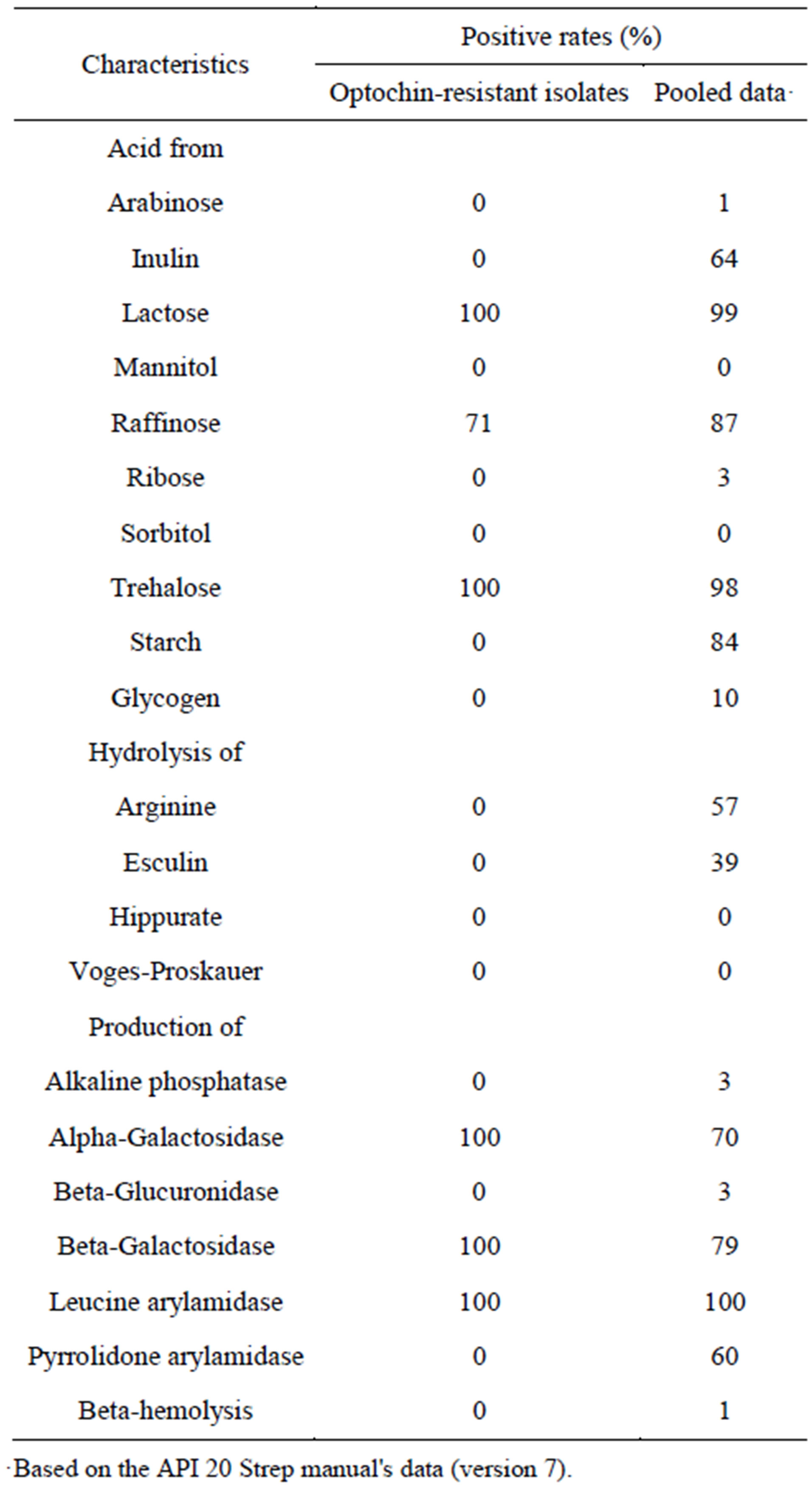
Table 3. Biochemical characteristics of optochin-resistant Streptococcus pneumoniae.
(SPJ661) had mutation in codon 44 (ATG to ATT, substitution of methionine with isoleucine, which was a novel type of mutation in this position). To identify the novel mutation (M44I) responsible for optochin resistance, strain R6 was transformed with cloned PCR products containing the M44I mutation. Two transformants (strains R6/R-7 and R6/4-16) were selected and tested for susceptibility to optochin. The transformants did not produce adequate inhibition zones around the optochin disks. Furthermore, when the optochin MICs for the transformants were tested, they were the same as those for the original strain SPJ661 (Table 4). Sequence analysis of the transformants confirmed the expected amino acid changes (Table 4).
4. Discussion
S. pneumoniae is a major pathogen that causes fatal infectious diseases such as pneumonia, septicemia, and meningitis, particularly in children and elderly individuals. The increasing frequency of resistance to antibiotic agents necessitates accurate identification for ensuring the correct diagnosis and treatment of patients, especially in Japan where approximately 35% of S. pneumoniae strains are penicillin intermediates (PISP) or penicillin resistant (PRSP), and 60% - 70% of the strains are macrolide resistant [21]. The optochin susceptibility test is a key method for differentiating S. pneumoniae from other a-hemolytic streptococci, and most clinical microbiology laboratories depend on this test for differentiating streptococci. However, Optr S. pneumoniae have been reported in the last two decades [6-13]. Both information of the isolation frequency of Optr S. pneumoniae in Japan and knowledge about the biochemical and genetic characteristics of the Optr isolates are required for accurate identification of S. pneumoniae, leading to correct diagnosis and treatment of patients.

Table 4. Comparisons of ATPase c-subunit amino acid sequences between optochin-resistant Streptococcus pneumoniae and optochin-sensitive R6 strain (ATCCBAA-255).
In this study, seven (0.68%) out of 1032 S. pneumoniae isolates collected by NFIDWG were Optr S. pneumoniae (Figure 1). The isolation frequency was similar to those found in the United States [6-8], Israel [9], Portugal [10,11], Brazil [12], and Argentina [13] (0.58% to 2.1%). Optochin resistance among Japanese pneumococcal isolates was not related to a specific pneumococcal serotype or antimicrobial resistance profile (Tables 1 and 2). In addition, Table 3 shows that the biochemical characteristics (loss of inulin and starch utilization and loss of arginine hydrolysis) of these Japanese Optr isolates were similar to those of the first clinical Optr isolates reported from Finland in 1987 [5]. These results suggest that the emergence of Optr isolates may not be by clonal expansion, but may be attributable to spontaneous mutations that are not caused by exposure to any antimicrobial agent. It was also shown that API 20 Strep is not useful for species identification of Optr S. pneumoniae because they had the biochemical alterations.
It was reported that point mutations in the atpC genes, which encode subunits of F0F1-ATPase, conferred optochin resistance on S. pneumoniae [8,12,14,18]. On the other hand, there are no reports about mutations in the atpC genes among optochin-sensitive S. pneumoniae [8, 13,14]. H+-ATPase is a membrane-bound multimeric enzyme complex responsible for proton translocation across plasma membranes. This enzyme is an F0F1 class ion transport ATPase. The F0 sector of the complex comprises membrane-embedded protein subunits that translocate hydrogen ions, while the F1 sector of the complex contains the catalytic subunit for ATP hydrolysis and synthesis. Certain amino acid residues in the transmembrane domains are essential for H+ translocation across the plasma membrane [8,22]. The molecular mechanism determining how optochin interacts with the F0 portion of ATPase remains unresolved, but the protontranslocating subunits of ATPase may be disrupted in the presence of optochin, resulting in proton pump failure and S. pneumoniae cell death. Amino acid changes in the subunit c in Optr isolates may prevent optochin from disrupting the proton transport pathway. Only 21 Optr clinical strains are known to have been characterized at the molecular level and 10 mutations in the atpC gene have been reported [13]. The most frequent modifications in our Optr clinical strains (Table 4) were found at position 48 in the subunit c where valine was substituted with phenylalanine (V48F), as also reported by Fenoll et al. [14]. This mutation may be involved in the high levels of optochin-resistance (strains SPJ48, 492, and 1298 in Table 4). The second most common modification was found at position 49 where alanine was substituted with serine (A49S) or threonine (A49T), which was also reported previously [8,12-14]. In both cases, the nonpolar hydrophobic alanine was replaced by a polar uncharged amino acid (serine or threonine) in the a-helix 2 domain. This polarity change may interfere with the binding of optochin to subunit c. We identified a novel mutation, M44I, in the same domain.
This is the first report of the characterization of Optr S. pneumoniae in Japan. Cortes et al. [13] reported that Optr strains maintain their virulence as well as optochin-susceptible S. pneumoniae. Nunes et al. [11] reported a prevalence of approximately 2.1% from a total of 1973 pneumococcal strains isolated during a period of 6 years in Portugal. Thus, when a-hemolytic streptococci resistant to optochin are isolated from patients with invasive infectious diseases, such as meningitis and pneumonia, we should perform additional examinations such as bile solubility tests or PCR assays before confirming isolates as viridans streptococci. Accurate identification and antimicrobial susceptibility testing are crucial for correct diagnosis and treatment of patients.
5. Acknowledgements
We acknowledge all members of the North Fukuoka Infectious Diseases Working Group for collecting Streptococcus pneumoniae clinical isolates. We thank Tadao Tanabe (University of Occupational and Environmental Health), Zenzo Nagasawa (Saga University Hospital), and Koji Kusaba (Saga University Hospital) for technical assistance. We also thank Kimiko Ubukata (Kitasato University) and Akihito Wada (National Institute of Infectious Diseases, Japan) for advice on Streptococcus pneumoniae transformation and for performing capsular serotyping, respectively.
REFERENCES
- D. A Musher, “Streptococcus pneumoniae,” In: G. L. Mandell, J. E. Bennett and R. Dolin, Eds., Mandell, Douglas and Bennett’s Principles and Practice of Infectious Diseases, 6th Edition, Churchill Livingstone, Philadelphia, 2005, pp. 2392-2411.
- K. P. Klugman, “Pneumococcal resistance to antibiotics,” Clinical Microbiology Reviews, Vol. 3, No. 2, 1990, p. 171.
- K. Z. Rochs, “Zur Differentialdiagnose der Streptokokken und Pneumokokken,” Virchows Archiv, Vol. 220, No. 3, 1915, pp. 327-346. dio:10.1007/BF01949112
- E. F. Bowers and L. R. Jeffries, “Optochin in the identification of Streptococcus pneumoniae,” Journal of Clinical Pathology. Vol. 8, 1955, pp. 58-60. dio:10.1136/jcp.8.1.58
- S. Kontiainen and A. Sivonen, “Optochin resistance in Streptococcus pneumoniae strains isolated from blood and middle ear fluid,” European journal of clinical microbiology, Vol. 6, No. 4, 1987, pp. 422-424. dio:10.1007/BF02013101
- A. P. Borek, D. C. Dressel, J. Hussong and L. R. Peterson, “Evolving clinical problems with Streptococcus pneumoniae: increasing resistance to antimicrobial agents, and failure of traditional optochin identification in Chicago, Illinois, between 1993 and 1996,” Diagnostic Microbiology and Infectious Disease, Vol. 29, No. 4, 1997, pp. 209-214. dio:10.1016/S0732-8893(97)00141-7
- A. Pikis, W. J. Rodriguez, J. M. Campos and J. M. Keith, “Optochin-resistant Streptococcus pneumoniae,” Pediatric Research, Vol. 41, 1997, p. 128. dio:10.1203/00006450-199704001-00775
- A. Pikis, J. M. Campos, W. J. Rodriguez and J. M. Keith, “Optochin resistance in Streptococcus pneumoniae: mechanism, significance, and clinical implications,” Journal of Infectious Diseases, Vol. 184, No. 5, 2001, pp. 582-590. dio:10.1086/322803
- F. Lejbkowicz, M. Goldstein, N. Hashman and L. Cohn, “Optochin-resistant Streptococcus pneumoniae isolated from a blood specimen,” Clinical Microbiology Newsletter, Vol. 21, No. 9, 1999, pp. 72-73. dio:10.1016/S0196-4399(00)80048-3
- S. I. Aguiar, M. J. Frias, L. Santos, J. Melo-Cristino and M. Ramirez, “Emergence of optochin resistance among Streptococcus pneumoniae in Portugal,” Microbial Drug Resistance, Vol. 12, No. 4, 2006, pp. 239-245. dio:10.1089/mdr.2006.12.239
- S. Nunes, R. Sá-Leão and H. de Lencastre, “Optochin resistance among Streptococcus pneumoniae strains colonizing healthy children in Portugal,” Journal of Clinical Microbiology, Vol. 46, No. 1, 2008, pp. 321-324. dio:10.1128/JCM.02097-07
- C. A. Dias, G. Agnes, A. P. Frazzon, F. D. Kruger, P. A. d'Azevedo, et al., “Diversity of mutations in the atpC gene coding for the c subunit of F0F1 ATPase in clinical isolates of optochin-resistant Streptococcus pneumoniae from Brazil,” Journal of Clinical Microbiology, Vol. 45, No. 9, 2007, pp. 3065-3067. dio:10.1128/JCM.00891-07
- P. R. Cortes, A. G. Orio, M. Regueira, G. E. Piñas and J. Echenipue, “Characterization of in vitro-generated and clinical optochin-resistant strains of Streptococcus pneumoniae isolated from Argentina,” Journal of Clinical Microbiology, Vol. 46, No. 6, 2008, pp. 1930-1934. dio:10.1128/JCM.02318-07
- A. Fenoll, R. Muñoz, E. García and A. G. de la Campa, “Molecular basis of the optochin-sensitive phenotype of pneumococcus: characterization of the genes encoding the F0 complex of the Streptococcus pneumoniae and Streptococcus oralis H+-ATPases,” Molecular Microbiology, Vol 12, No. 4, 1994, pp. 587-598. dio:10.1111/j.1365-2958.1994.tb01045.x
- K. Ubukata, Y. Asahi, A. Yamane and M. Konno, “Combinational detection of autolysin and penicillin-binding protein 2B genes of Streptococcus pneumoniae by PCR,” Journal of Clinical Microbiology, Vol. 34, No. 3, 1996, pp. 592-596.
- A. M. Whatmore, A. Efstratiou, A. P. Pickerill, K. Broughton, G. Woodard, D. Sturgeon, et al., “Genetic relationships between clinical isolates of Streptococcus pneumoniae, Streptococcus oralis, and Streptococcus mitis: characterization of ‘Atypical’ pneumococci and organisms allied to S. mitis harboring S. pneumoniae virulence factor-encoding genes,” Infection and Immunity, Vol. 68, No. 3, 2000, pp. 1374-1382. dio:10.1128/IAI.68.3.1374-1382.2000
- D. J. Lane, “16S/23S rRNA sequencing,” In: E. Stackebrandt and M. Goodfellow, Eds., Nucleic Acid Techniques in Bacterial Systematics, Wiley, Chichester, 1991, pp. 115-175.
- N. Cogné, J. P. Claverys, F. Denis and C. Martin, “A novel mutation in the α-helix 1 of the c subunit of the F1/F0 ATPase responsible for optochin resistance of a Streptococcus pneumoniae clinical isolate,” Diagnostic Microbiology and Infectious Disease, Vol. 38, No. 2, 2000, pp. 119-121.
- R. Muñoz, C. G. Dowson, M. Daniels, T. J. Coffey, C. Martin, R. Hakenbeck and B. G. Spratt, “Genetics of resistance to third-generation cephalosporins in clinical isolates of Streptococcus pneumoniae,” Molecular Microbiology, Vol. 6, No. 17, 1992, pp. 2461-2465.
- A. Tomas and R. Hotchkiss, “Regulation of the transformability of pneumococcal cultures by macromolecular cell products,” Proceedings of the National Academy of Sciences of the United States of America, Vol. 51, No. 3, 1964, pp. 480-487. dio:10.1073/pnas.51.3.480
- Y. Niki, H. Hanaki, T. Matsumoto, M. Yagisawa, S. Kohno, N. Aoki, et al., “Nationwide surveillance of bacterial respiratory pathogens conducted by the Japanese Society of Chemotherapy in 2007: general view of the pathogens’ antibacterial susceptibility,” Journal of Infection and Chemotherapy, Vol. 15, No. 3, 2009, pp. 156-167. dio:10.1007/s10156-009-0674-z
- A. E. Senior, “The proton-translocating ATPase of Escherichia coli,” Annual review of biophysics and biophysical chemistry, Vol. 19, 1990, pp. 7-41. dio:10.1146/annurev.bb.19.060190.000255
NOTES
*Corresponding author.