American Journal of Analytical Chemistry
Vol.4 No.10A(2013), Article ID:37309,11 pages DOI:10.4236/ajac.2013.410A1002
Quantification of Global Protein Disulfides and Thiol-Protein Mixed Disulfides to Study the Protein Dethiolation Mechanisms
1Department of Neuroscience, Pharmacology Unit, University of Siena, Siena, Italy
2Division of Biochemistry, Department of Medical Biochemistry and Biophysics, Karolinska Institute, Stockholm, Sweden
3Brighton and Sussex Medical School, Trafford Centre, Falmer, Brighton, UK
Email: *paolo.cherubini@unisi.it
Copyright © 2013 Lucia Coppo et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received June 28, 2013; revised July 28, 2013; accepted August 15, 2013
Keywords: Protein Disulfides; S-Glutathionylation; Thiol pKa; Diamide; Oxidative Stress
ABSTRACT
The redox state of cellular thiols is widely studied because it was recently linked to many different diseases and pathologies. In this work we quantified the concentrations of protein disulfides (PSSP) and thiol-protein mixed disulfides (XSSP) in rat tissues (liver, kidney and heart) and cells (Raw 264.7) by an improved method of XSSP and PSSP determination after oxidative stress induced by diamide. Under native and denaturing conditions, a thiol block by N-ethymaleimide was introduced to avoid thiol exchange reaction activations by protein SH groups (PSH) (PSH + XSSP ↔ PSSP + XSH) and alterations of original XSSP/PSSP levels. Low molecular weight thiols (XSH) and PSH were respectively measured by HPLC on supernatants and on corresponding pellets by DTNB (Ellman’s reagent) after dithiothreitol reduction. PSSP concentrations of liver, heart and kidney were respectively 0.304, 0.605 and 0.785 µmoles/g and after diamide exposure they were significantly augmented of about 65% - 70% in liver and heart, but not in the kidney. Normal XSSP, that were 6 - 20 times lower than normal PSSP were induced by diamide in liver and heart of about 40 times, but not in kidney. Thermodynamic criteria regarding the pKa values of thiols engaged as PSSP and GSSP were used to interpret dethiolation mechanisms via thiol exchange reactions.
1. Introduction
The redox state of glutathione (GSH) and protein SH groups (PSH) is the key point in the redox regulation of cells. Thiols are engaged in molecular competitions [1] to react with electrophiles and oxidants generated due to oxidative or nitrosative stress [2-6] or intracellularly as signaling molecule, like hydrogen peroxide [7]. Thiol oxidations, in particular S-glutathionylation (GSSP) or protein disulfides (PSSP) formation, switch on/off essential PSH, and the PSH regeneration is promoted by enzymes, NADPH consumption, and thiol exchange reactions:
P-SH + XS-SX’ ↔ XS-SP + X’-SH (1)
P’-SH + PS-SX ↔ PS-SP’ + X-SH (2)
X-SH + XS-SP ↔ P-SH + XS-SX (3).
Thiol exchange reactions are involved in a great variety of physiological processes of which the cellular redox regulation [2,3,8-11], the blood coagulation [12], the control of plasma thiols and disulfides [6], the platelet aggregation [13], the activity of SNARE proteins [14] are only few examples that emphasize their biological importance. Although the macromolecular events regarding the thiol exchange processes, generally described by (1 - 3) reactions, are mostly enzymatic, it is however possible that under appropriate thermodynamic and steric conditions, the non-enzymatic reactions cooperate with the enzymatic ones to perform physiological actions. At the same time, it is also possible that the activation of nonenzymatic reactions could prelude the onset of pathological actions as suggested by various examples. In fact, since SH enzymes, such as peroxiredoxins and sulfiredoxins, are involved in models of Alzheimer or Parkinson diseases with neuroprotective roles [15,16], it is possible that the disruption of these mechanisms is involved in the onset of pathological events. For example, it is known that increased S-glutathionylated proteins [17] and increased formation of specific PSSP (e.g., protein disulfides of glyceraldehyde 3-phosphate dehydrogenase) [18] have been found in the Alzheimer disease and in mitochondrial enzymes in an animal model of Parkinson disease [19].
After the cellular XSSP and PSSP production, the return to PSH is characterized by rather complex molecular events of deglutathionylation. The dethiolation of albumin mixed disulfides [20,21] is paradigmatic for a better comprehension of the mechanisms of thiol exchange processes occurring at cellular level [20,21]. Albumin has only one free Cys residue (Cys34) with a low pKa value (of about 5 - 7), that in dependence of pKa difference of thiols of XSSP may be equally well transformed nonenzymatically into PSH, by reaction (3) (if pKa of PSH < pKa of XSH) or into a new thiol-protein mixed disulfide (X’SSP), equivalent to reaction (2), (if pKa of PSH > pKa of XSH) [20, 21]:
X’-SH + PS-SX ↔ X’S-SP + X-SH (4).
The same possibilities of XSSP and PSSP metabolism can potentially exist at cellular level in dependence of the difference of pKa values of thiols engaged in XSSP or PSSP.
Studies regarding the GSSP/PSSP relationships may serve to deeper understand the physiological and pathological mechanisms of protein dethiolation. These mechanisms involve various protein factors [22,23], and their biological relevance is much augmented after the discovery in cells of a great variety S-glutathionylated proteins [24-26], and of PSSP, engaged not only in structural functions but also in nuclear [3], allosteric [9,11, 27], or catalytic [28,29] actions, and of heterogeneous PSSP produced by functionally different SH proteins [8, 10].
To facilitate the comprehension of the GSSP/PSSP metabolism by thiol exchange reactions and the mechanisms of protein dethiolation, we here propose a method of XSSP and PSSP determination on rat tissues and cells (Raw 264.7).
This method based on a good control of oxidative artifacts and checked by specific experiments of oxidative stress, resulted to be easy, rapid, and reliable. The assay is characterized by a double robust pretreatment of samples with N-ethylmaleimide (NEM), and by the disulfide reduction by dithiothreitol (DTT).
The protein-protein disulfides (PSSP) measure is obtained by the following calculation:
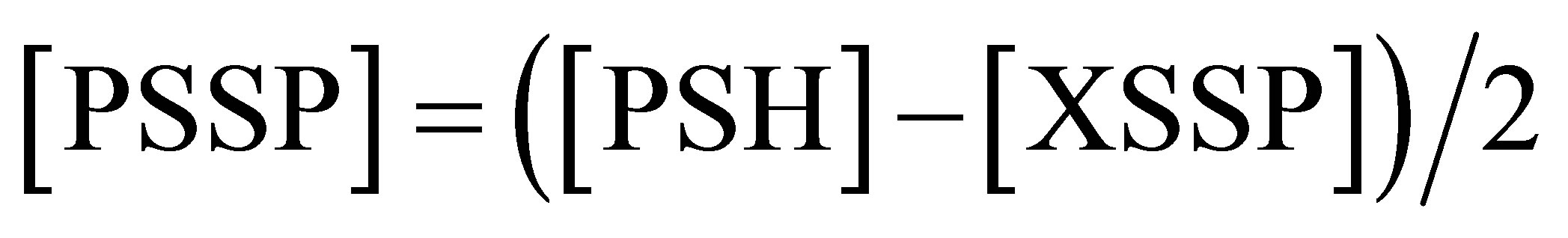 (5)
(5)
where PSH, assayed on the protein pellet, derives from reduced XSSP and PSSP, whereas XSSP is assayed on the supernatants, as XSH released from reduced XSSP, measured by HPLC.
The importance of thermodynamic factors of the SH group that condition the XSSP/PSSP metabolism by thiol exchange reactions is discussed.
2. Materials and Methods
2.1. Chemicals
Methanol and HPLC grade reagents were obtained from Merck (Darmstadt, Germany). Monobromobimane (mBrB) was obtained from Calbiochem (La Jolla, CA, USA). Diamide, N-ethylmaleimide, 5,5’-dithiobis (2-nitrobenzoic acid) (DTNB), and all other reagents were from Sigma-Aldrich Corp. (St. Louis, MO, USA).
2.2. Animals
Adult male Sprague-Dawley rats (Charles River, Italia), weighting ~350 g, were housed under controlled light (12 h on and 12 h off), temperature (20˚C ± 2˚C) and humidity (55% ± 5%). The animals consumed food and water at libitum. Experiments were carried out in accordance with the Guidelines of the Council of European Community 86/609/EEC and the approval of the Bioethical Committee of the University of Siena.
The rats were sacrificed after intraperitoneal anaesthesia (50 mg/kg sodium pentobarbital and 0.4 mg/kg scopolamine) and tissues were removed, rapidly washed in saline, blotted and used for experiments or placed in liquid nitrogen and stored at −80˚C.
2.3. Tissue Homogenates
From each tissue four independent shares (0.2 g of tissue) were processed and subdivided in the following 4 groups: CTR (control), DIA (diamide group), NEM (N-ethylmaleimide group), and (DIA + NEM) (diamide plus NEM group) (Figure 1).
Tissues samples of the four groups were homogenized by an Ultra-Turrax homogenizer (20 s) in ice-cooled plastic tubes, at weight-volume ratio of 1:5, in PBE (0.2 M phosphate buffer, pH 7.4, with 2.5 mM K3EDTA), except the NEM group that was prepared in 310 mM NEM in PBE.
Another portion of the same tissue (0.2 g at same weight/volume ratio), named basal (BAS) was directly homogenized in 6% (w:v) TCA.
2.4. Second Alkylation
The protein pellets were washed three times with 1.5% TCA. Next, the NEM and DIA + NEM pellets were resuspended in 1.0 ml of 310 mM NEM, whereas the CTR and DIA groups were resuspended in the same PBE volume.
After 10 min under stirring at RT, samples were TCAprecipitated (6% TCA, final concentration), centrifuged

Figure 1. Experimental design of PSSP determination. Flow chart defines single steps. Diamide mainly oxidizes thiols to disulfides, restrains overoxidation phenomena to disulfides and is a good tool to study thiol exchange reactions. The sequential NEM treatments serve to block all free thiols. The last pellet resuspended at pH 8.2 and treated with DTT serves to reduce XSSP and PSSP in all groups. The last pellet resuspended at pH 8.2 without DTT treatment (+H2O) serves to reduce XSSP by reaction (2) in CTR, DIA and BAS (not reported in Figure) groups. The last pellet of NEM and DIA + NEM groups, resuspended at pH 8.2 without DTT treatment (+H2O), serves to reduce the PSSP subpopulation, namely the pH-dependent PSSP (see text).
(7000 × g, 5 min, 4˚C) and washed, as above reported.
2.5. pH Adjustment and DTT Treatment
The protein pellets of the four groups and those of BAS group were resuspended at pH 8.2 with 1.2 ml of a basic solution, prepared with additions of 9 vol of 0.1 M phosphate buffer, pH 7.4, with 1 vol of 1 N NaOH.
Each sample was split in two aliquots and treated with 7.7 mM DTT (final concentration) or distilled water. The comparison between DTT and no-DTT samples was performed because it was unexpectedly found that, despite the double NEM treatment, the samples not reduced by DTT at pH 8.2 revealed a positive signal with DTNB.
After 20 min at RT under stirring, all the samples, with and without DTT were precipitated with 6% TCA and centrifuged as above. Supernatants SNs were used to assay XSH by HPLC and the pellets were washed and centrifuged as above and then used to assay PSH by DTNB.
2.6. PSH Assay
The pellets of DTT and no-DTT samples were resuspended in 0.2 ml of 10 mM 5,5’-dithiobis(2-nitrobenzoic acid) (DTNB) in 0.2 M phosphate buffer, pH 7.4. After 20 min at RT under stirring in the dark, samples were centrifuged (7000 × g, at 4˚C, 5 min) and after appropriate dilution of the clear SNs were assayed at 410 nm in a V-550 JASCO spectrophotometer by the Ellman’s method (DTNB, εmM 410 =13.64 mM−1∙cm−1) [30].
The PSH data and those of XSSP (see below) of DTTtreated samples of NEM and DIA + NEM groups were used to calculate the PSSP concentrations of tissues according to Equation (5).
2.7. HPLC Analysis
XSH released from disulfides (XSSP) of DTT-treated samples subjected or non-subjected to NEM treatment (see data of Table 1) were measured by HPLC, by the method previously reported [6,31]. 0.1 ml of SNs were diluted with 0.04 ml H2O, and 0.03 ml of this solution plus 0.07 ml H2O were saturated with solid NaHCO3 (pH ~ 8.0) and incubated with 1mM mBrB (final concentration) in the dark at RT for 15 min. After centrifugation, SNs were acidified with 3 µl of 37% HCl to pH 3 - 4 and transferred into vials for the HPLC determination.
The HPLC apparatus (Hewlett-Packard 1100 Series) was equipped with fluorescence detection and an OmniSpher C-18 reversed-phase column, Varian. The chromatographic conditions were: mobile phases 0.25% (v:v) acetic acid, adjusted to pH 3.09 with 1 mM NaOH (A) and methanol (B). Elution profile was 0.8 min, 20% B; 8 - 15 min, 20% - 40% B; 15 - 25 min, 40% - 100% B (1.0 ml/min flow rate; fluorimetric detector: excitation, 380 nm; emission, 480 nm). CSH and GSH retention times were 6.0 min and 12.2 min, respectively.
2.8. XSSP Determination of Protein Pellets Activated by Reaction (2)
XSSP determination was also performed by the previous described HPLC procedure on XSH released from the protein pellets by the spontaneous activation of reaction (2) at neutral-basic pH of not-NEM treated and not-DTT treated samples (see in Table 2 data of BAS, CTR, and DIA groups).
2.9. Cell Line and Cell Culture
Raw 264.7 mouse macrophage cells were grown in T75
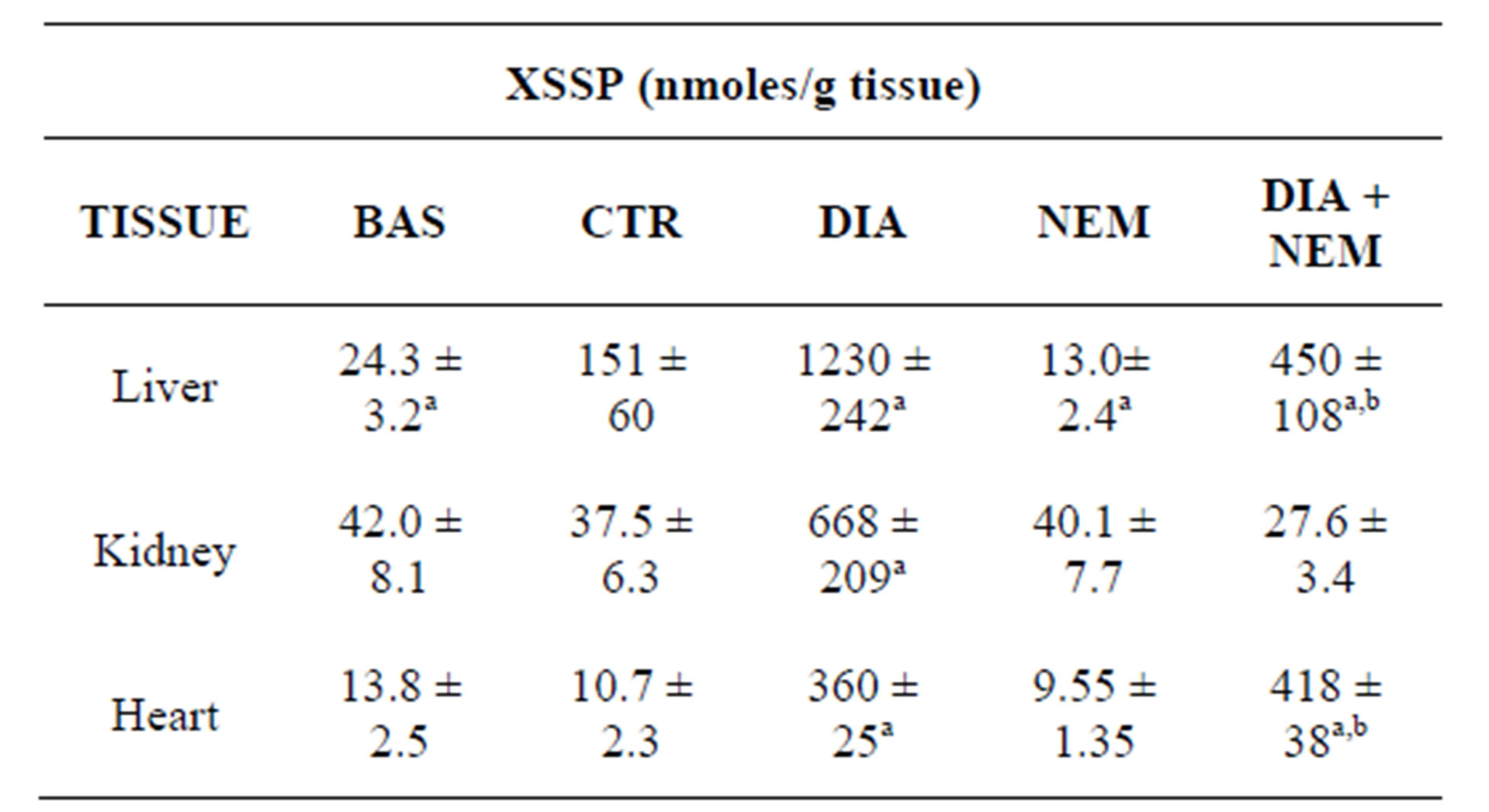
Table 1. XSSP concentrations of tissue homogenates subjected to oxidative stress, by air and/or diamide.
aSignificant as compared to CTR (P < 0.05); bSignificant as compared to NEM (P < 0.05).

Table 2. The concentrations of XSSP and PSH of tissue homogenates measured in samples not subjected to DTT reduction. Data of XSSP of non-NEM treated groups reveal the spontaneous activation of reaction (2) at pH = 8.2.
aSignificant as compared to CTR (P < 0.05); b% represents the percent ratio between the PSH concentrations measured in the same samples non-reduced and reduced by DTT (see PSH in Table 4). ND = not determined.
flasks under standard incubation conditions (37˚C, 5% CO2) in RPMI-1640 medium containing 100 U/ml penicillin, 100 μg/ml streptomycin, 2 mM L-glutamine, 10% heat-inactivated fetal bovine serum. Cells were used at 90% confluence (2 × 106 cell/ml).
2.10. Sample Preparation from Cells
Each flask was washed twice with DPBS 1X (Dulbecco’s Phosphate Buffered Saline) and the cells were harvested with a scraper in 2 ml of DPBS 1X. The total cells were divided in four equal shares (1 ml each, containing about 4 × 106 cell/ml) that were named CTR, DIA, DIA + NEM and NEM groups. Stock solutions of NEM and diamide were prepared in DPSB 1X.
The DIA and DIA + NEM groups were treated with 1 mM diamide (final concentration), the NEM group was treated with 50 mM NEM (final concentration) and the same volume of DPBS 1X was added to the CTR to respect the total volume of all samples. After 10 min of incubation at RT, the group DIA + NEM was treated with 50 mM NEM, whereas the same volume of DPBS 1X was added to the other groups.
After 10 min of incubation at RT, all groups were acidified with 6% TCA (final concentration), centrifuged at 10,000 × g for 10 min and the supernatants were stored at −80˚C for successive analysis.
The protein pellet samples were washed twice with 1 ml 1.5% TCA, and the NEM and DIA + NEM groups were resuspended and treated with 1 ml 50 mM NEM (final concentration) and incubated at RT for 15 min. After TCA precipitation and washing as above, all pellet samples (CTR, DIA, NEM, and DIA + NEM) were resuspended in 0.5 ml NaOH 0.05 N (final pH ~ 8.2) and divided into two equal shares.
One part was treated with 7.7 mM DTT (final concentration) and the other with the same volume of water. Samples were kept for 20 min under stirring at RT and then treated with 6% TCA, centrifuged 5 min at 10,000 × g and the pellets used for the PSH assay.
2.11. Spectrophotometric Analysis of Cell PSH
Pellets were washed three times with 1.5% TCA, resuspended in 0.14 ml of 0.2 M phosphate buffer pH 7.4 and then 0.01 ml sample were used for protein assay and the remaining sample was used for DTNB analysis. Samples were incubated with 5 mM DTNB (final concentration) for 10 min under stirring at RT and centrifuged at 13000 × g, 5 min. SNs were analyzed at 410 nm by spectrophotometer (Microplate Reader Synergy HT , Biotek).
2.12. Protein Assay
Protein concentration was determined by DC Protein Assay kit from Biorad.
2.13. Statistical Analysis
The results are presented as mean ± standard deviation (SD) of 5 independent experiments. Statistical analysis was performed using a Microsoft Excel® macro (Microsoft Corporation). Data were analyzed by Student’s t test. Results were considered statistically significant when P < 0.05.
The comparison of PSH and XSSP data between CTR and BAS, CTR and NEM, and DIA and DIA + NEM groups served to evaluate the entity of oxidation, produced by air oxygen or DIA.
3. Results
3.1. The Oxidative Artifact Problem
The onset of oxidative artifacts is a recurrent risk present in the thiol assay in biological samples of cells and tissues. In our specific case, this event can activate the reaction (2) and alter the original XSSP/PSSP concentrations of cells and tissues. These pitfalls are generally avoided by the procedure of SH alkylation by NEM, that we modified to have the best guarantee of blockage of those PSH, that under native conditions were masked or partially masked but that can eventually participate to thiol exchange reactions after denaturation, protein precipitation and adjustment to neutral-basic pH values. Therefore we adopted a double procedure of NEM utilization, in native and denatured samples (Figure 1).
To analyze the possible formation of oxidative artifacts during the sample manipulation we compared the results of protein thiols (PSH) and protein mixed disulfides (XSSP) concentrations obtained in native (CTR) and denatured samples (the BAS group), whose BAS homogenates had been prepared directly in TCA. The same comparison was carried out in homogenates treated with diamide (DIA + NEM group) or without NEM exposure (DIA group). Finally, the comparison between DIA + NEM and NEM groups served to explore the effect of residual thiols (GSH and PSH), remained during the DIA treatment, on the production of artifacts. All these experiments were performed on fresh or stored at –80˚C tissues, without observing differences of XSSP/ PSSP results.
The protein-protein disulfides (PSSP) concentrations were measured using the formula:
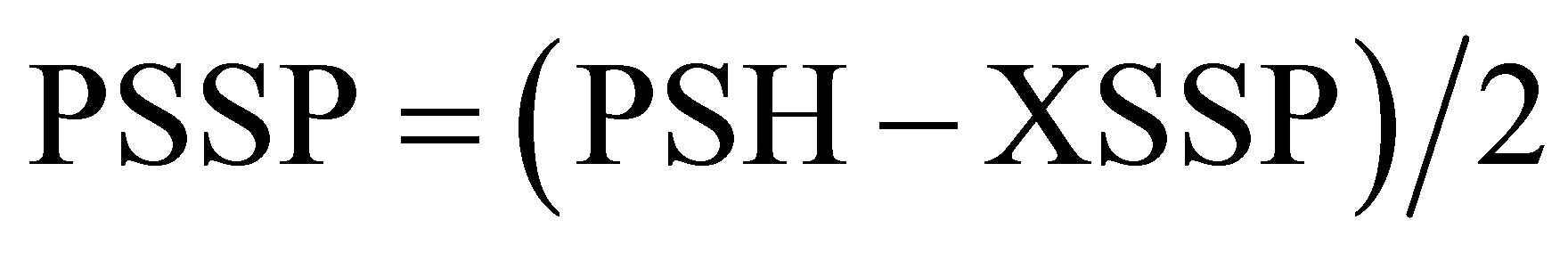 where PSH, assayed with DTNB, were the total PSH, deriving from XSSP/PSSP reduction by DTT, and XSSP, assayed with conventional methods by HPLC, were the sum of XSH on the corresponding supernatant.
where PSH, assayed with DTNB, were the total PSH, deriving from XSSP/PSSP reduction by DTT, and XSSP, assayed with conventional methods by HPLC, were the sum of XSH on the corresponding supernatant.
The PSH oxidation of native samples was measured as PSH decrease (Table 3) as well as XSSP increase (Table 1) after treatment with diamide.
Diamide generates disulfides (XSSX, XSSP, and PSSP) [32], which are reduced by DTT. Conversely DTT is ineffective for the reduction of derivatives of sulfinic (PSO2H) or sulfonic (PSO3H) acids.
After oxidative stress, by diamide or air exposure, the PSH concentrations of tissues, except for the heart, remained unchanged (Table 3). These results suggest that the sample oxidations, by diamide or air (compare the DIA group with CTR group and CTR with BAS), can only generate disulfides and not higher oxidation products (sulfinic or sulfonic acids).
Moreover, the comparison of PSH concentrations of non-DTT treated and DTT-treated samples (see the percentages of non-DTT on DTT in Table 3) evidenced modest or absent PSH over-oxidations of tissue homogenates, except for the heart. This would indicate a scarce exposure or susceptibility of the majority of the PSH population to be oxidized to disulfides.
In experiments of oxidative stress, the XSSP levels (Table 1) increased as expected. They were the sum of glutathionylated and cysteinylated proteins (GSSP and CSSP) in that other S-thiolated species were not found in tissues (and in cells, see after) at our experimental conditions. In particular, the protein mixed disulfides (XSSP) comparison between BAS and CTR groups showed that the air oxygen is able to increase the XSSP levels only in the liver homogenates. The diamide exposure for 20 min
(compare the DIA and CTR groups) and for 10 min (compare the NEM and DIA + NEM groups) augmented a lot the XSSP levels in all tissue homogenates except in the case of kidney of the DIA + NEM group.
3.2. The PSSP Concentration in Tissue Homogenates Subjected to Oxidative Stress
The correct protein-protein disulfides concentration of tissues is obtained in samples subjected to the NEM pretreatment. The basal PSSP concentration of different rat tissues is that expressed by the NEM group in Table 4. These values were increased after diamide exposure (see the DIA + NEM group, Table 4).
In control tissues, the PSSP levels of liver, kidney and heart ranged between about 0.3 - 0.8 μmoles/g, of liver and kidney, respectively. The PSSP concentration has been measured in rat stomach and liver by other authors [33]. The hepatic values reported in Table 4 were similar to those shown in the literature [33].
The protein-protein disulfides levels of normal tissues were higher than those of protein mixed disulfides (see the XSSP of NEM group in Table 1). In particular the hepatic ones resulted about 25 times higher than XSSP, whereas the cardiac ones were nearly 60 fold higher than the corresponding XSSP.
After diamide treatment of tissue homogenates (see data of DIA + NEM group in Table 4) the PSSP concentrations increased significantly in liver and heart, being
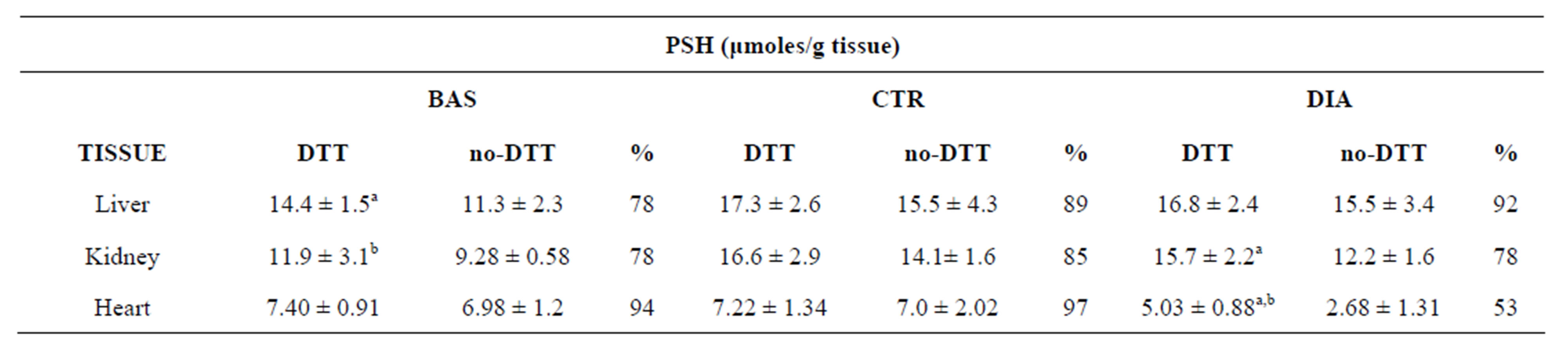
Table 3. PSH concentrations of tissue homogenates subjected to oxidative stress by diamide. Data reveal a modest oxidation entity of the PSH population to disulfides.
BAS and CTR groups are control groups respectively prepared under denaturing and native conditions (see Methods). PSH were assayed with the Ellman’s method [30]. The percent of no-DTT/ DTT ratio (%) reflects the attack of oxidants (diamide plus air oxygen) to PSH. Means ± SD. aSignificant as compared to no-DTT in the same group (P < 0.05). bSignificant as compared to DTT-treated samples of CTR (P < 0.05).
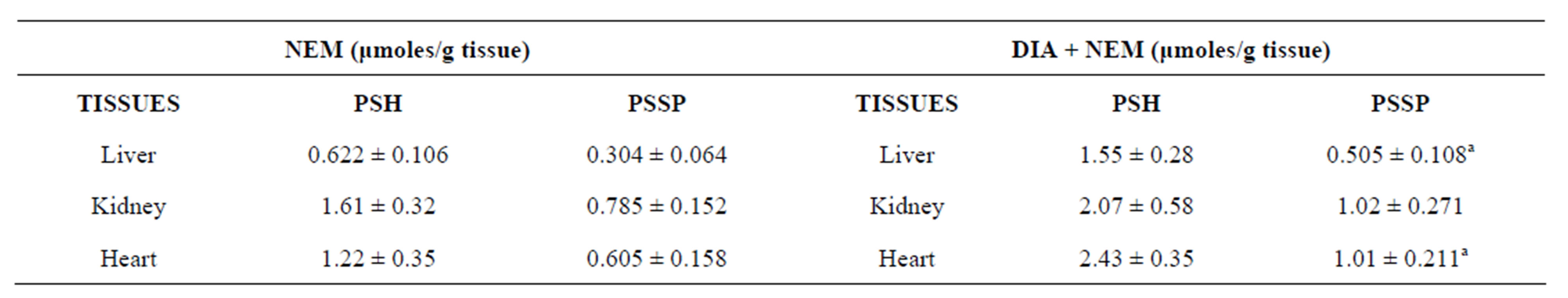
Table 4. PSSP concentration in tissue homogenates.
PSH assayed by DTNB derive from the reduction of PSSX and PSSP with DTT. PSSP is calculated by the formula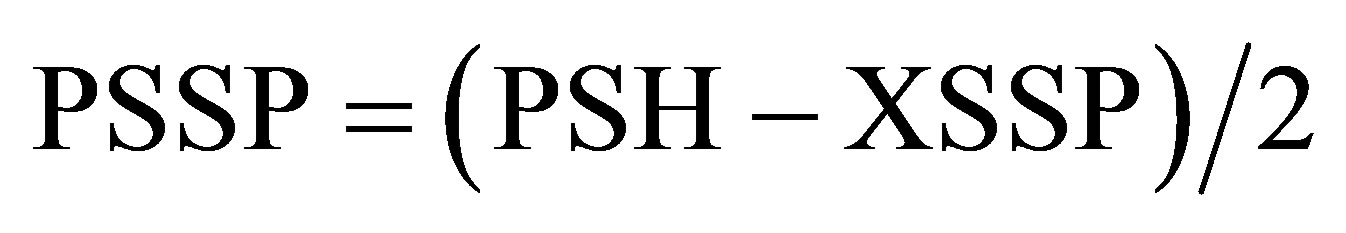 . The PSSP concentration increased after oxidative stress by diamide in all the tissues (not significant in the kidney).Means ± SD. aSignificant as compared to NEM (P < 0.05).
. The PSSP concentration increased after oxidative stress by diamide in all the tissues (not significant in the kidney).Means ± SD. aSignificant as compared to NEM (P < 0.05).
their levels about 65% higher than the corresponding controls, whereas those of kidney were non-significantly augmented.
Other authors with different methods and models [8,10, 33-35] have measured changes in PSSP/GSSP concentrations, and a parallel GSSP and PSSP growth had been found after treatment with diamide in agreement with our data [8,35].
3.3. The Spontaneous Activation of Reaction (2)
We have previously demonstrated [31,36] that the protein pellets of cell and tissue homogenates, resuspended at neutral-basic pH, can spontaneously activate the reaction (2) and release GSH. The measure of GSH concentration over time has been exploited to assay the reversible process of XSSP (GSSP) formation of biological samples subjected to oxidative stress. The XSSP measurements in controls (CTR and BAS groups) and diamide treated homogenates (DIA group), not subjected to DTT reduction of Table 2 confirm this notion.
3.4. The Putative Presence of pH-Dependent PSSP
During the setup of the experiments carried out to verify the NEM efficiency to block thiols, it was found that our samples were positive to the DTNB assay. In particular, whereas the XSSP concentrations of NEM group and DIA + NEM group, not subjected to DTT reduction, displayed zero XSSP values (Table 2), as expected, the DTNB assay on the corresponding pellet resuspended at pH 8.2 for 20 min showed surprisingly the presence of thiols (Table 2). The phenomenon was of remarkable entity and present in all groups except in NEM group of the liver. We assume that PSH deriving from a sub-PSSP population susceptible to pH-dependent hydrolysis were present in our samples. This population could be originally present in each tissue even if we cannot exclude the presence of artifacts produced during the PSSP assay.
3.5. The PSSP Determination in RAW 264.7 Cells
We next assayed the method of PSSP determination in cultured cells (RAW 264.7). At the same time we experimented whether in samples of NEM-treated cells not subjected to DTT reduction, it was possible to hydrolyze PSSP at pH 8.2, to confirm the presence of pH-dependent PSSP. The results of Figure 2, indicate that in NEMtreated cells of NEM and DIA + NEM groups, reduced by DTT, it is possible to measure the cellular PSSP concentration applying Equation (5) (data not shown), and that in non-DTT treated samples of these groups, after the sample incubation at pH = 8.2, PSH can be dosed in these samples.
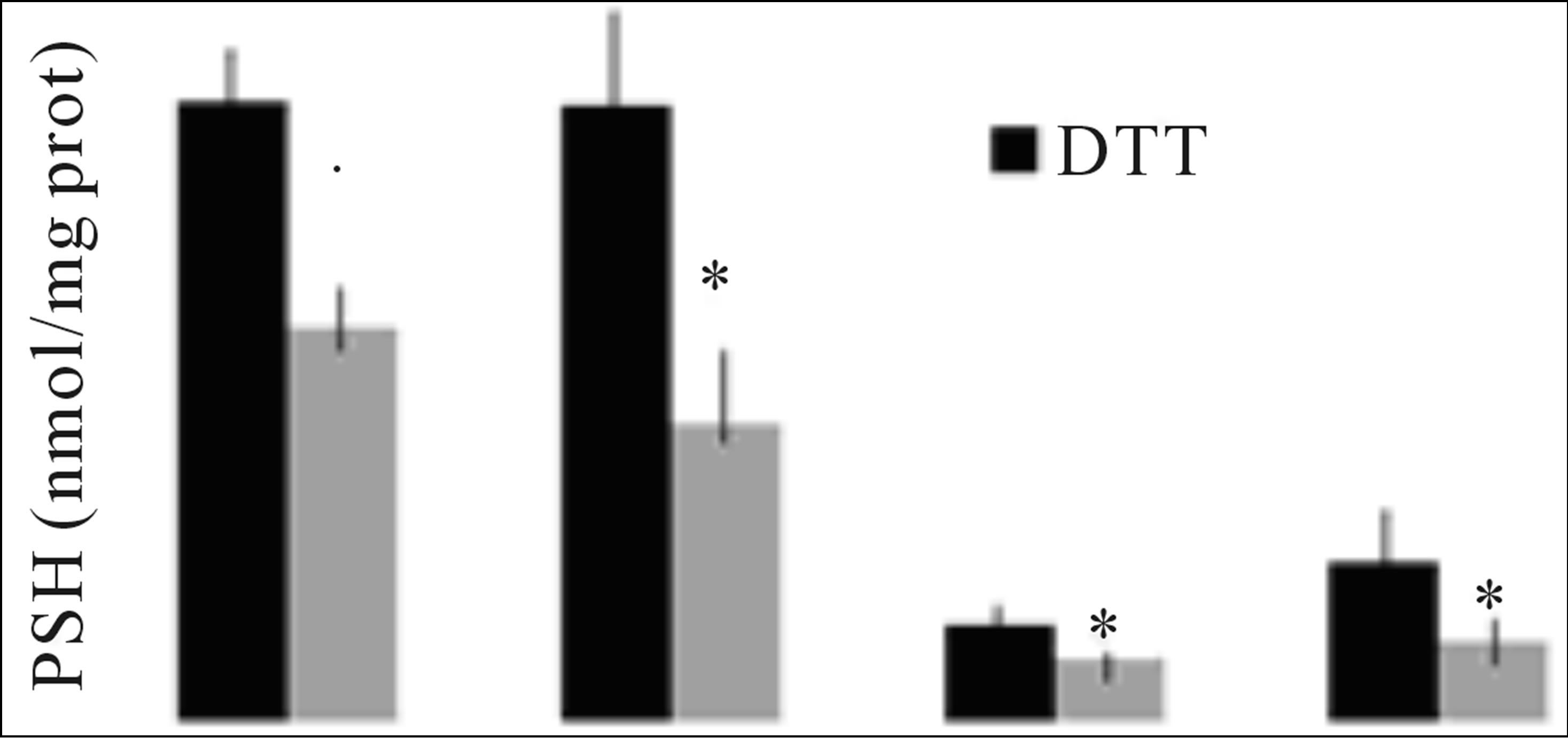
Figure 2. PSH concentration in RAW 264.7 cells. Samples of each group were treated with and without DTT (see materials and methods). Means ± SD *P < 0.05: with respect to DTT-treated groups.
4. Discussion
The thiol and disulfide determinations in cells and tissues (for example the GSH/GSSG ratio) have always presented the problem of oxidative artifacts that has usually been solved by the block of thiols by alkylating agents. In our case, the problem was to avoid the activation of reaction (2) and the alteration of original disulfide concentrations (XSSP/PSSP), due to artifact production and conformational changes of denatured proteins. The double NEM exposure, first under native and then under denatured conditions, was effective to solve the problem and get reliable XSSP/PSSP results equivalent to those of the literature [31,33]. Tissue PSSP levels were measured and their concentrations varied within a range of about 0.3 µmoles/g of liver and of 0.8 µmoles/g of kidney, with values much higher than the corresponding XSSP. XSSP were the sum of GSSP and CSSP, and they displayed a XSSP minimum of 10 nmoles/g of heart and a maximum of 40 nmoles/g of kidney. Other XSSP, at homocysteine and cysteinylglycine, were tested but their concentrations resulted negligible or absent (data not shown).
After homogenate treatment with 1 mM diamide, the PSSP levels were induced in all tissues (Table 4), as expected, and a similar inductive behavior was observed for XSSP (Table 1). The entity of XSSP increases was proportionally higher than that of PSSP, but not as absolute value in that induced XSSP remained lower than induced PSSP. This could indicate that after diamide exposure, the PSSP formation, in addition to thiol exchange reactions, could be determined by radical reactions (PS. + PS. → PSSP).
In non-NEM treated homogenates, the diamide exposure (DIA group) provoked a XSSP induction with respect to control (CTR) much higher than that observed in NEM treated samples (compare XSSP of NEM and DIA + NEM groups) (Table 1). Even though these differences could be attributed to the longer period of diamide exposure (20 min for DIA and 10 min for DIA + NEM), we assume that, in addition to the time effect, the induction difference could be attributed to activations of thiol exchange reactions of residual thiols (namely GSH and PSH) of DIA group.
In rats treated with diamide, the GSSP and CSSP concentrations in erythrocytes [6] increased reversibly over time and the phenomenon was attributed to complex processes of thiol exchange reactions in plasma and erythrocytes responsible of the return to normal GSSP and CSSP levels in both compartments. Future similar experiments could be useful to establish the possible PSSP formation and a better comprehension of the GSH and CSH role during dethiolation phenomena of XSSP and PSSP.
In pellet samples treated with NEM but not reduced by DTT (see PSH data of Table 2), we got the indication that the PSSP population of tissues and cells could contain a sub-population, that is easily hydrolysable at pH 8.2. This population (measured as PSH percentage over the total PSH in Table 2) seemed to be present at relatively high concentrations in normal tissues and do not be induced by diamide. Surprisingly it was absent in the normal, but present in the induced liver. At the moment it was called pH-dependent PSSP, but we do not know whether they are originally present in cells and tissues or are artificially generated during the PSSP assay. However, examples of PSSP or XSSX with labile disulfide bonds, namely easily hydrolysable at moderate basic pH, already exist. For example, pH-sensitive PSSP have been observed in plasma [27], and the dimer of albumin PSSP is broken at moderately alkaline pH [37]. Another example of labile disulfide bond is represented by DTNB that at alkaline pH, below 9.5, starts to be split into its thiolated anions. Besides, in preliminary studies regarding the thiol content and the anti-aggregating properties of garlic ethanolic extracts on human platelets, we have recently found (data not shown) that extracts, that did not contain free thiols, are able to give a rapid intense DTNB response when they are brought at pH = 8.0.
After PSSP formation, by radical reactions or by reaction (2), the PSH regeneration is enzymatically regulated by thioredoxin with NADPH consumption. However, under particular conditions, regarding the PSSP conformation (the PSSP accessibility), and the pKa values of PSH forming PSSP, the thiol exchange reactions of PSSP, like the reduction by thioredoxin, might contribute to reform original PSH.
The dethiolation processes of XSSP of albumin [21,22] and the notions regarding the mechanisms of thiol exchange reactions [38] are useful for a better prediction of the processes of PSSP dethiolation.
In dependence of the pKa difference of XSH and PSH of XSSP, the XSH releasing from XSSP of albumin may occur via reaction (3) or (4). In particular, the reaction (4) of albumin is equivalent to reaction (2), since these reactions differ only for thiol (PSH or XSH) that intervenes in the nucleophilic attack. On the contrary the reactions (2) and (3) express different thermodynamic tendencies that are identifiable on the basis of the leaving groups. These groups are those thiols that in thiol exchange reactions of XSSP and have the lowest pKa value [21,22], namely XSH in reactions (2) or (4), or PSH in reaction (3), respectively.
Similar considerations can be used to predict the PSSP dethiolation via thiol exchange reactions. For example, the representation of the reaction: PSSP’ + GSH ↔ GSSP’ + PSH expresses the concept that PSH has a lower pKa value than P’SH. In turn, if GSSP’ P’SH had a pKa value higher than GSH, the eventual successive GSSP’ dethiolations by thiols (P”SH, XSH or GSH) would be more prone to generate protein disulfides (P”SSP’), via reaction (2), (GSSP’ + P”SH ↔ P’SSP” + GSH), or thiol-protein mixed disulfides (XSSP’), via reaction (4), than GSSG by reaction (3) in that the reaction (3) would be hampered for thermodynamic reasons.
Dethiolations via reaction (3) are typical of GSSP of rat hemoglobin, whose Cys125 of the beta chain has a pKa lower than 7, and reactivity much higher than GSH [6,39-41], whose pKa value is about 9.0. The mechanisms activated by reactions (3) must be equally common as those of reaction (2), because a great variety of S-glutathionylated proteins exist [24-26] whose SH groups have relatively low pKa values [5,26,42]. The dethiolations of these proteins would be facilitated by enzymes (glutaredoxin) and by the GSSG reduction by glutathione reductase with NADPH consumption.
From these considerations on PSH of rat hemoglobin, it is easy to explain the paradox by which, after protein denaturation and pH adjustment, pellets of rat erythrocytes in the absence of free GSH are rapidly dethiolated by reaction (2) although the reaction should be thermodynamically impeded because GSH has a pKa value higher than Cys125 of the beta chain of rat hemoglobin. In the absence of free GSH, hemoglobin is initially dethiolated by reaction (6) that is equivalent to reaction (3), where PSH is the leaving group:
GS-SP + P’-SH ↔ GS-SP’ + P-SH (6).
Successively, if P’SH of reaction (6) had pKa values higher than GSH, the possible interactions of GSSP’ with P”SH would give GSH as leaving group, as we have demonstrated [31,36]. This would occur by activation of reaction (7), which is equivalent to reaction (2):
GS-SP’ + P”-SH ↔ G-SH + P’S-SP” (7).
Under native conditions, the proteins subjected to dethiolation with GSH release described by reaction (2) are those having two vicinal Cys, whose distance is included between 2 - 6 amino acid residues. When one of these Cys is engaged as GSSP, the protein is prone to give intramolecular PSSP [43,44]. All PSSP, formed or not formed by vicinal thiols, are induced by diamide [8,19, 35,43,44] and in particular cases the return to original PSH can be performed by thiol exchange reactions, schematically represented by the coupling of the reverse of reaction (2) with the reaction (3). This would be possible if both the PSH of PSSP had a pKa value lower than GSH. Conversely, if one PSH had a pKa value higher than GSH, the coupling of reactions (2) and (3) could not be realized, in that disfavored for thermodynamics reasons, and the PSSP reduction should require the enzymatic intervention of thioredoxin with NADPH support. We assume that the investigation of the GSSP/ PSSP relationship during time-course experiments of oxidative stress, by the GSSP and PSSP analysis here proposed, could establish cases of PSH regeneration from PSSP via thiol exchange reactions.
In conclusion, we have here proposed an improved method to assay XSSP and PSSP in biological samples that may be useful to deeper understand the mechanisms of protein dethiolation regenerating PSH after oxidative stress events. In our opinion, the knowledge of pKa values of proteins involved in PSSP and XSSP may help to clarify the complex scenario of protein dethiolation at cell level and the reason why the interest on GSH must go beyond its antioxidant function [45].
REFERENCES
- J. W. Cuozzo and C. H. Kaiser, “Competition between Glutathione and Protein Thiols for Disulfide-Bond Formatin,” Nature Cell Biology, Vol. 1, No. 3, 1999, pp. 130- 135. http://dx.doi.org/10.1038/11047
- S. O. Kim, K. Merchant, R. Nudelman, W. F. Beyer Jr., T. Keng, J. DeAngelo, A. Hausladen and J. S. Stamler, “OxyR: A Molecular Code for Redox-Related Signaling,” Cell, Vol. 109, No. 3, 2002, pp. 383-396. http://dx.doi.org/10.1016/S0092-8674(02)00723-7
- A. Delaunay, D. Pflieger, M.-B. Barrault, J. Vinh and M. B. Toledano, “A Thiol Redox Peroxidase Is an H2O2 Receptor and Redox-Transducer in Gene Activation,” Cell, Vol. 111, No. 4, 2002, pp. 471-481. http://dx.doi.org/10.1016/S0092-8674(02)01048-6
- P. Di Simplicio, F. Franconi, S. Frosali and D. Di Giuseppe, “Thiolation and Nitrosation of Cysteines in Biological Fluids and Cells,” Amino Acids, Vol. 25, No. 3-4, 2003, pp. 323-339. http://dx.doi.org/10.1007/s00726-003-0020-1
- Y. M. Janssen-Heininger, B. T. Mossman, N. H. Heintz, H. J. Forman, B. Kalyanaraman, T. Finkel, J. S. Stamler, S. G. Rhee and A. van der Vliet, “Redox-Based Regulation of Signal Transduction: Principles, Pitfalls, and Promises,” Free Radical Biology & Medicine, Vol. 45, No. 1, 2008, pp. 1-17. http://dx.doi.org/10.1016/j.freeradbiomed.2008.03.011
- R. Priora, L. Coppo, A. Margaritis, D. Di Giuseppe, S. Frosali, D. Summa, J. Heo and P. Di Simplicio, “The Control of S-Thiolation by Cysteine via Gamma-Glutamyltranspeptidase and Thiol Exchanges in Erythrocytes and Plasma of Diamide-Treated Rats,” Toxicology and Applied Pharmacology, Vol. 242, No. 3, 2010, pp. 333-343. http://dx.doi.org/10.1016/j.taap.2009.11.003
- B. D’Autreaux and M. B. Toledano, “ROS as Signalling Molecules: Mechanisms That Generate Specificity in ROS Homeostasis,” Nature Reviews Molecular Cell Biology, Vol. 8, No. 10, 2007, pp. 813-824. http://dx.doi.org/10.1038/nrm2256
- J. P. Brennan, R. Wait, S. Begum, J. R. Bell, M. J. Dunn and P. Eaton, “Detection and Mapping of Widespread Intermolecular Protein Disulfide Formation during Cardiac Oxidative Stress Using Proteomics with Diagonal Electrophoresis,” Journal of Biological Chemistry, Vol. 279, No. 40, 2004, pp. 41352-41360. http://dx.doi.org/10.1074/jbc.M403827200
- V. M. Chen and P. J. Hogg, “Allosteric Disulfide Bonds in Thrombosis and Thrombolysis,” Journal of Thrombosis and Haemostasis, Vol. 4, No. 12, 2006, pp. 2533-2541. http://dx.doi.org/10.1111/j.1538-7836.2006.02236.x
- R. C. Cumming, N. L. Andon, P. A. Haynes, M. ark, W. H. Fischer and D. Schubert, “Protein Disulfide Bond Formation in the Cytoplasm during Oxidative Stress,” Journal of Biological Chemistry, Vol. 279, No. 21, 2004, pp. 21749-21758. http://dx.doi.org/10.1074/jbc.M312267200
- B. Schmidt, L. Ho and P. J. Hogg, “Allosteric Disulfide Bonds,” Biochemistry, Vol. 45, No. 24, 2006, pp. 7429- 7433. http://dx.doi.org/10.1021/bi0603064
- J. Ahamed, H. H. Versteeg, M. Kerver, V. M. Chen, B. M. Mueller, P. J. Hogg and W. Ruf, “Disulfide Isomerization Switches Tissue Factor from Coagulation to Cell Signaling,” Proceedings of the National Academy of Sciences of the United States of America, Vol. 103, No. 38, 2006, pp. 13932-13937. http://dx.doi.org/10.1073/pnas.0606411103
- A. Margaritis, R. Priora, S. Frosali, D. Di Giuseppe, D. Summa, L. Coppo, A. Di Stefano and P. Di Simplicio, “The Role of Protein Sulfhydryl Groups and Protein Disulfides of the Platelet Surface in Aggregation Processes Involving Thiol Exchange Reactions,” Pharmacological Research, Vol. 63, No. 1, 2011, pp. 77-84. http://dx.doi.org/10.1016/j.phrs.2010.09.004
- L.V. Bock, B. Hutchings, H. Grubmuller and D. J. Woodbury, “Chemomechanical Regulation of SNARE Proteins Studied with Molecular Dynamics Simulations,” Biophysical Journal, Vol. 99, No. 4, 2010, pp. 1221-1230. http://dx.doi.org/10.1016/j.bpj.2010.06.019
- K. Krapfenbauer, E. Engidawork, N. Cairns, M. Fountoulakis and G. Lubec, “Aberrant Expression of Peroxiredoxin Subtypes in Neurodegenerative Disorders,” Brain Research, Vol. 967, No. 1-2, 2003, pp. 152-160. http://dx.doi.org/10.1016/S0006-8993(02)04243-9
- F. Hattori and S. Oikawa, “Peroxiredoxins in the Central Nervous System,” Subcellular Biochemistry, Vol. 44, 2007, pp. 357-374. http://dx.doi.org/10.1007/978-1-4020-6051-9_17
- S. F. Newman, R. Sultana, M. Pierluigi, R. Coccia, J. Cai, V. M. Pierce, J. B. Klein, D. M. Turner and D. A. Butterfield, “An Increase in S-Glutathionylated Proteins in the Alzheimer’s Disease Inferior Parietal Lobule, a Proteomics Approach,” Journal of Neuroscience, Vol. 85, 2007, pp. 1506-1514.
- R. C. Cumming and D. Schubert, “Amyloid-Beta Induces Disulfide Bonding and Aggregation of GAPDH in Alzheimer’s Disease,” The FASEB Journal, Vol. 19, No. 14, 2005, pp. 2060-2062.
- I. S. Kil and J. W. Park, “Regulation of Mitochondrial NADP+-Dependent Isocitrate Dehydrogenase Activity by Glutathionylation,” Journal of Biological Chemistry, Vol. 280, No. 11, 2005, pp. 10846-19854. http://dx.doi.org/10.1074/jbc.M411306200
- D. Summa, O. Spiga, A. Bernini, V. Venditti, R. Priora, S. Frosali, A. Margaritis, D. Di Giuseppe, N. Niccolai and P. Di Simplicio, “Protein-Thiol Substitution or Protein Dethiolation by Thiol/Disulfide Exchange Reactions: The Albumin Model,” Proteins, Vol. 69, No. 2, 2007, pp. 369- 378. http://dx.doi.org/10.1002/prot.21532
- O. Spiga, D. Summa, S. Cirri, A. Bernini, V. Venditti, M. De Chiara, R. Priora, S. Frosali, A. Margaritis, D. Di Giuseppe, P. Di Simplicio and N. Niccolai, “A Structurally Driven Analysis of Thiol Reactivity in Mammalian Albumins,” Biopolymers, Vol. 95, No. 4, 2011, pp. 278-285. http://dx.doi.org/10.1002/prot.21532
- C. Kumsta and U. Jacob, “Redox-Regulated Chaperones,” Biochemistry, Vol. 48, No. 22, 2009, pp. 4666-4676. http://dx.doi.org/10.1021/bi9003556
- K. Liberek, A. Lewandowska and S. Zietkiewicz, “Chaperones in Control of Protein Disaggregation,” The EMBO Journal, Vol. 27, 2008, pp. 328-335. http://dx.doi.org/10.1038/sj.emboj.7601970
- M. Fratelli, H. Demol, M. Puype, S. Casagrande, I. Eberini, M. Salmona, V. Bonetto, M. Mengozzi, F. Duffieux, E. Miclet, A. Bachi, J. Vandekerckhove, E. Gianazza and P. Ghezzi, “Identification by Redox Proteomics of Glutathionylated Proteins in Oxidatively Stressed Human T Lymphocytes,” Proceedings of the National Academy of Sciences of the United States of America, Vol. 99, No. 6, 2002, pp. 3505-3510. http://dx.doi.org/10.1073/pnas.052592699
- P. Ghezzi and P. Di Simplicio, “Glutathionylation Pathways in Drug Response,” Current Opinion in Pharmacology, Vol. 7, No. 4, 2007, pp. 398-403. http://dx.doi.org/10.1016/j.coph.2007.04.006
- C. C. Winterbourn and M. B. Hampton, “Thiol Chemistry and Specificity in Redox Signaling,” Free Radical Biology and Medicine, Vol. 45, No. 5, 2008, pp. 549-561. http://dx.doi.org/10.1016/j.freeradbiomed.2008.05.004
- P. J. Hogg, “Disulfide Bonds as Switches for Protein Function,” Trends in Biochemical Sciences, Vol. 28, No. 4, 2003, pp. 210-214. http://dx.doi.org/10.1016/S0968-0004(03)00057-4
- A. Holmgren, “Thioredoxin and Glutaredoxin Systems,” Journal of Biological Chemistry, Vol. 264, No. 24, 1989, pp. 13963-13966.
- H. Nakamura, “Thioredoxin and Its Related Molecules: Update 2005,” Antioxidants and Redox Signaling, Vol. 7, No. 5-6, 2005, pp. 823-828. http://dx.doi.org/10.1089/ars.2005.7.823
- G. L. Ellman, “Tissue Sulfhydryl Groups,” Archives of Biochemistry and Biophysics, Vol. 82, No. 2, 1959, pp. 70-77. http://dx.doi.org/10.1016/0003-9861(59)90090-6
- R. Priora, L. Coppo, S. Salzano, P. Di Simplicio and P. Ghezzi, “Measurement of Mixed Disulfides Including Glutathionylated Proteins,” Methods in Enzymology, Vol. 473, 2010, pp. 149-159. http://dx.doi.org/10.1016/S0076-6879(10)73007-X
- N. S. Kosower and E. M. Kosower, “Diamide: An Oxidant Probe for Thiols,” Methods in Enzymology, Vol. 251, 1995, pp. 123-133. http://dx.doi.org/10.1016/0076-6879(95)51116-4
- L. Nagy, M. Nagata and S. Szabo, “Protein and NonProtein Sulfhydryls and Disulfides in Gastric Mucosa and Liver after Gastrotoxic Chemicals and Sucralfate: Possible New Targets of Pharmacologic Agents,” World Journal of Gastroenterology, Vol. 13, No. 14, 2007, pp. 2053-2060.
- N. N. Gessler, M. N. Rudchenko and T. A. Belozerskaia, “Stress Factor-Induced Changes in the Activity of Antioxidant Protective Mechanisms in the Wild Type Strain of Neurospora Crassa and in Its Photoreceptor Complex Mutants,” Mikrobiologiia, Vol. 77, 2008, pp. 163-170.
- R. E. Hansen, D. Roth and J. R. Winther, “Quantifying the Global Cellular Thiol-Disulfide Status,” Proceedings of the National Academy of Sciences of the United States of America, Vol. 106, No. 2, 2009, pp. 422-427. http://dx.doi.org/10.1073/pnas.0812149106
- R. Rossi, E. Cardaioli, A. Scaloni, G. Amiconi and P. Di Simplicio, “Thiol Groups in Proteins as Endogenous Reductants to Determine Glutathione-Protein Mixed Disulfides in Biological Systems,” Biochimica et Biophysica Acta, Vol. 1243, No. 2, 1995, pp. 230-238. http://dx.doi.org/10.1016/0304-4165(94)00133-I
- L. O. Andersson, “Hydrolysis of Disulfide Bonds in Weakly Alkaline Media. II. Bovine Serum Albumin Dimer,” Biochimica et Biophysica Acta, Vol. 200, No. 2, 1970, pp. 363-369. http://dx.doi.org/10.1016/0005-2795(70)90178-9
- P. A. Fernandes and M. J. Ramos, “Theoretical Insights into the Mechanism for Thiol/Disulfide Exchange,” Chemistry, Vol. 10, No. 1, 2004, pp. 257-266. http://dx.doi.org/10.1002/chem.200305343
- P. Di Simplicio, M. G. Cacace, L. Lusini, F. Giannerini, D. Giustarini and R. Rossi, “Role of Protein-SH Groups in Redox Homeostasis the Erythrocyte as a Model System,” Archives of Biochemistry and Biophysics, Vol. 355, No. 2, 1998, pp. 145-152. http://dx.doi.org/10.1006/abbi.1998.0694
- P. Di Simplicio, F. Giannerini, D. Giustarini, L. Lusini and R. Rossi, “The Role of Cysteine in the Regulation of Blood Glutathione-Protein Mixed Disulfides in Rats Treated with Diamide,” Toxicology and Applied Pharmacology, Vol. 148, No. 1, 1998, pp. 56-64. http://dx.doi.org/10.1006/taap.1997.8305
- R. Rossi, D. Barra, A. Bellelli, G. Boumis, S. Canofeni, P. Di Simplicio, L. Lusini, S. Pascarella and G. Amiconi, “Fast-Reacting Thiols in Rat Hemoglobins Can Intercept Damaging Species in Erythrocytes more Efficiently than Glutathione,” The Journal of Biological Chemistry, Vol. 273, 1998, pp. 19198-19206. http://dx.doi.org/10.1074/jbc.273.30.19198
- P. Di Simplicio, L. Lusini, F. Giannerini, D. Giustarini, A. Bellelli, G. Boumis, G. Amiconi and R. Rossi, “Nitric Oxide and the Cell,” In: S. Moncada, G. Nisticò, G. Bagetta and E. A. Higgs, Eds., Proliferation Differentiation and Death, Portland Press, Colchester, 1998, pp 47- 59.
- P. J. Pomposiello and B. Demple, “Redox-Operated Genetic Switches: The SoxR and OxyR Transcription Factors,” Trends in Biotechnology, Vol. 19, No. 3, 2001, pp. 109-114. http://dx.doi.org/10.1016/S0167-7799(00)01542-0
- C. Gitler, B. Zarmi and E. Kalef, “General Method to Identify and Enrich Vicinal Thiol Proteins Present in Intact Cells in the Oxidized, Disulfide State,” Analytical Biochemistry, Vol. 252, No. 1, 1997, pp. 48-55. http://dx.doi.org/10.1006/abio.1997.2294
- R. Franco and J. A. Cidlowski, “Apoptosis and Glutathione: Beyond an Antioxidant,” Cell Death and Differentiation, Vol. 16, 2009, pp. 1303-1314. http://dx.doi.org/10.1038/cdd.2009.107
NOTES
*Corresponding author.