CellBio
Vol.1 No.2(2012), Article ID:25503,4 pages DOI:10.4236/cellbio.2012.12004
Calcium Integrin Binding Protein Associates with Integrins αVβ3 and αIIbβ3 Independent of β3 Activation Motifs
Department of Cell and Developmental Biology, State University of New York Upstate Medical University, Syracuse, USA
Email: *blystons@mail.upstate.edu
Received September 11, 2012; revised October 15, 2012; accepted October 25, 2012
Keywords: Leukocyte; Integrin; Cytoskeleton; Hematopoietic; Activation; Signaling
ABSTRACT
The Calcium Integrin Binding protein (CIB) has been identified as interacting specifically with the cytoplasmic tail of the integrin αIIb domain to induce receptor activation and integrin αIIbβ3 mediated cell adhesion to extracellular proteins. In K562 cells stably expressing mutated integrin αVβ3, or chimeric αVβ3 carrying αIIb cytoplasmic tail, we report that the interaction of CIB with β3 integrins is not αIIbβ3 specific but binds αIIb as well as αV cytoplasmic tail domains. A double mutation of two proline residues to alanine residues in the αIIb cytoplasmic domain, previously shown to disturb its conformation, inhibits chimeric αV/αIIbβ3-CIB interaction. This demonstrates that αIIb cytoplasmic domain loop-like conformation is required for interaction with CIB. Moreover, mutations of β3 cytoplasmic domain residues Tyr-747 and/or Tyr-759 to phenylalanine residues (Y747F, Y759F, and Y747,759F) as well as residues Ser-752 to proline or alanine (S752P and S752A), do not affect the αIIbβ3 or αVβ3 interaction with CIB. Since tyrosine residues Tyr-747 and/or Tyr-759 are the sites of tyrosine phosphorylation of β3 subunit, these results suggest that the β3 integrin-CIB interaction occurs through a β3-phosphorylation independent mechanism. Likewise, ablation of conformation-dependent affinity change in β3 Ser752Pro mutation had no affect on CIB-β3 interaction. In summary, our results demonstrate that the αIIb-subunit integrin and CIB interaction is non-exclusive and requires the loop-like αIIb-cytoplasmic domain conformation. An interaction of CIB with αV-containing integrins provides an additional role for this molecule in keeping with its expression outside of platelets.
1. Introduction
The Calcium Integrin Binding (CIB) protein is a 22-kDa intracellular member of the family of regulatory calciumbinding proteins that includes calmodulin, calcineurin B and recoverin. It was originally detected through the use of yeast two-hybrid systems as interacting specifically with the integrin αIIb-cytoplasmic tail in the presence of calcium [1]. Various methods have subsequently confirmed this interaction in vitro [2-4] and in vivo [5]. This CIB-αIIb interaction has been shown to promote the affinity of the αIIbβ3 integrin to fibrinogen [5]. Moreover, it has been shown that the association of CIB with integrin αIIbβ3 during outside-in signaling is required for platelet spreading on fibrinogen [6] through regulation of focal adhesion kinase (FAK) activation [7]. Furthermore, it was reported that CIB binds to the GTPase Rac3 and that co-expression of active Rac3 and CIB in Chinese hamster ovary (CHO) cells also expressing αIIbβ3 results in enhanced αIIbβ3-mediated cell adhesion and spreading on fibrinogen [8]. In addition to its interaction with the αIIbβ3 integrin, FAK and Rac3 regulatory functions, CIB may have other functions because it interacts with several diverse cytosolic proteins involved in neuronal function (FnK and Snk kinase) [9] and Alzheimer’s disease (Presenilin 2) [10].
A previous model based on homology modeling and nuclear magnetic resonance (NMR) studies suggests that the N-terminal domain of CIB containing positively charged amino residues could interact with the acidic C-terminal tail of αIIb whereas the C-terminal domain of CIB could bind the αIIb membrane proximal α-helix structure carrying the highly conserved GFFKR motif [4]. In yeast two-hybrid assays using fragments of CIB as bait or in intrinsic tryptophan fluorescence assays, the C-terminus of CIB serves as a site of interaction of CIB with the αIIb cytoplasmic tail [3]. This study localized the CIB binding site on αIIb to within a 15-amino acid residue stretch including residues within the membrane-splanning region as well as several residues within the membrane proximal region of the αIIb cytoplasmic domain (from L983 to R997). Because the membrane proximal GFFKR motif of αIIb is highly conserved among α integrin subunits, the specificity of the CIB-αIIb interaction suggests the presence of a supplementary determinant involved in this exclusive relationship. The αIIb and β3 subunit cytoplasmic domains form α-helices and exhibit two-stranded coiled-coil structures in which there are several electrostatic and hydrophobic interactions [11,12]. The inhibition of these interactions is emerging as the molecular mechanism leading to the receptor activation and, through competition with αIIb subunit in binding the β3 subunit; CIB may contribute to this process. The αIIb subunit cytoplasmic domain is organized in a “closed” conformation where the highly conserved N-terminal and membrane proximal GFFKR motif form an α-helix followed by a turn, and the acidic C-terminal loop interacts with the N-terminal helix. The substitutions of proline residues 998 and 999 with alanine residues generate an “open” conformation where the interactions between the N-terminal helix and C-terminal moiety are abolished, inducing receptor activation [13].
In the present study using K562 cells expressing mutated integrins, we report that, in addition to the N-terminal membrane proximal GFFKR motif, the acidic C-terminal of the αIIb subunit cytoplasmic domain as well as the αIIb “closed” conformation are important for the αIIb- CIB interaction. Furthermore, we show for the first time that CIB also interacts with the αV cytoplasmic domain. Moreover, mutations of tyrosine or serine residues on the β3 subunit cytoplasmic domain, previously shown to prevent β3 tyrosine phosphorylation or receptor activation, failed to prevent interaction with CIB. Together these data indicate that in K562 cells, in addition to the N-terminal GFFKR motif, the binding of CIB to the cytoplasmic domain is αIIb cytoplasmic domain conformation dependent and is not αIIbβ3 specific, providing a basis for understanding CIB-integrin interaction in vivo and providing an additional role for CIB in non-platelet cell types.
2. Materials and Methods
2.1. Construction of Chimeric and Mutant Integrins
The human erythroleukemic cell line K562 transfected with cDNA encoding various mutated β3 or truncated αV integrin subunits were derived and maintained as previously described [14]. In addition, K562 cells were transfected with cDNA encoding full-length β3 and with cDNA encoding either truncated or double substituted proline residues to alanine residues of the αIIb cytoplasmic domain fused with the αV extracellular moiety. Transfected cells were selected for expression by fluorescent cell sorting flow cytometry using mAb to αV and β3 as previously described [14]. The recombinants αV/αIIb, in which αV cytoplasmic sequence has been replaced with the corresponding αIIb sequence were prepared in the following way: human wild type αIIb cDNA (a kind gift of Dr Ulhas P. Naik of the University of Delaware) was subjected to PCR using the IHY009 primer
5’CGAGGGGAGGTTACCGTGTGGACACAGCTG3’
introducing a new BstEII site (underlined) at the 2933 position and the IHY010 primer
5’GGCGGATCCATCACTCCCCCTCTTCATC3’
introducing a new Bam-HI site (underlined) at the 3210 position. After BstEII-BamHI digestion, the PCR generated product was ligated into pcDNA3.1 zeo (-) αV digested with BstEII-BamHI, creating pcDNA3.1 zeo (-) αV/CTαIIb construct. Similarly, using αIIb cDNA as template and the same forward and reverse primers;
5’CCTCTGGATCCTCTTCTTACAGGGGTGGC3’,
5’CATCTGGATCCAGGGGTGGCCGTTACCGCTTG3’
and
5’CCGGATCCGCTTGAAGAAGCCTTGACGTTCC3’,
5’GGCCGGATCCGCTTGAAGAATTAGACCTTCC3’,
we generated PCR products with new stop codons and BamHI sites at 3087 and 3096, 3078 and 3090, 3063 and 3077 positions respectively. As shown previously, using αIIb cDNA as template and IHY009 with 5’GCGGAACCGGGCAGCCCTGGAAGAAGATG3’
and IHY010 with
5’CATCTTCTTCCAGGGCTGCCCGGTTCCGC3’ in a first round PCR we made PCR products which were then used as a template for nested PCR using IHY009 and IHY010 primers. These final products were digested with BstEII and BamHI and ligated in to pcDNA3.1 zeo (-) αV/CTαIIb digested with the same enzymes. These products, with β3 cDNA, were stably expressed in K562 cells to generate KαV/αIIbWTβ3, KαV/αIIb999β3, KαV/αIIb995β3, KαV/αIIb990β3 and KαV/αIIbPP998/999AAβ3 mutants expressing αV/αIIbβ3 integrin receptors having the extracellular αV domain fused to the αIIb transmembrane and cytoplasmic lacking the 9, 13 or 18 last amino acids or with the proline residues 998 and 999 mutated to alanine residues respectively. The mutations were assessed and confirmed for each mutant by DNA sequence analysis of the recombinant cDNA using the forward IHY009 primer in automated DNA sequencing analysis ABI 3100 Genetic Analyzer (SUNY Upstate Medical University, NY) and are depicted in Figure 1.
2.2. Cell Transfection and Flow Cytometry Analysis
K562 cells, an erythroleukemic, promonocyte cell line were grown in Iscove’s Modified Eagle’s Medium (IMDM) plus 10% fetal calf serum (FCS). About 1 × 107 cells were washed twice with IMDM, resuspended in 500 μl of HEPES Buffered Saline containing 25 μg of each purified recombinant αV/αIIb subunit cDNA (in pcDNA3.1 expression vector carrying a zeocin resistance gene) and β3 integrin cDNA (in pRc/RSV expression vector carrying a geneticin resistance gene). After incubation on ice for 10 min, cells were electroporated twice at 225 volts, 500 μF capacitance and 500 ohms and then resuspended in 10 ml of IMDM plus 10% fetal calf and incubated for 24 hrs. The cells were transferred into 10 ml of the same medium supplemented with 1.2 μg/ml of geneticin, incubated for 10 days and then in the same medium described above and also containing 1mg/ml zeocin. Transfected cells and populations of transfectants expressing high levels of integrins were obtained by fluorescence cell sorting using a mouse monoclonal anti-β3 subunit antibody (mAb 7G2). The expression level of the recombinant receptors was maintained by periodic sorting with magnetic beads linked to an anti-β3 subunit (mAb AP3) and anti-αV subunit (mAb L230) and monitored by flow cytometry using the same antibody. Flow cytometry was carried out using a Coulter Epics XL flow cytometer (Coulter, Miami, FL).
2.3. Analysis of β3 Integrin-CIB Interaction
K562 cells (10 × 106) expressing equivalent amounts of transfected integrins were washed twice and resuspended in 300 μl of IMDM containing 100 μM sodium vanadate (Na3VO4) with 2.0 mM MnCl for 15 min and lysed in 500 μl phosphate buffered saline (PBS) containing 0.5% NP40, 200 mM CaCl2, 150 mM NaCl, 1.0 mM phenylmethylsulfonyl fluoride, 10 µg/ml aprotinin, 10 µg/ml leupeptin, and 2.0 mM Na3VO4. The lysates were centrifuged at 15,000 rpm for 10 minutes at 4˚C and then precleared with gelatin-Sepharose for 1 hour at 4˚C. Precleared lysates were then immunoprecipitated with goat anti-mouse-Sepharose beads (ICN, Cosa Mesa, CA) coated with either anti-β3 (mAb 7G2) and anti-αV (mAb 3F12) or anti-CIB (mAb UN2-NH) monoclonal antibodies for 4 h at 4ºC. Immunoprecipitates were washed and separated on 7.5% (for β3 detection) or 12% (for CIB detection) SDS-PAGE. Electrophoresed proteins were transferred to polyvinylidene difluoride membranes. Equivalence of immunoprecipitated β3-integrin or CIB was verified by probing immunoprecipitated products with anti-β3 (mAb 7G2) or anti-CIB (mAb UN2-NH) monoclonal antibodies and visualized by enhanced chemiluminescence (ECL, Amersham Biosciences) and compared with prestained molecular weight markers (Rainbow, Amersham Biosciences).
2.4. Antibodies and Reagents
The monoclonal anti-β3 integrin antibody 7G2 used in immunoprecipitation and β3 western blotting was a gift from Dr. Eric J. Brown of UCSF. Monoclonal antibodies AP3 (anti-β3), M148 (anti-αIIb) and L230 (anti-αV) have been used to assess the surface expression on transfected K562 cells as previously described [15]. The monoclonal anti-CIB antibody UN2-NH was a gift from Dr. Ulhas P. Naik of the University of Delaware. All other reagents unless otherwise noted were purchased from Sigma.

Figure 1. Schematic of chimeric, mutant, and truncated integrin construction. A comparison of the cytoplasmic domain sequences of αV and αIIb integrins is shown along with the sites of chimeric splicing, point mutations, and cytoplasmic truncations. cDNA modifications were performed as described in Methods and verified by DNA sequencing. A line indicates the conserved charged residue (K or R) of each integrin at the boundary of the transmembrane domain (TM), immediately preceding the cytoplasmic tail.
3. Results and Discussion
3.1. Expression of Mutant and Chimeric Integrins in K562 Cells
We have previously demonstrated that the tyrosine residues 747 and the 759 of the integrin β3 cytoplasmic domain as well as the integrin αV cytoplasmic tail are involved in the phosphorylation of β3 integrins [14,15]. Likewise, others have shown that serine residue 752 is involved in β3 integrin activation [16] and in β3 integrinmediated FAK phosphorylation [17]. Moreover, CIB is considered to bind specifically to the αIIbβ3 integrin through interaction with αIIb cytoplasmic domain [1]. β3 phosphorylation and the conformational change associated with increased affinity are critical to β3 function. Further, while CIB association is reportedly specific to αIIb, CIB expression is not restricted to platelets.
In this study we sought to establish the role of α-subunit structure and β3 activation motifs in CIB association with β3 integrins. To that end, K562 cells expressing stable recombinants of αV or αIIb wild type integrin subunits associated with β3 subunit bearing either phenylalanine (phe) substitutions of Tyr-747 (KαVβ3Y747F, KαIIbβ3Y747F), Tyr-759 (KαVβ3Y759F, KαIIbβ3Y759F), or both (KαVβ3-Y747/759F, KαIIbβ3Y747/759F), proline (pro) substitution of serine (ser) residue 752 (KαVβ3S752P, KαIIbβ3 S752A) or conservative alanine (ala) substitution of Ser-752 (KαVβ3S752A, KαIIbβ3
S752A). We also used K562 cells expressing stable recombinants of wild type β3 subunits associated with an αV subunit where the cytoplasmic domain has been deleted (KαVNTβ3) or fused with αIIb transmembrane and cytoplasmic lacking off the 9 (KαV/αIIb999β3), 13 (KαV/ αIIb995β3) or 18 (KαV/αIIb990β3) last amino acids or bearing pro residues 998 and 999 substitution to Ala residues (KαV/αIIbPP998/999AAβ3) as depicted in Figure 1. Surface expression of the β3 integrins was detected on cells transfected with cDNA encoding for mutated αV, αIIb or β3. After several rounds of cell sorting, the FACS profiles of recombinants or wild types were similar with the exception of the mutants carrying αV or αIIb subunits where the deletion includes the membrane-proximal GFFKR motif (Table 1). This suggests that there are no obvious differences in the expression stability of the wild-type and various mutants and also confirms the involvement of the membrane-proximal GFFKR motif in efficient cell surface expression which has been shown by several reports to prevent either the heterodimerization of the α and β subunits [18,19] or their cell surface expression [20]. Moreover, when expressed in lymphocytes [20] or K562 cells [21], the deletion of this motif does not induce constitutive activation of the receptor (data not shown) in contrast with previous data of the receptor expressed transiently in CHO cells [22-24]. This suggests that the mechanism of integrin activation is cell type dependent.

Table 1. Expression of integrins in K562 cells.
3.2. Interaction of CIB with αβ3-Integrin Cytoplasmic Domains
We previously showed that upon cell activation by a RGD peptide, the β3 integrin becomes phosphorylated at Tyr747 in β3 integrins associated either with αIIb or αV whereas the phosphorylation at Tyr759 residue occurs only when the β3 integrin is associated with αIIb suggesting a regulation of this critical biochemical event by the α subunits [15]. Manganese cation (Mn2+) can induce β3 integrin activation and tyrosine phosphorylation [14]. Previous reports showed that the mutation of Ser752Pro limits β3 integrin activation [16] and exposes a CIB binding site on αIIb cytoplasmic domain [2]. To determine the involvement of β3 cytoplasmic tyrosine and serine residues in the interaction of β3 integrins with CIB, we used CIB blotting to examine immunoprecipitates from Mn2+ treated K562 cells expressing either αV or αIIb wild type subunits associated with wild type β3 or β3 bearing Phe substitutions at Tyr747 (Y747F) and Tyr759 (Y759F) both individually and concurrently (Y747,759F) or Pro (S752P) and Ala (S752A) substitution at Ser752. We show that CIB co-immunoprecipitates with all mutated β3 subunits associated with αV or αIIb wild type (Figure 2). Therefore, in K562 cells, the CIB-β3 integrin interaction is not αIIbβ3 specific, suggesting the ability of the αV cytoplasmic domain to interact with CIB. To confirm this CIB-αV interaction, we immunoprecipitated lysate from Mn2+ treated K562 cells expressing αV in which the cytoplasmic domain had been deleted (KαVNTβ3) in comparison with cells expressing wild type αVβ3 (KαVβ3 WT) and αIIbβ3 (KαIIbβ3 WT) as control. As shown in Figure 3(a), the deletion of the αV cytoplasmic domain prevents the co-immunoprecipitation of CIB by anti-β3 and anti-αV antibodies, confirming that CIB interacts with a site in the αV cytoplasmic domain.
Several reports have shown that CIB can interact with the αIIb cytoplasmic domain by a calcium dependent mechanism and induce the activation of αIIbβ3 integrin; enabling αIIbβ3-mediated cell adhesion or spreading on fibrinogen [6,7]. The membrane proximal GFFKR motif of the αIIb cytoplasmic domain has been shown to be a site of this interaction [3]. However, since this motif is highly conserved among α integrin subunits and since CIB is considered to interact specifically with the αIIb
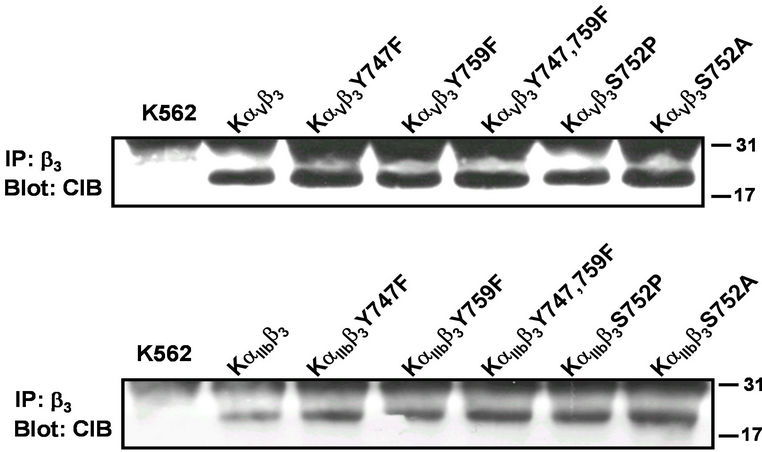
Figure 2. Effect of mutations of β3 cytoplasmic domains on interaction with CIB. CIB blots of αVβ3 and αIIbβ3 immunoprecipitations from KαVβ3Y747F, KαIIbβ3Y747F, KαVβ3Y759F, KαIIbβ3Y759F, KαVβ3Y747/759F, KαIIbβ3Y747/759F, KαVβ3- S752P, KαIIbβ3, KαVβ3S752A, KαVβ3S752P, and KαIIbβ3S752A cells. Cell lysates were immunoprecipitated with mAb 7G2 (anti-β3) and CIB detected within immunoprecipitates by Western blot using mAb UN2-NH (anti-CIB). Shown is a representative of multiple experiments.
 (a)
(a) (b)
(b)
Figure 3. Effect of mutations of αV and αIIb cytoplasmic domains on interaction with CIB. K562 cells expressing cDNA encoding for αIIbβ3 or αVβ3 wild type, αV/αIIbβ3 and αVNTβ3 cells (a) or αV/αIIbβ3 bearing mutations on αIIb subunit cytoplasmic domain (b) were immunoprecipitated with mAb 7G2 (anti-β3) and co-precipitated CIB detected by Western blot using mAb UN2-NH (anti-CIB). Total β3 was determined in a fraction of immunoprecipitates using mAb 7G2 (anti-β3). Shown is a representative of multiple experiments.
cytoplasmic domain, the CIB-αIIbβ3 interaction is probably mediated through a supplementary αIIb cytoplasmic domain. As previously suggested by Hwang and Vogel [4], the αIIb C-terminal LEEDDEEGE domain is suited to play this role. The current model of αIIb cytoplasmic domain structure suggests a “closed” conformation where the highly conserved N-terminal membrane-proximal GFFKR motif forms a α-helix followed by a turn, and the C-terminal loop interacts with the N-terminal helix in an intramolecular interaction [11]. The substitution of αIIb cytoplasmic domain residues Pro998 and Pro999 with alanine residues has been shown to convert this structure to an “open” conformation; inducing the αIIbβ3 activation [13]. To assess the involvement of those different αIIb cytoplasmic domains or their conformation in the αIIb-CIB interaction, we immunoprecipitated β3 from Mn2+ treated K562 cells expressing αIIbβ3 (KαIIbβ3) or chimeric αV/αIIbβ3 integrin receptors bearing the extracellular αV domain fused to the αIIb transmembrane and cytoplasmic domain full length (WT) or lacking the final 9 (999), 13 (995) or 18 (990) last amino acids or with the residues Pro998 and Pro999 substituted by Ala residues (PP/AA). Blotting with anti-CIB antibody (UN2-HN) reveals that CIB can interact only with the full-length αIIb cytoplasmic domain with the native conformation. Thus, the deletion of only last 9 last amino acids (LEEDDEEGE) forming the acid C-terminus (999) or with the conserved motif GFFKR prevents the interaction of the αIIb cytoplasmic domain with CIB suggesting that both domains are involved in this interaction. Interestingly, the destruction of the αIIb cytoplasmic domain conformation by the substitution of residues Pro-998 and Pro-999 by Ala residues (PP/AA) also inhibits this interaction (Figure 3(b)). Thus, more than the presence of LEEDDEEGE and GFFKR domains, the interaction of CIB with the αIIb cytoplasmic domain requires its native conformation. Using the K562 cells expressing chimeric αV/αIIbβ3 integrins with αIIb cytoplasmic domain bearing the deletions or mutations mentioned above, we confirmed these observations by immunoprecipitating the cell lysates with beads coupled to anti-CIB (UN2-NH) and then blotted the transferred proteins with an anti-β3 antibody. In these reverse immunoprecipitations, CIB association with β3 required the entire cytoplasmic tail in native conformation (Figure 4).
Previous evidence suggests that CIB induces the activation of the αIIbβ3 integrin by interacting specifically with the αIIb cytoplasmic tail. In the present work, we show that CIB can be co-immunoprecipitated with β3 integrin associated with αV or αIIb, suggesting a non-exclusive interaction. This observation has been confirmed by the inhibition of the CIB interaction by the deletion of αV cytoplasmic domain. The comparison of the αIIb and αV cytoplasmic tail sequences reveals a few similarities. Both α subunits retain the WKxGFFKR motif that is thought to be CIB binding site [2,4]. In addition, both α subunits contain a PP motif and a very acidic sequence. These two features distinguish αV and αIIb from all other α subunits. Although the WKxGFFKR motif of αIIb cytoplasmic domain is considered as necessary for the CIB binding, it is unlikely to be the unique factor determining this interaction since this motif is common to all α subunits. We show here that the deletion of the distal domain LEEDDEEGE (mutant 999), previously considered as a potential site for interaction with the N-terminus of CIB [4], prevents the co-immunoprecipitation with the dimeric integrin. As expected, the deletion of the membrane proximal domain GFFKR (mutant 995) also prevents this coimmunoprecipitation. Our data suggest a simultaneous participation of these two αIIb cytoplasmic domains in the interaction with CIB. Moreover, we show here that, even in the presence of these domains, this interaction requires the native αIIb cytoplasmic domain conformation suggesting that the spatial disposition of the αIIb binding sites of CIB is also a determinant of this interaction.
It has been shown that αIIb-CIB interaction is calcium dependent [1] and a model suggests that the αIIb cytoplasmic domain contains a high affinity cation binding site [25]. Additionally, Vinogradova et al. have proposed that calcium binding to αIIb cytoplasmic domains is determined by intramolecular interaction between the residues Arg997, Glu1001, Asp1003, and Asp1004 of the native conformation [26]. Since the substitution of residues Pro-998 and Pro-999 by Ala residues (PP/AA) inducing the “open” conformation provokes the destruction of these intramolecular interactions, the inhibition of the CIB-αIIb interaction could be due to inability of the PP/AA mutant to bind calcium cations. The αIIb-β3 interaction is thought to maintain the receptor in a low affinity state through a transmembrane subunit contact domain and GFFKR juxta-transmembrane region and to be calcium dependent [25]. A structural study suggests that
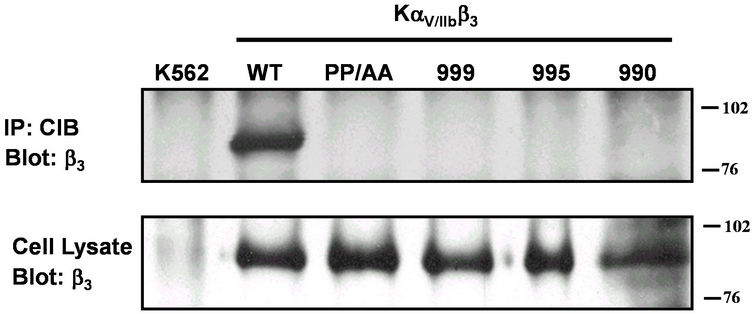
Figure 4. Effect of mutations on αIIb cytoplasmic domains on interaction with CIB. K562 cells expressing cDNA encoding for αV/αIIbβ3 bearing indicated mutations or truncations of the αIIb subunit cytoplasmic domain were immunoprecipitated with mAb UN2-NH (anti-CIB) and β3 detected by Western blotting using mAb 7G2 (anti-β3). Total β3 was determined by Western blot of lysates of cells probed with mAb 7G2 (anti-β3). Shown is a representative of multiple experiments.
the calcium cation is likely to enhance and stabilize the intramolecular interactions of the αIIb cytoplasmic domain but is not essential for the αIIb-β3 interaction. However calcium stabilizes the αβ complex in low-affinity states [27] by decreasing the dissociation rate [2].
We previously showed that β3-tyrosine phosphorylation requires the αV cytoplasmic domain [14] and can be regulated differentially when the β3 subunit is associated with αIIb or αV [15]. This clearly indicates that this critical biochemical process is regulated by the α subunit cytoplasmic domain; probably through steric hindrance determining the access of β3 subunit cytoplasmic sites for its protein binding partners required for post-ligand events. The incomplete homology between αIIb and αV cytoplasmic domain suggests two different conformations generating two different steric hindrances. Therefore, the PP/AA mutation of αIIb cytoplasmic domain gives a new tool to explore the influence of the steric access to β3 cytoplasmic domain interacting proteins. Regardless, the previously reported selective association of CIB with αIIb does not hold true in this hematopoietic cell system and does not offer a mechanism to explain α-subunit regulation of β3 tyrosine phosphorylation. Naik et al., in the yeast two-hybrid system, using the αIIb cytoplasmic domain as “bait,” showed that CIB interacts specifically with the αIIb cytoplasmic domain [1]. Here, we show that this interaction is not αIIb exclusive and suggest that cell environment is a factor determining the α cytoplasmic domain interaction specificity. A recent work suggests that CIB may exist in multiple structural and metal ion-bound states in vivo, which may also play a role in its regulation of target proteins such as platelet integrins [28]. A third partner could also be involved since Rac3 and the polo-like kinase Skn have been shown to interact with CIB [8,9].
4. Acknowledgements
We thank Matthew P. Williams for his excellent technical assistance in cell transfection and sorting. This work was supported by NIH DK79884 awarded to Scott D. Blystone and an American Heart Postdoctoral Fellowship awarded to Innocent H. Yamodo.
REFERENCES
- U. P. Naik, P. M. Patel and L. V. Parise, “Identification of a Novel Calcium-Binding Protein That Interacts with the Integrin AlphaIIb Cytoplasmic Domain,” The Journal of Biological Chemistry, Vol. 272, No. 8, 1997, pp. 4651- 4654.
- L. Vallar, C. Melchior, S. Plancon, H. Drobecq, G. Lippens, V. Regnault and N. Kieffer, “Divalent Cations Differentially Regulate Integrin AlphaIIb Cytoplasmic Tail Binding to beta3 and to Calciumand Integrin-Binding Protein,” The Journal of Biological Chemistry, Vol. 274, No. 24, 1999, pp. 17257-17266.
- W. T. Barry, C. Boudignon-Proudhon, D. D. Shock, A. McFadden, J. M. Weiss, J. Sondek and L. V. Parise, “Molecular Basis of CIB Binding to the Integrin Alpha IIb Cytoplasmic Domain,” The Journal of Biological Chemistry, Vol. 277, No. 32, 2002, pp. 28877-28883.
- P. M. Hwang and H. J. Vogel, “Structures of the Platelet Calciumand Integrin-Binding Protein and the AlphaIIbIntegrin Cytoplasmic Domain Suggest a Mechanism for Calcium-Regulated Recognition; Homology Modelling and NMR Studies,” Journal of Molecular Recognition, Vol. 13, No. 2, 2000, pp. 83-92. doi:10.1002/(SICI)1099-1352(200003/04)13:2<83::AID-JMR491>3.0.CO;2-A
- S. Tsuboi, “Calcium Integrin-Binding Protein Activates Platelet Integrin Alpha IIbbeta 3,” The Journal of Biological Chemistry, Vol. 277, No. 3, 2002, pp. 1919-1923.
- U. P. Naik and M. U. Naik, “Association of CIB with GPIIb/IIIa during Outside-In Signaling Is Required for Platelet Spreading on Fibrinogen,” Blood, Vol. 102, No. 4, pp. 1355-1362.
- M. U. Naik and U. P. Naik, “Calcium-and IntegrinBinding Protein Regulates Focal Adhesion Kinase Activity during Platelet Spreading on Immobilized Fibrinogen,” Blood, Vol. 102, No. 10, 2003, pp. 3629-3636.
- L. Haataja, V. Kaartinen, J. Groffen and N. Heisterkamp, “The Small GTPase Rac3 Interacts with the integrinBinding Protein CIB and Promotes Integrin Alpha(IIb) Beta(3)-Mediated Adhesion and Spreading,” The Journal of Biological Chemistry, Vol. 277, No. 10, 2002, pp. 8321- 8328.
- S. Ma, M. A. Liu, Y. L. Yuan and R. L. Erikson, “The Serum-Inducible Protein Kinase Snk Is a G1 Phase Polo-Like Kinase That Is Inhibited by the Calciumand Integrin-Binding Protein CIB,” Molecular Cancer Research, Vol. 1, No. 5, 2003, pp. 376-384.
- C. Whitehouse, J. Chambers, K. Howe, M. Cobourne, P. Sharpe and E. Solomon, “NBR1 Interacts with Fasciculation and Elongation Protein Zeta-1 (FEZ1) and Calcium and Integrin Binding Protein (CIB) and Shows Developmentally Restricted Expression in the Neural Tube,” European Journal of Biochemistry, Vol. 269, No. 2, 2002, pp. 538-545. doi:10.1046/j.0014-2956.2001.02681.x
- O.Vinogradova, A. Velyvis, A. Velyviene, B. Hu, T. Haas, E. Plow and J. Qin, “A Structural Mechanism of Integrin Alpha(IIb)Beta(3) ‘Inside-Out’ Activation as Regulated by Its Cytoplasmic Face,” Cell, Vol. 110, No. 5, 2002, pp. 587-597.
- A. M. Weljie, P. M. Hwang and H. J. Vogel, “Solution Structures of the Cytoplasmic Tail Complex from Platelet Integrin Alpha IIband Beta 3-Subunits,” Proceedings of the National Academy of Sciences, Vol. 99, No. 9, 2002, pp. 5878-5883.
- T M. Leisner, J. D. Wencel-Drake, W. Wang and S. C. Lam, “Bidirectional Transmembrane Modulation of Integrin AlphaIIbBeta3 Conformations,” The Journal of Biological Chemistry, Vol. 274, No. 18, 1999, pp. 12945- 12949.
- S. D. Blystone, M. P. Williams, S. E. Slater and E. J. Brown, “Requirement of Integrin Beta3 Tyrosine 747 for Beta3 Tyrosine Phosphorylation and Regulation of Alphavbeta3 Avidity,” The Journal of Biological Chemistry, Vol. 272, No. 45, 1999, pp. 28757-28761.
- S. D. Blystone, “Kinetic Regulation of Beta 3 Integrin Tyrosine Phosphorylation,” The Journal of Biological Chemistry, Vol. 277, No. 49, 2002, pp. 46886-46890.
- T. E. O’Toole, J. Ylanne and B. M. Culley, “Regulation of Integrin Affinity States through an NPXY Motif in the Beta Subunit Cytoplasmic Domain,” The Journal of Biological Chemistry, Vol. 270, No. 15, 1995, pp. 8553-8558. doi:10.1074/jbc.270.15.8553
- P. D. Tahiliani, L. Singh, K. L. Auer and S. E. LaFlamme, “The Role of conserved Amino Acid Motifs within the Integrin Beta3 Cytoplasmic Domain in Triggering Focal Adhesion Kinase Phosphorylation,” The Journal of Biological Chemistry, Vol. 272, No. 12, 1997, pp. 7892-7898. doi:10.1074/jbc.272.12.7892
- A. A. De Melker, D. Kramer, I. Kuikman and A. Sonnenberg, “The two Phenylalanines in the GFFKR Motif of the Integrin Alpha6A Subunit Are Essential for Heterodimerization,” Biochemical Journal, Vol. 328, 1997, pp. 529-537.
- R. Pardi, G. Bossi, L. Inverardi, E. Rovida and J. R. Bender, “Conserved Regions in the Cytoplasmic Domains of the Leukocyte Integrin Alpha L Beta 2 Are Involved in Endoplasmic Reticulum Retention, Dimerization, and Cytoskeletal Association,” The Journal of Immunology, Vol. 155, No. 3, 1995, pp. 1252-1263.
- E. Loh, W. Qi, G. Vilaire and J. S. Bennett, “Effect of Cytoplasmic Domain Mutations on the Agonist-Stimulated Ligand Binding Activity of the Platelet Integrin AlphaIIbBeta3,” The Journal of Biological Chemistry, Vol. 271, No. 47, 1996, pp. 30233-30241.
- T. E. O’Toole, Y. Katagiri, R. J. Faull, K. Peter, R. Tamura, V. Quaranta, J. C. Loftus, S. J. Shattil and M. H. Ginsberg, “Integrin Cytoplasmic Domains Mediate Inside-Out Signal Transduction,” The Journal of Cell Biology, Vol. 124, No. 6, 1994, pp. 1047-1059.
- P. E. Hughes, T. E. O’Toole, J. Ylanne, S. J. Shattil and M. H. Ginsberg, “The Conserved Membrane-Proximal Region of an Integrin Cytoplasmic Domain Specifies Ligand Binding Affinity,” The Journal of Biological Chemistry, Vol. 270, No. 21, 1995, pp. 12411-1247.
- T. E. O’Toole, J. Mandelman, J. Foryth, S. J. Shattil and M. H. Ginsberg, “Modulation of the Affinity of Integrin Alpha IIb Beta 3 (GPIIb-IIIa) by the Cytoplasmic Domain of Alpha IIb,” Science, Vol. 254, No. 5033, 1991, pp. 845- 847. doi:10.1126/science.1948065
- D. F. Kucik, T. E. O’Toole, A. Zheleznyak, D. K. Busettini and E. J. Brown, “Activation-Enhanced Alpha(IIb) Beta(3)-Integrin-Cytoskeleton Interactions outside of Focal Contacts Require the Alpha-Subunit,” Molecular Biology of the Cell, Vol. 12, No. 5, 2001, pp. 1509-1518.
- T. A. Haas and E. F. Plow, “The Cytoplasmic Domain of Alphaiib Beta3. A Ternary Complex of the Integrin Alpha and Beta Subunits and a Divalent Cation,” The Journal of Biological Chemistry, Vol. 271, No. 11, 1996, pp. 6017- 6026.
- O. Vinogradova, T. Haas, E. F. Plow and J. Qin, “A Structural Basis for Integrin Activation by the Cytoplasmic Tail of the Alpha IIb-Subunit,” Proceedings of the National Academy of Sciences, Vol. 4, 2000, pp. 1450- 1455. doi:10.1073/pnas.040548197
- C. Spillmann, D. Osorio and R. A. Waugh, “Integrin Activation by Divalent Ions Affects Neutrophil Homotypic Adhesion,” Biomedical Engineering, Vol. 30, No. 8, 2002, pp. 1002-1011.
- A. P. Yamniuk, L. T. Nguyen, T. T. Hoang and H. J. Vogel, “Metal Ion Binding Properties and Conformational States of Calciumand Integrin-Binding Protein,” Biochemistry, Vol. 43, No. 9, 2004, pp. 2558-2568.
NOTES
*Corresponding author.