Open Journal of Soil Science
Vol.3 No.7(2013), Article ID:39644,17 pages DOI:10.4236/ojss.2013.37035
Assessment of Soil C and N Stocks and Fractions across 11 European Soils under Varying Land Uses
![]()
1Natural Resource Ecology Laboratory, Colorado State University, Fort Collins, Colorado, USA; 2Environmental Science Department, Second University of Naples, Caserta, Italia; 3Department of Soil and Crop Sciences, Colorado State University, Fort Collins, Colorado, USA.
Email: *Karolien.Denef@Colostate.edu
Copyright © 2013 Karolien Denef et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received August 6th, 2013; revised September 6th, 2013; accepted September 11th, 2013
Keywords: Soil organic carbon; soil nitrogen; soil organic matter fractions; natural 15N and 13C abundance
ABSTRACT
In this study, we measured the stocks and pool sizes of soil organic carbon (SOC) and total soil nitrogen (TN), and their natural 13C and 15N abundance across a wide range of temperate European ecosystems. The objectives were to examine any distinct isotope patterns with land use or climate, and how C and N in these different ecosystems are distributed among soil organic matter (SOM) fractions to better predict soil C and N dynamics and longer term persistence. Soils were sampled to 30 cm depth at 11 sites of the Nitro Europe (NEU) network and included four forests, three grasslands and four croplands. Surface soil samples were fractionated using a combined size-density fractionation protocol separating light (LF) from heavy particulate organic matter (hPOM) by density and silt-from-clay-associated SOM by size. Down-profile natural abundance 15N patterns pointed towards a closed N cycle in the forest sites, while 13C patterns suggested differences in plant water use efficiency across the C3 grassland sites. The forests and grassland sites stored the majority of surface SOC and TN in the LF and hPOM pools. Sustained sequestration of C and N in these rather labile pools will rely on management practices that minimize soil disturbance and increase C input. We also found that the mineral fraction (silt and clay) in the cropland soils stored less C and N per unit of fraction mass compared to the forests and grasslands, which points towards a lower mineral-OM stabilization efficiency of cropland soils. Finally, our study revealed total POM (LF plus hPOM) as a strong predictor of SOC and TN differences, particularly among the non-cropped sites. This study shows that these sites, independent of soil type and climate, store a large fraction of C and N in POM pools that are particularly vulnerable to soil disturbance such as caused by land use change.
1. Introduction
Globally, soils store a tremendous amount of organic carbon (~2300 Pg C) and nitrogen (~140 Pg N) [1], of which the majority is contained in the soil organic matter (SOM). The benefits of SOM for soil fertility are widely recognized due to its contribution to soil structure and aeration, nutrient and water holding capacity. The organic matter content of a soil reflects the balance between plant, animal, microbial and erosional inputs, and losses due to mineralization, leaching and erosion. It is therefore a dynamic property, greatly vulnerable to land use and climate [2] and with important feedbacks to the atmospheric green house gas (GHG) balance and the rate of climate change [3]. The increased demand for accurate soil C and N stock assessments and predictions of C and N changes as a result of land use/cover and climate change has triggered large-scale and long-term measurements of soil C and N stocks and pools globally [4-6]. Such empirical studies provide critical information to quantify the response of soil properties to management and changing climate across regions, ultimately supporting climate change and farm policy, food security, and overall ecosystem health. Monitoring soil C and N stocks is imperative not only for understanding how soils change in response to land use and shifting climate patterns, but also for validating and reducing the uncertainty around estimates of biogeochemical prediction models as Century [7], DayCent [8], Roth C [9] and DNDC [10] commonly used in regional and national GHG inventories. Model outputs typically come with large uncertainties [11,12], which is at least partly due to the still limited knowledge about ecosystem processes and properties involved in C and N cycling [13]. Consequently, direct measurements by repeated soil inventories are urgently needed to improve our understanding of C and N dynamics and further constrain model estimates [6,12].
Whole soil C and N stock changes are difficult to detect in the short term as the bulk of SOM is stabilized and has turnover times measured in hundreds to thousands of years [14]. This would require long term measurements to capture modest changes in a robust manner [15]. Soil organic matter fractionations can be helpful in studying long-term C and N dynamics, as they can reveal early indications of changes in SOC and TN stocks [16-19] hence predict how ecosystems will respond to a particular change. A number of physical fractionation methods have been successful in terms of isolating more active dissolved (DOM) and particulate (POM) organic matter pools that are likely to participate in short term soil processes (e.g. nutrient cycling) and respond more rapidly to land use or management changes, from more passive mineral-associated SOM pools that are relatively recalcitrant and more resistant to disturbance [20-22]. Structural analyses have revealed unique molecular characteristics of these SOM fractions based on their degree of decomposition, the dominance of different chemical classes, and the relative contributions from plant and microbially-derived C linking their physical characteristics (i.e. how they are operationally defined) to their ecological function [23]. Particulate organic matter fractions, obtained either by particle size (often >53 μm) or density (usually 1.3 - 1.8 g cm−3) separation, are composed primarily of identifiable and young plant litter in various stages of decomposition that is chemically similar to its source material, whereas SOM associated with silt and clay minerals (usually < 53 μm), occluded within aggregates, or associated in higher density organo-mineral complexes, is generally further decomposed and more microbially altered [24,25], containing a greater proportion of microbial products and more resistant C as compared with larger or lighter fractions [26]. Assessing how the bulk of SOM in different ecosystems is distributed among these ecologically meaningful fractions can provide insights into SOM behavior upon management or land use change. This will ultimately help decision-making on best management practices to maintain or increase soil C and N. For example, Stewart et al. [21] stressed the importance of conservation management (reduced disturbance and increased C input) in the Virginia Coastal Plains to avoid rapid losses of SOC, as most of the change in SOC upon changes in management was observed in the active POM pools.
Soil 13C and 15N natural abundance measurements are another useful tool forinferring changes in C and Ndynamics and reducing uncertainty in estimates of soil C storage. The natural abundance of 13C and 15N in SOM has been shown to be an integrative measure of the ecosystem factors and processes that produced it, and its measurement has yielded insight into the dominant processes guiding the biological and physical dynamics in a soil system [e.g., 27]. Soil 13C values are closely related to vegetation cover (C3 versus C4 plants) and moisture availability. They can inform about historic vegetation or climatic shifts, and can be used to measure SOC turnover [28]. Mineralization and processes associated with SOM formation induce additional variations in the soil13C abundance, contributing to the often observed 13C enrichment with soil depth [29,30]. Soil 15N values reflect soil N sources and their fractionation during N transformation [31,32]. Generally, closed N cycles (i.e. limited N losses) are expressed by 15N patterns with lower 15N at the soil surface and relatively higher 15N with depth, as has been found in temperate forests not subject to significant N inputs, whereas 15N enriched soils suggest open N cycles with greater N losses via fractionating pathways such as nitrification and denitrification [31,32]. Nitrogen fertilizer or atmospheric deposition inputs may complicate the interpretation of natural abundance 15N gradients in soil [33].
The aim of our investigation was to assess how C and N in different ecosystems are distributed among SOM fractions to predict longer term persistence and SOM behavior when subject to disturbance, such as changes in climate, management or land use. To do this we 1) assessed SOC and TN stocks as well as 13C and 15N natural abundance of 11 soil profiles (0 - 0 cm) across Europe, and 2) evaluated the distribution and variability of SOC and TN among SOM fractions in these temperate soils of varying land uses. Land use affects SOC and TN stocks, their distribution among SOM fractions and their 13C and 15N values across the soil profile. Our hypothesis was that forests and grasslands would store more C and N in the more labile POM fractions compared to croplands. We also expected a more uniform 13C and 15N pattern across the soil profile of the cropland sites due to tillage, and for the forest sites, a 15N pattern indicative of a closed N cycle, with depleted 15N values in the surface soils as compared to 15N values at deeper depths.
2. Materials and Methods
2.1. Site Description
Soil cores were collected from 11 sites across Europe during spring and summer of 2007 representing a variety of climates, land-use and soil properties. The sites cover different climatological zones (Figure 1) from Finland in the north to Italy in the south, and from the UK in the west to Hungary in the east [34,35] and are part of the Nitro Europe Level-3 “Super Sites” network. Addition-
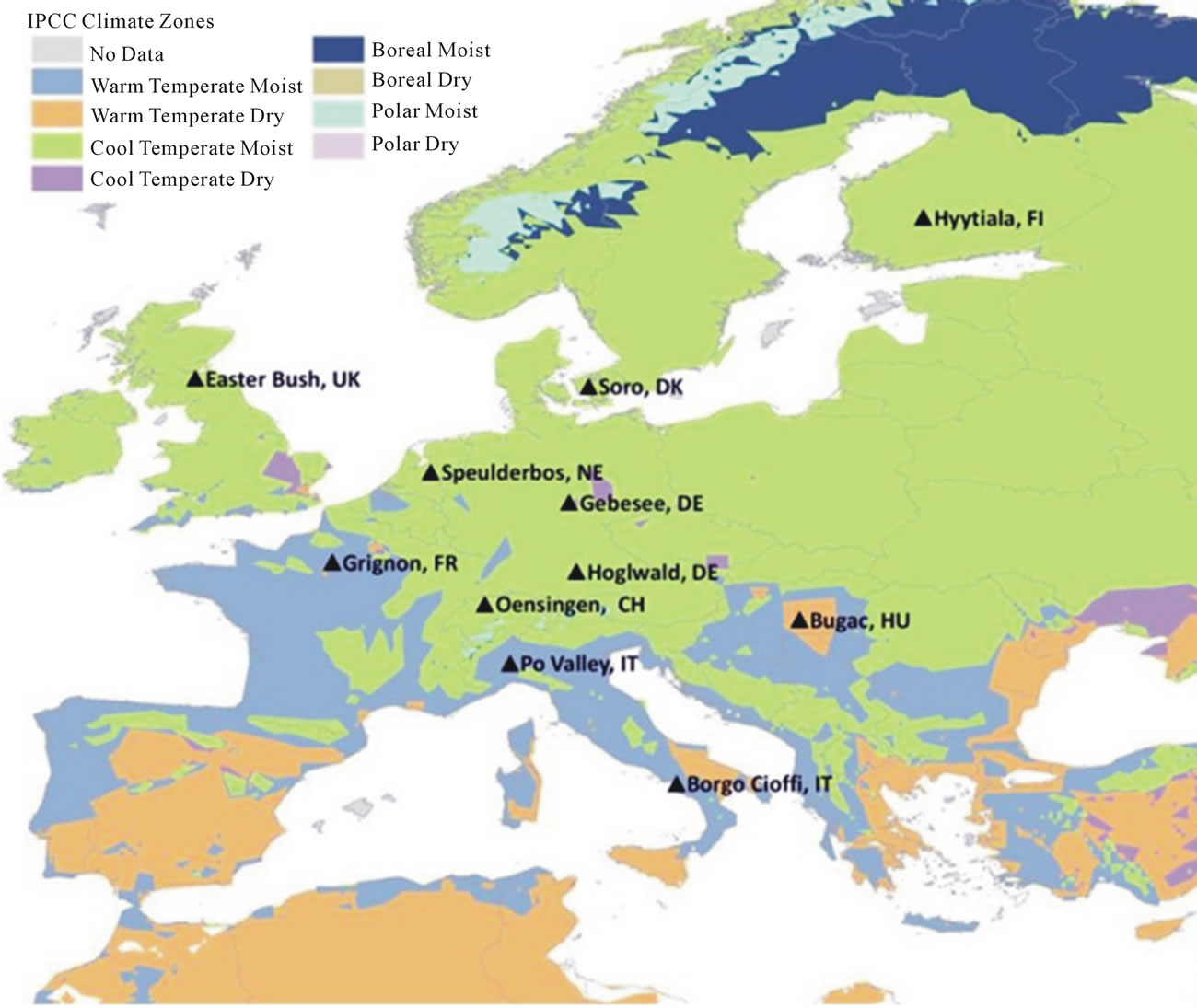
Figure 1. Map with location of sampled sites and IPCC climate zones.
ally, the sites also differ in terms of vegetation, soil properties (e.g. bulk density, texture, pH) and land use, including four forests, three grass lands and four croplands. All croplands and grasslands, except for the grassland in Bugac, are fertilized with different amounts of fertilizer. Grasslands are either grazed or mowed. Forest sites differ in tree species of coniferous or deciduous type. The main site characteristics are summarized in Table 1. Additional information can be found in Schaufler et al. [36] and Skiba et al. [35].
2.2. Soil Sampling and Preparation
To allow for data comparison and integration with other soil and gas measurements at the NEU Level-3 “Super Site” network [35], soil samples were collected in proximity to the six replicate flux measurement chambers which were randomly placed at each site (for details, see [37]). Around each chamber, after removing the litter layer (where present), one soil core was taken at each of the four corners of a 16 m2 area to a depth of 30 cm with a 5 cm diameter sample probe. Each of the four soil cores was split into four depths (0 - 5 cm, 5 - 10 cm, 10 - 20 cm and 20 - 30 cm) and composited by depth, resulting in one composite sample for each depth for each replicate area (n = 6) per site. Soil samples of all depths were sieved to 2 mm and oven-dried (105˚C) prior to further analyses. One additional soil core was taken at each sampling location for bulk density measurement. Bulk density was measured for each of the four depths, as the ratio of the dry (105˚C) weight of the soil within that depth divided by the volume, measured from the diameter and height of the core, after correcting for any rocks or coarse fragments. No significant core compaction was observed during sampling.
2.3. Bulk Soil Analyses
Oven-dried soil samples were finely ground and analyzed for SOC, TN and isotopic composition (δ13C [‰] and
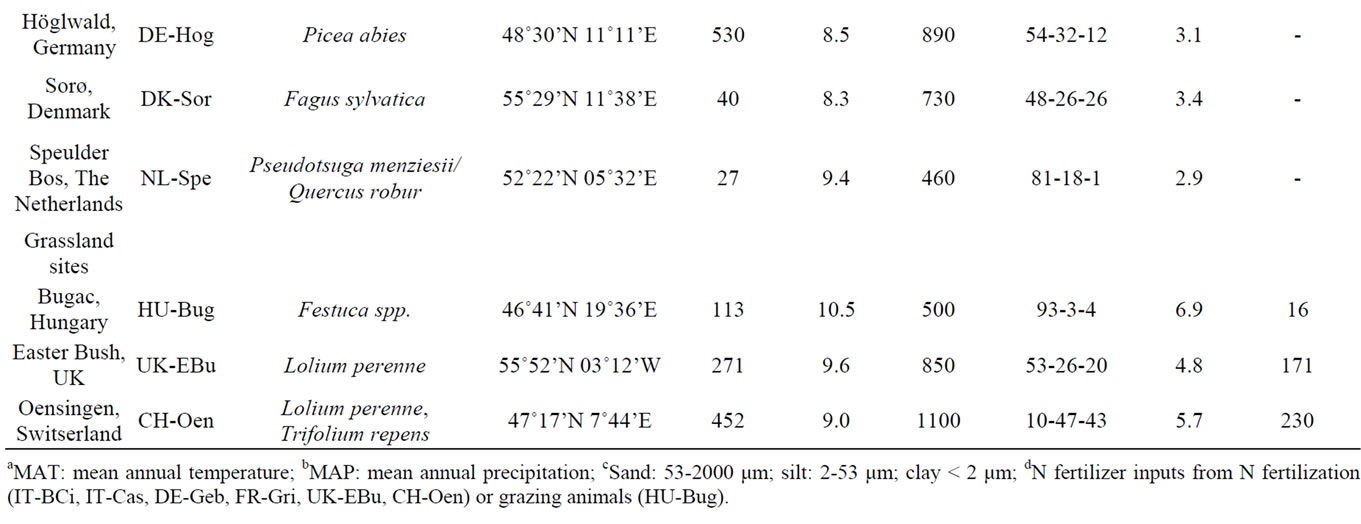
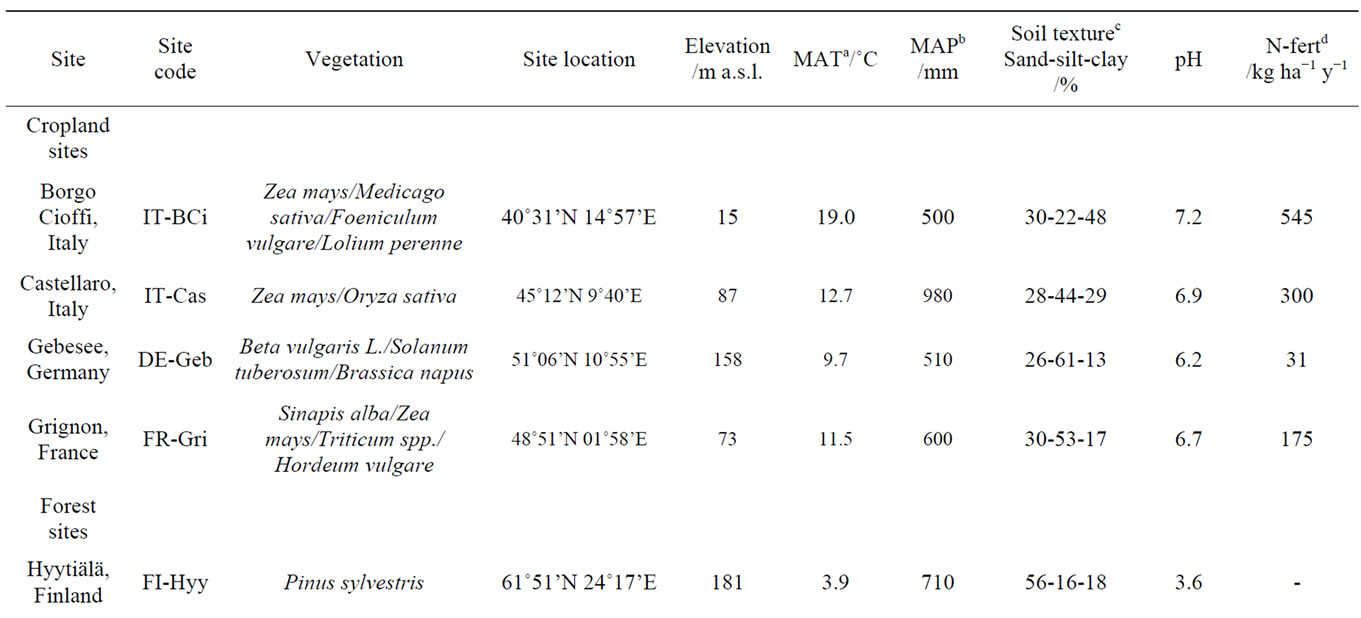
Table 1. Sampling sites with information on land use, vegetation, location, climate and soil characteristics.
δ15N [‰]) using an elemental analyzer (EA) (Thermo Finnigan: EA 1112) coupled with an isotope ratio mass spectrometer (IRMS) (Thermo Finnigan: Deltaplus). Carbonates were removed prior to EA and IRMS analysis by exposure to HCl vapor for 6 hours [38].
2.4. Soil Fractionation
All surface samples (0 - 5 cm) were fractionated using a combined size-density fractionation protocol, adopted from Marzaioli et al. [39]. This method separates total light fraction (LF) from heavy particulate organic matter (hPOM) by density separation and silt-from clay-associated SOM by size. In brief, 2 mm sieved soil samples underwent a density separation in sodium polytungstate (SPT; ρ = 1.85 g cm−3). This step allows the separation of LF (floats in the SPT) from heavy fraction (settles out in the SPT). The heavy fraction, which is composed of aggregates, sand and POM, was dispersed by glassbeads in de-ionized water, and wet sieved on a 53 μm mesh sieve, to separate the hPOM plus sand fraction (retained on the 53 μm screen) from the silt plus clay fraction. Silt and clay sized fractions were separated by means of wet centrifugation (127 g for 7 min for silt, and 1730 g for 15 min for clay following addition of flocculant, i.e. 0.25 M CaCl2-MgCl2). All recovered fractions were analyzed for SOC and TN after carbonate removal [38], as described above.
2.5. Statistical Analyses
To test the assumption of normality, we examined QQ plots of the quantiles of studentized residuals versus standard normal quantiles. Soil organic C and TN stocks and concentrations were root-transformed to achieve normality. Statistical tests were performed using transformed data, but non transformed values were used to report average values in tables and figures. The effects of land use, climate (mean annual temperature (MAT) and precipitation (MAP)), soil properties (pH, texture) and their interactions on SOC and TN stocks were analyzed using the ANCOVA model with the proc mixed module in SAS 9.2 (SAS Institute). The effects of land use on fraction C and N content were analyzed using the ANOVA model with the proc mixed module. The variable “site within land use” was considered a random effect in all analyses to resolve non-independencies and to account for the bysite variation. Separation of means was tested with Tukey’s honestly significant difference at a significance level of 0.05. Linear relationships between total SOC and TN content and individual C and N fractions, isolated by the different fractionation methods, were tested using ordinary least squares linear regression.
3. Results
3.1. Bulk Soil Organic Carbon and Total Nitrogen
Soil organic C and TN stocks in the 0 - 30 cm depth ranged between 3.80 and 11.19 kgm−2 for SOC and 0.17 and 0.90 kgm−2 for TN and differed greatly among sites, even within a land use type (Table 2; see also Tables 3 and 4 for individual depths). The highest SOC and TN stocks were found at the Bugac grassland site and Sorö forest site, respectively, while the lowest SOC and TN stocks were found at the Borgo Cioffi cropland site and Hyytiälä forest site, respectively. Over the entire 30 cm depth, SOC and TN stocks did not significantly differ with land use, climate (MAT, MAP), soil properties (pH, texture) or their interactions.
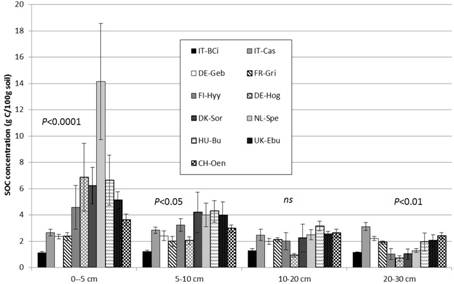
Significant differences between land use types were observed for SOC and TN concentrations at the different depths (Figure 2; Table 5). The SOC and TN concentration in the surface 0 - 5 cm depth was significantly lower in cropland sites compared to grassland (SOC: P = 0.0085; TN: P = 0.05) and forest sites (SOC: P < 0.0001). At the deepest depth (20 - 30 cm), TOC and TN concentrations were significantly lower in the forest sites compared to grassland and cropland sites (P < 0.05). SOC and TN concentration significantly decreased with depth in the grassland and forest sites (P < 0.0001), but not in the cropland sites (Figure 2).
3.2. Bulk Soil 13C and 15N
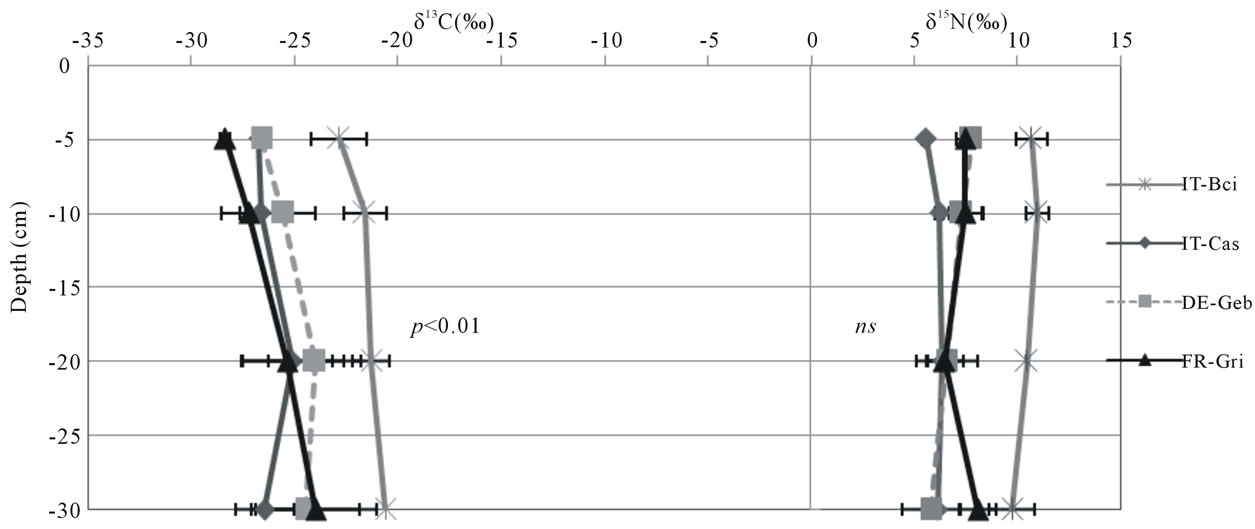
Delta 13C values ranged between −29.9‰ and −20.6‰, whereas δ15N values ranged between −2.1‰ and 11.0‰ (Figure 3). The highest δ13C and δ15N values were found at the Borgo Cioffi cropland site, while the lowest δ13C and δ15N values were found at the Hyytiälä and Speulderbos forest sites, respectively. At all sites, bulk soil became enriched in 13C up to several parts permil with depth (average increase 2.4‰). Similar results were found for 15N in grassland (as high as 3.6‰ increase with depth) and forest sites (as high as 10.4‰ increase with depth), but for cropland sites, the 15N signature of the soil did not significantly change with depth (Figure 3). Land use significantly affected the 15N (P = 0.0147) but not the 13C signature of the bulk soil (Table 5). The land use effect on 15N was dependent on depth, with forest soils being more depleted in 15N in the 0-5 cm depth (as low as −2.1‰ at Speulderbos) compared to the cropland (P < 0.0001) and grassland sites (P = 0.0079). At deeper depths, no significant differences among land use types were found in the isotopic composition of the SOC and TN.
3.3. Carbon and Nitrogen Content of Soil Organic Matter Fractions
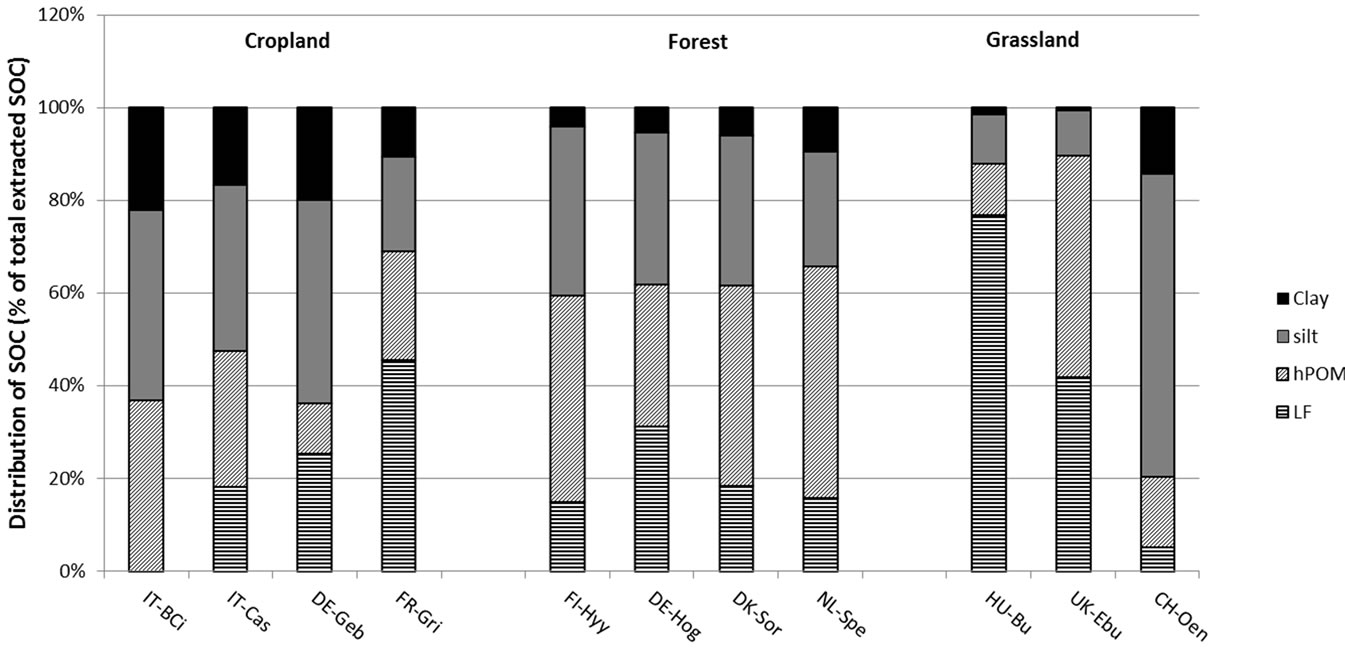
Soil organic C and TN were differently distributed among the isolated SOM fractions at the different sites (Figure 4). In the majority of forest and grassland soils, total POM (LF plus hPOM) accounted for most of the SOC and TN. Over 60% of SOC and TN were found in total POM in the forest soils at Hyytiälä, Hogwald, Sorö and Speulderbos, and even over 90% were found in the grassland soils at Bugac and Easter Bush (Figure 4). The grassland site at Oensingen, on the other hand, had the majority of SOC and TN (80%) stored in the mineral fractions (silt and clay). Cropland soils stored most of their SOC and TN in the mineral fractions, with the exception of the cropland soil at Grignon, which stored nearly 70% of SOC and TN in its POM fractions (Figure 4).
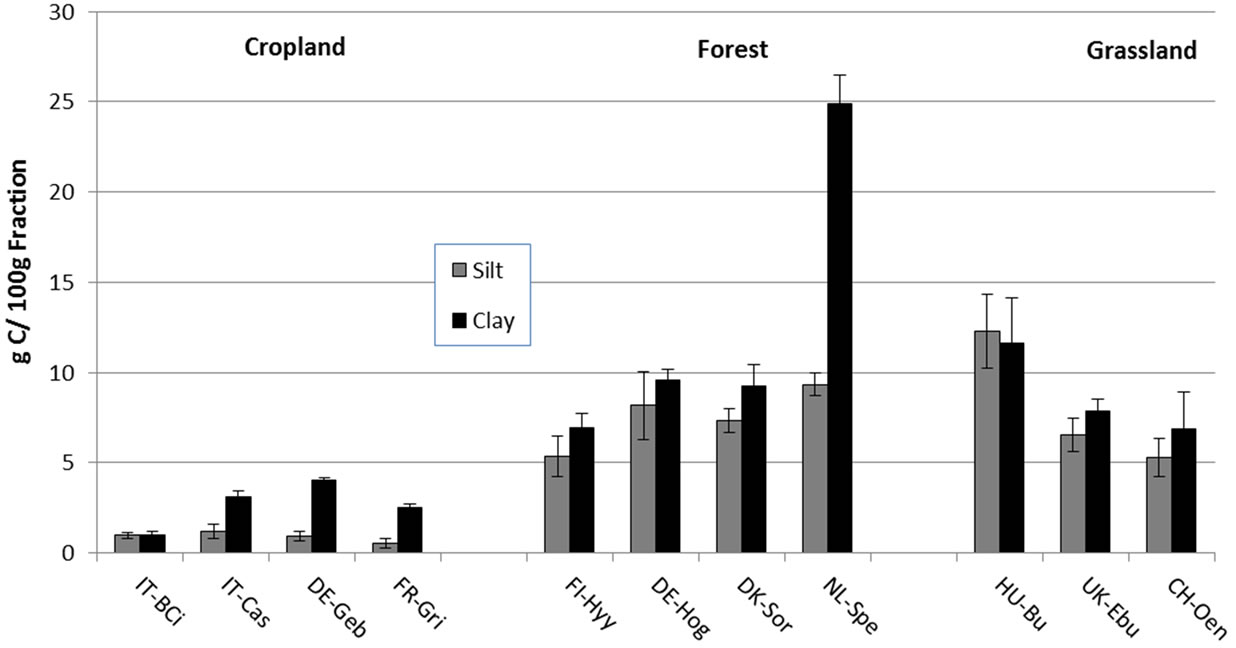
When fraction C and N were expressed on a per fraction mass basis, the silt and clay fractions were significantly less enriched in SOC and TN in all the cropland sites (Borgo Cioffi, Castellaro, Gebesee and Grignon) compared to the grassland and forest sites (Figure 5; Table 5). At most sites, more SOC and TN were associated with clay compared to silt minerals (Figure 5).
3.4. Relationship between Soil Organic Matter Fractions and Total Carbon and Nitrogen
In order to evaluate the potential of the isolated SOM fractions for predicting SOC and TN changes, we correlated each individual fraction’s C and N content against the SOC and TN calculated combining all fractions. A significant linear relationship was found between SOC (and TN) and total POM (LF plus hPOM) C (and N) when all sites were included as well as within a land use type, with the predictive strength (R2) being much greater for the non-cropped sites (R2 > 0.85) compared to the croplands (R2 = 0.46 for TN; R2 = 0.71 for TC) (Table 6). No reasonable relationship was observed when correlating total SOC and TN with any of the other fractions (Table 6), suggesting that total POM is the largest con
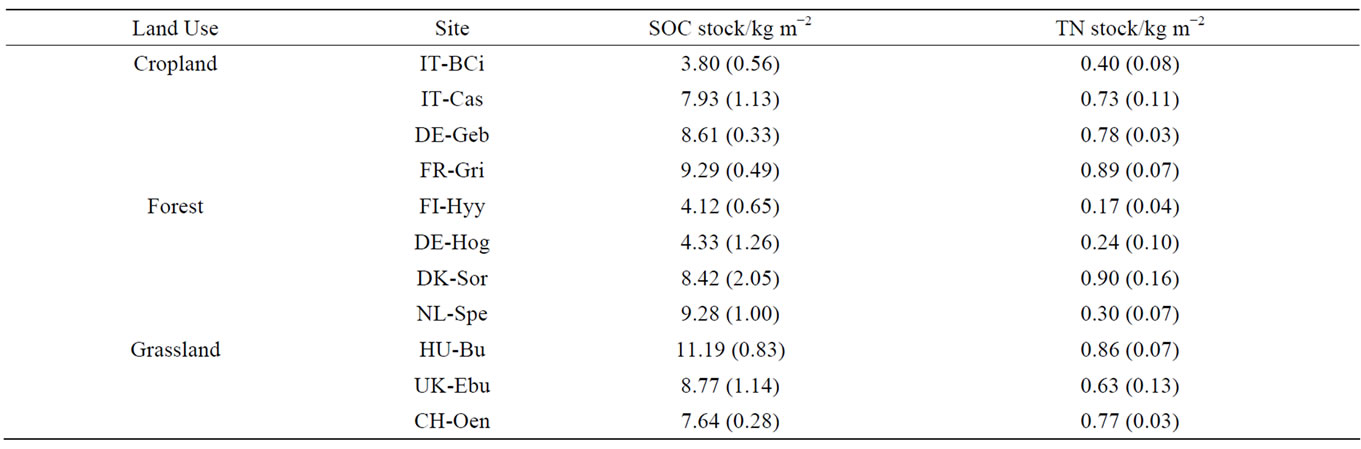
Table 2. Total 0 - 30 cm soil organic carbon (SOC) and nitrogen (TN) stocks (kg m−2). Values represent averages of 6 field replicates, with standard deviation in brackets.
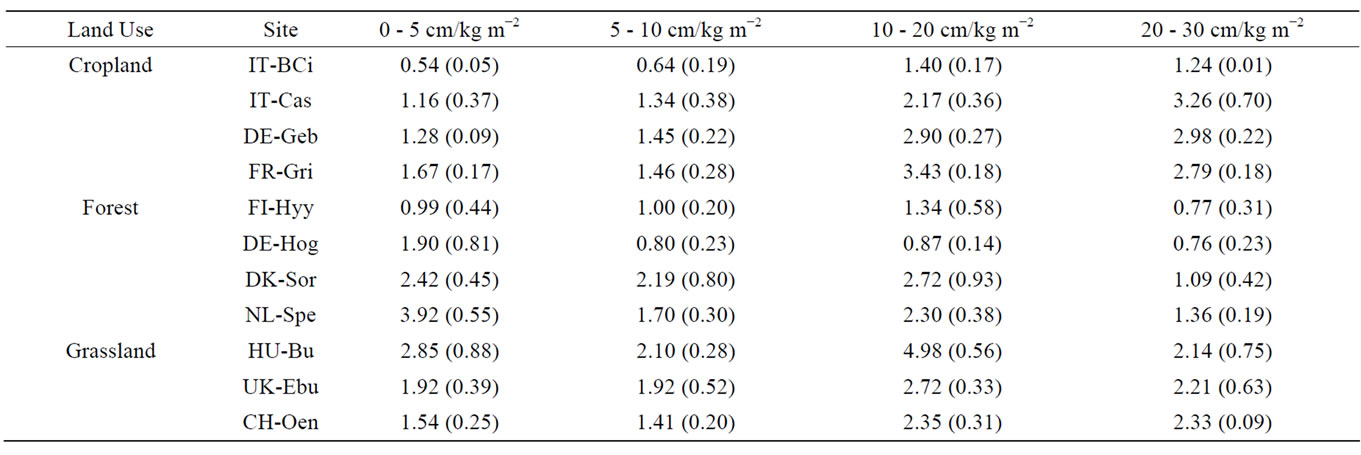
Table 3. Soil organic carbon (SOC) stocks (kg m−2) at 0 - 5 cm, 5 - 10 cm, 10 - 20 cm and 20 - 30 cm depths. Values represent averages of 6 field replicates within a site, with standard deviation in brackets.
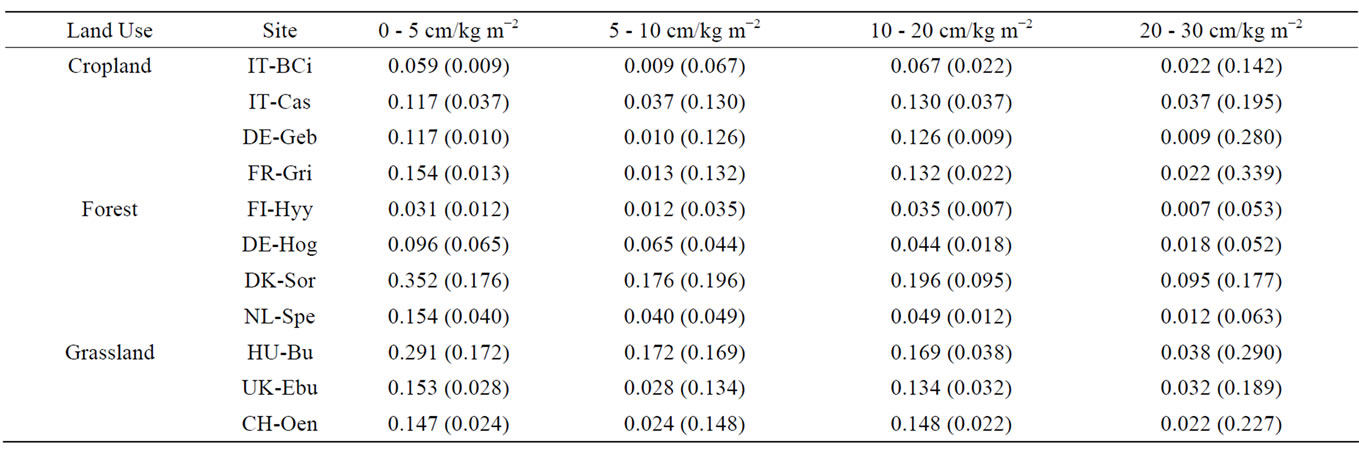
Table 4. Total soil nitrogen (TN) stocks (kg m−2) at 0 - 5 cm, 5 - 10 cm, 10 - 20 cm and 20 - 30 cm depths. Values represent averages of 6 field replicates within a site, with standard deviation in brackets.
tributor to surface soil C and N differences across the different sites.
4. Discussion
Recent research efforts supported by the European Union (e.g., CarboEurope and NitroEurope) have initiated large scale assessments of the C and N fluxes from soils across Europe. Our investigation contributes to this effort by providing SOC and TN stocks for 11 NitroEurope flux measurement sites across Europe [34], and allows for a more in-depth evaluation of how soil C and N is stabilized in these systems by evaluating the distribution of C
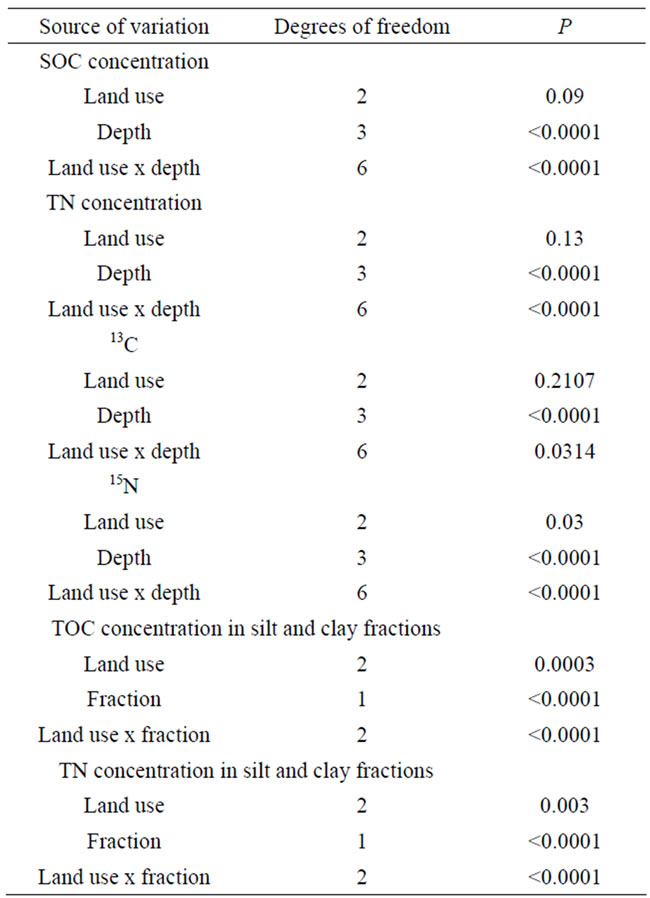
Table 5. Summary of the results of the analysis of variance on SOC and TN concentrations, SOC and TN distribution among fractions (%), and 13C and 15N.
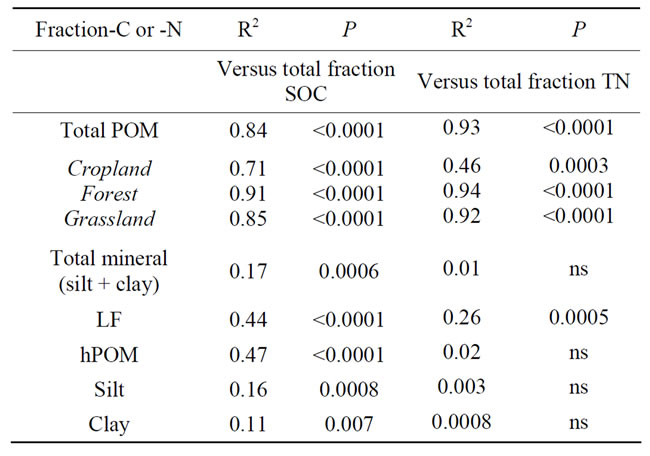
Table 6. Statistical results for linear regressions between total and individual fraction SOC and N in the 0 - 5 cm soil depth.
and N among SOM fractions. This level of detail can inform on how SOM will behave when ecosystems undergo changes in management or land use, ultimately helping decision-making on how to best preserve or increase soil C and N. Furthermore, our investigation serves the broader objectives of the NitroEurope (NEU) project, by providing a dataset that can be integrated with other soil and gas flux measurements, enabling full C and N budget assessments at the different measurement sites. Such assessments will generate extensive empirical field data from widely different ecosystems across Europe which can be used in the validation of biogeochemical process models such as DNDC, RothC or DayCent.
4.1. Total SOC and TN Stocks
In the mineral soil surface (0 - 5 cm), SOC and TN stocks were significantly lower in cropland compared to the other land use systems, as has been found in other studies [e.g.,16,40]. This was a direct result of lower soil C and N concentrations and not just a mere effect of differences in bulk density (Figure 2). The loss of surface SOC and TN in cropped soils has been linked to lower C inputs to the soil, the removal of crop residues, reduced vegetation cover, enhanced mineralization of SOM, erosion and deteriorated soil aggregation. Whereas SOC and TN concentrations were rather uniform across the 0 - 30 cm profile in the croplands, a clear stratification with depth was observed in the non-cropped systems. Similar differences in profile distribution of SOC among different land uses (i.e. more stratified with depth under conservation management than under conventional cropping) were reported by Franzluebbers [41] using an extensive soil survey dataset in Georgia USA, and suggests that SOC stratification should be viewed as an improvement in soil quality.
For assessing whole-ecosystem responses to management, deeper soil depths need to be included to avoid underestimation of total SOC and TN stocks in intensively managed ecosystems [42]. No significant land use effect on SOC and TN stocks could be discerned when considering the entire 0 - 30 cm depth. Against our expectations, we even found higher SOC stocks in a few cropland sites than in some of the forest and grassland soils. This is in contrast to other inventories reporting distinct differences of SOC stocks between cropland and grassland/forest. For example, the European SOC inventories reviewed by Wiesmeier et al. [43] that were restricted to the 0 - 30 cm depth, showed on average 40% and 43% lower SOC stocks for cropland compared with grassland and forest, respectively. The large differences across the sites in our study in terms of soil properties, climate, vegetation and management history could have obscured any effects of land use on total profile SOC and TN stocks. Also, our investigation excluded the litter layer. In forests, the O-horizon can account for a large portion of the total SOC stock. For forests in South-East Germany, Wiesmeier et al. [43] found 2.5 kg m−2 SOC to be stored in the O horizon, or about 35% of what was stored in the A, B and C horizons combined. In addition, C and N contained in litter fragments > 2 mm were not included
 (a)
(a) 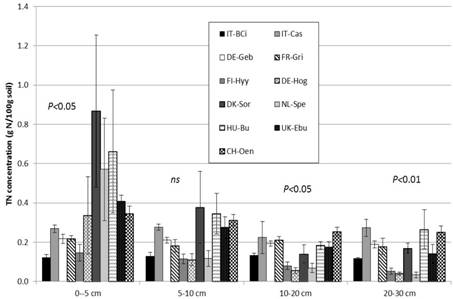 (b)
(b)
Figure 2. Soil organic carbon (a) and total nitrogen (b) concentrations (g C or N/100 g soil) in each sampling depth. Bars and error bars represent means and standard deviations of six replicate samples within a site. Sites code is as in Table 1. Statistical results are presented for land use effects per depth.
in this study, but could contribute to a significant part of the stocks in grassland and forest ecosystems.
Unlike other continental studies [e.g., 2], no clear relationships were found between SOC and TN stocks and any of the climate or soil parameters available for these sites, likely due to the relatively large number of influencing parameters compared to the number of sites included in this study. The highest SOC stocks were found at the Bugac grassland site, which is characterized by a dry-warm continental climate and low-intensity management in terms of grazing and N inputs. These conditions could promote high C inputs while keeping SOM decomposition rates relatively low. The lowest TN stocks were found at the Hyytiälä coniferous forest site, which is characterized by severe N limitation potentially as a result of the low N deposition compared to other forest sites [36,44].
4.2. 13C and 15N Abundance
The 15N pattern in the forest soils with negative 15N in the surface mineral soils and 15N enrichment at deeper
 (a)
(a)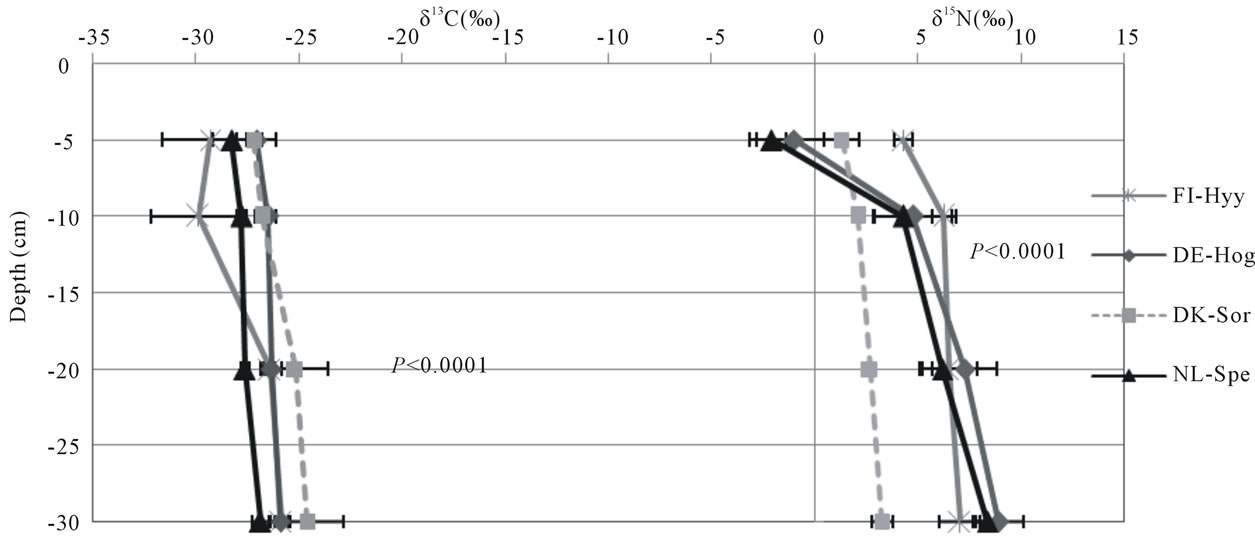 (b)
(b)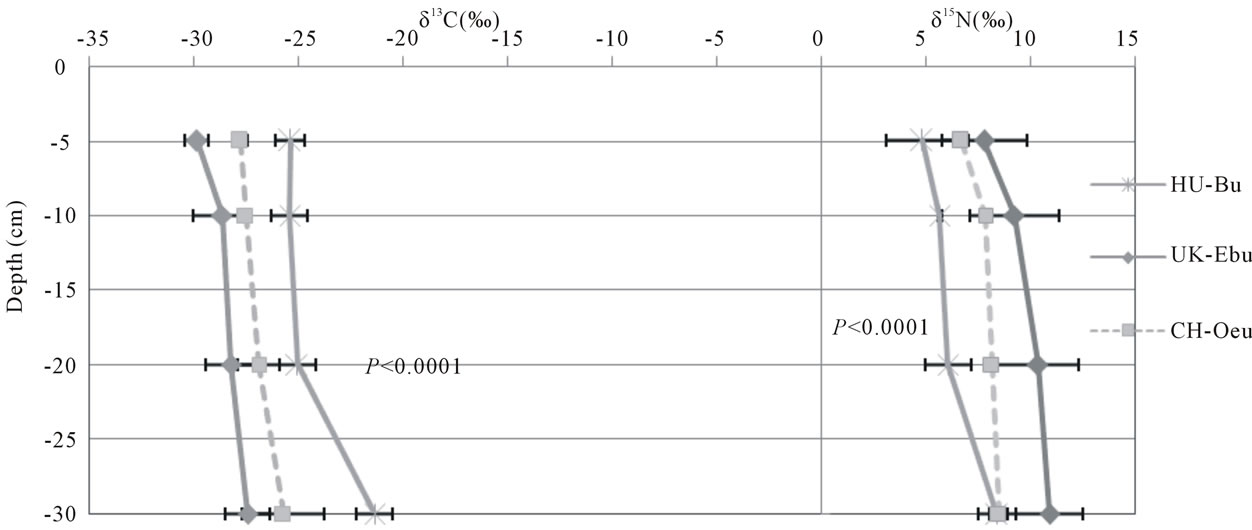 (c)
(c)
Figure 3. Delta 13C (‰) and delta 15N (‰) values across depths for cropland (a), forest (b) and grassland (c) sites. Symbols and error bars represent means and standard deviations of six replicate samples within a site. Sites code is as in Table 1. Statistical results are presented for depth effects per land use.
 (a)
(a)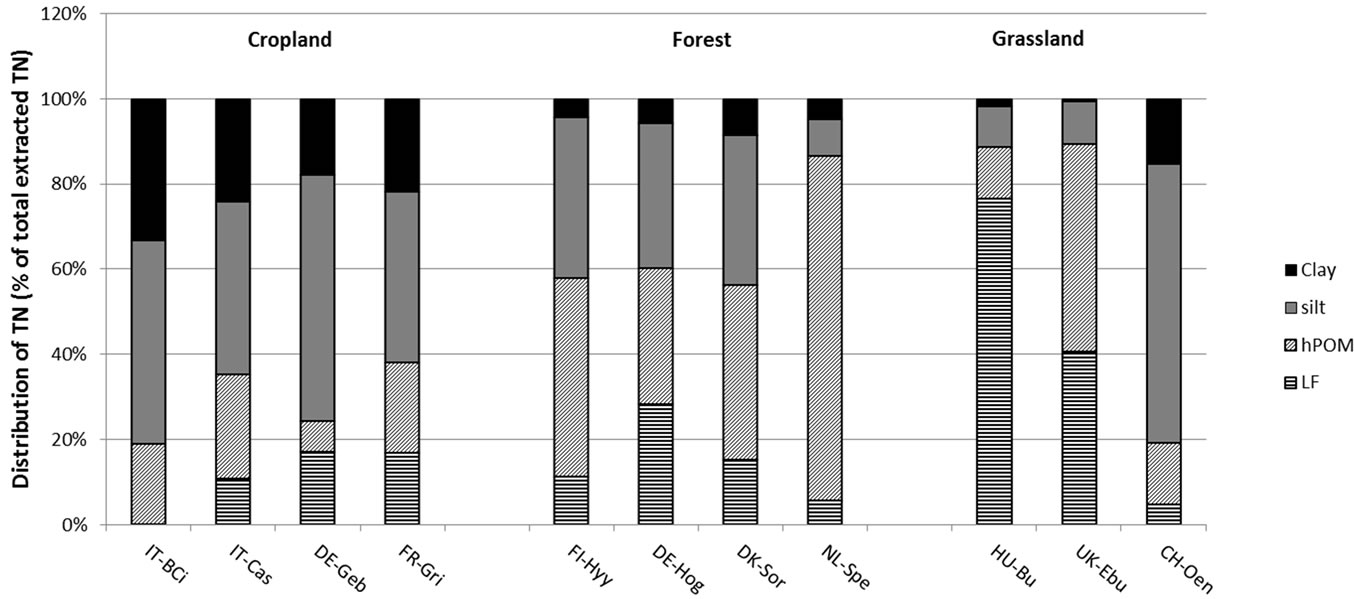 (b)
(b)
Figure 4. Proportion of soil organic carbon (a) and total nitrogen (b) (%) in isolated soil organic matter fractions (light fraction, LF; heavy particulate organic matter, hPOM; silt; clay) in 0 - 5 cm depth. Bars represent means of six replicates within a site. Sites code is as in Table 1.
depths supports our hypothesis of a closed N cycle as has been found in many other temperate forests [31,32]. The higher 15N values in the surface layers of the grassland and cropland soils suggest greater losses of soil N at these sites through processes which discriminate against 15N in soils. In systems that receive N inputs from fertilizer or animal grazing, soil 15N values may also be indicative of the dominant source of soil N. The more enriched 15N values at Easter Bush compared to the other grassland sites could suggest greater N inputs from animal manure [45] due to the more intense grazing at this site [36]. Similarly, the greater 15N values at Borgo Cioffi compare to all other cropland sites could be a result of the use of animal waste as an organic fertilizer for the winter fodder crop at this site [46] owing to the preferential volatilization of 14N ammonia from manure [45].
15N enrichment with soil depth, as found in all forest and grassland sites, is a common observation which has been related to different mechanisms, including 15N isotope discrimination during microbial N transformations, differential preservation of 15N-enriched SOM components during N decomposition, and more recently to the
 (a)
(a)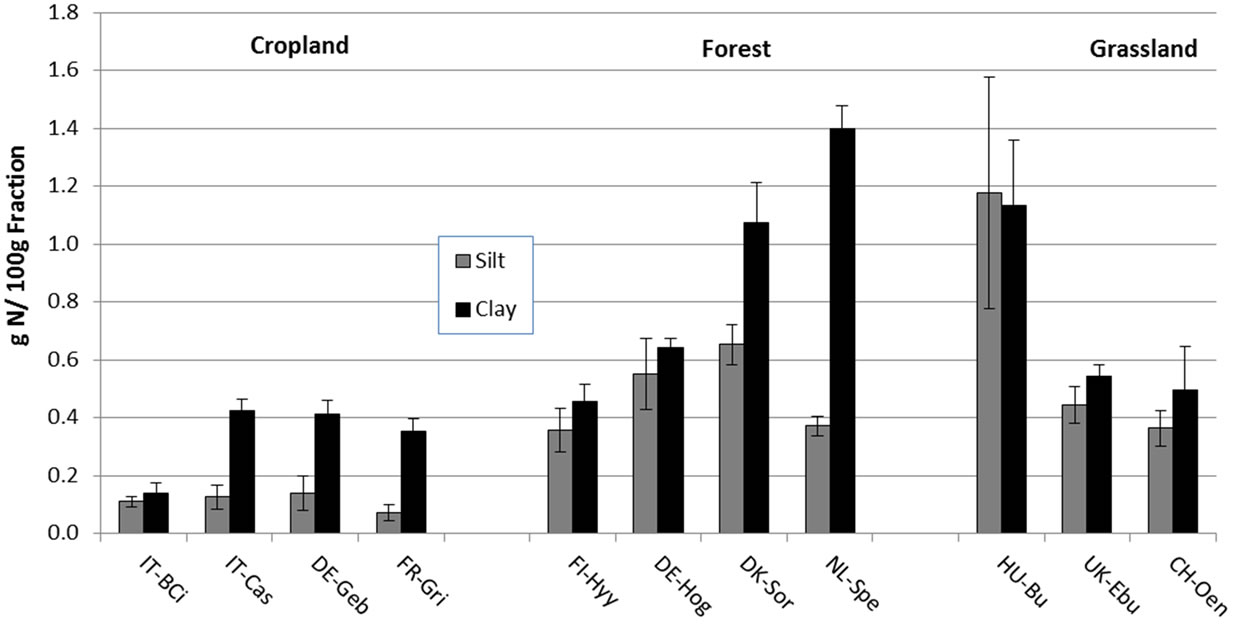 (b)
(b)
Figure 5. Concentration of soil organic carbon (a) and total nitrogen (b) in isolated silt and clay fractions (g C or N/100 g fraction). Bars and error bars represent means and standard deviations of six replicate samples within a site. Sites code is as in Table 1.
buildup of microbial 15N-enriched microbial necromass [47]. The lack of 15N variation within the soil profiles of the cropland soils are probably due to mixing of soil layers by plowing. However, this mixing effect was not apparent for 13C, which showed a slight enrichment with depth in all sites, independent of land use. The observed variation in 13C within a depth layer among the cropland sites is likely a result of the different amount of C4 relative to C3 plant C inputs at these sites. The 13C of the grassland sites, all cultivated with C3 grasses, increased from wetter to drier sites, which could be driven by soil moisture differences and associated differences in plant water use efficiency. Plants grown under water stress become more water-use efficient and exhibit higher 13C values than plants grown under adequate moisture conditions [48]. The highest 13C values were found at Bugac, which is characterized by a warm temperate dry climate (Figure 1), likely causing higher evapotranspiration and water use efficiency (hence lower 13C discrimination in C3 plants) than at the cool temperate moist sites at Easter Bush and Oensingen.
4.3. C and N Distribution among SOM Fractions
The soil fractionation data revealed some interesting and clear trends across the different land-use systems, despite the lack of clear trends in the whole-soil, and is a primary reason to fractionate soils [49]. Undisturbed land uses, such as forest and grassland sites (with the exception of Oensingen) stored the majority of SOM in the POM fractions (LF and hPOM), whereas cropland sites (with the exception of Grignon) stored relatively more SOM in the mineral (silt and clay) fractions. Similar results were reported by Franzluebbers & Stuedemann [50] who found lower POM-C as a proportion of TOC in cropland comred to pasture and forest. These observations suggest that less disturbed ecosystems accumulate relatively more C and N in POM fractions, likely as a result of higher C inputs and slower decomposition rates. The distribution of TOC and TN at the grassland site in Oensingen was more similar to that of the croplands with the majority (80%) contained in the mineral fraction. This may be due to the history of intense cropping at this site, with conversion to grass-clover only in 2001, i.e. six years prior to soil sampling [51].
Another possible cause for the relatively higher proportion of mineral-associated OM at Oensingen and most cropland sites compared to the other sites could be due to differences in soil texture. Much finer soil textures were found in the cropland sites (between 70% and 74% silt and clay) and Oensingen (90% silt and clay) than at the other sites, hence more mineral surfaces are available in these soils for SOM to accumulate. In fact, a negative relationship between soil texture, as represented by wholesoil silt plus clay content, and the percentage of organic C stored in the POM fraction was observed across all study sites (P = 0.007; r2 = 0.58).
Our results are in contrast to other studies, where the mineral-associated OM fraction accounted for most of the total C content independent of land use [e.g., 40]. Relatively high POM proportions have been reported for grassland and forest soils but typically not more than 40% [22,50,52]. This suggests that the European ecosystems included in our study appear to store large amounts of C and N in a rather labile SOM fraction that could potentially be rapidly lost if soils were to be disturbed (e.g., when converted to cropland) [22,50]. The large amounts of POM-C and -N in these systems could also suggest that the mineral (silt and clay) fraction in these systems is C and N saturated [53-55] and cannot store additional C and N inputs. This saturation idea is supported by the higher C and N concentrations in the silt and clay fractions on a per-fraction mass basis in the grassland and forest soils than in the cropland soils (Figure 5), suggesting that the non-cropped systems were closer to or at saturation. To further explore this idea, we calculated the theoretical soil C protective capacity of the different sites based on the linear regressions between soil texture and mineral (silt + clay) C content (g kg−1 soil) developed by Six et al. [54] for different land uses and clay mineralogies. We used the <50 μm regression equations of Six et al. [54] for cultivated, grassland and forest to calculate the silt + clay protective capacity (g C kg soil−1) from <53 μm silt + clay contents obtained in the soil textural analyses performed on our soils (Table 6). These values were then used to derive an estimate of C saturation deficit of the different soils (Table 6) based on Stewart et al. [55] as follows:

with values closer to 1 indicating a lower degree of silt + clay C saturation, and values closer to 0 indicating a higher degree of silt + clay C saturation.
For all forest and grassland sites, with the exception of Easter Bush, the silt + clay C protective capacity estimates were within the 95% confidence intervals of the measured silt + clay C values (Table 7), indicating some degree of silt + clay C saturation in these sites. In contrast, the estimated silt + clay C protective capacity of the cropland soils were greater than the measured silt + clay C values (Table 7). Correspondingly, greater C saturation deficits were calculated for these soils (0.43 - 0.72) compared to the forest soils (−1.26 - 0.07) and grassland soils (0.01 - 0.30, with the exception of Easter Bush: 0.72). These results support the idea that most forest and grassland ecosystems were closer to mineral C saturation, so that any additional C inputs will accumulate in the more labile POM fractions. Hence, any increase in SOC would be captured in differences in the non-mineral-associated OM pools, which was confirmed by the strong correlation between total SOC and total POM (LF + hPOM) (Figure 6), and by the lack of a similar correlation with the mineral-associated SOM fractions (Table 6). A linear relationship between POM-C and TOC was also found by Franzluebbers and Stuedeman [50] for pastures in the Southern Piedmont of the USA, whereas the relationship between non-particulate organic C and TOC flattened off, suggesting mineral C saturation beyond which TOC preferentially accumulates in the POC pool. This would also mean that upon any shift in land-use or any soil disturbance, these ecosystems risk losing significant amounts of SOC from enhanced POM decomposition. If these systems can unlimitedly accumulate C and N in POM pools is not clear and the existence of a saturation level for these pools remains to be tested. Yet, there are some indications that the POM pool can become saturated, in particular through aggregate-protected C saturation [54, 56].
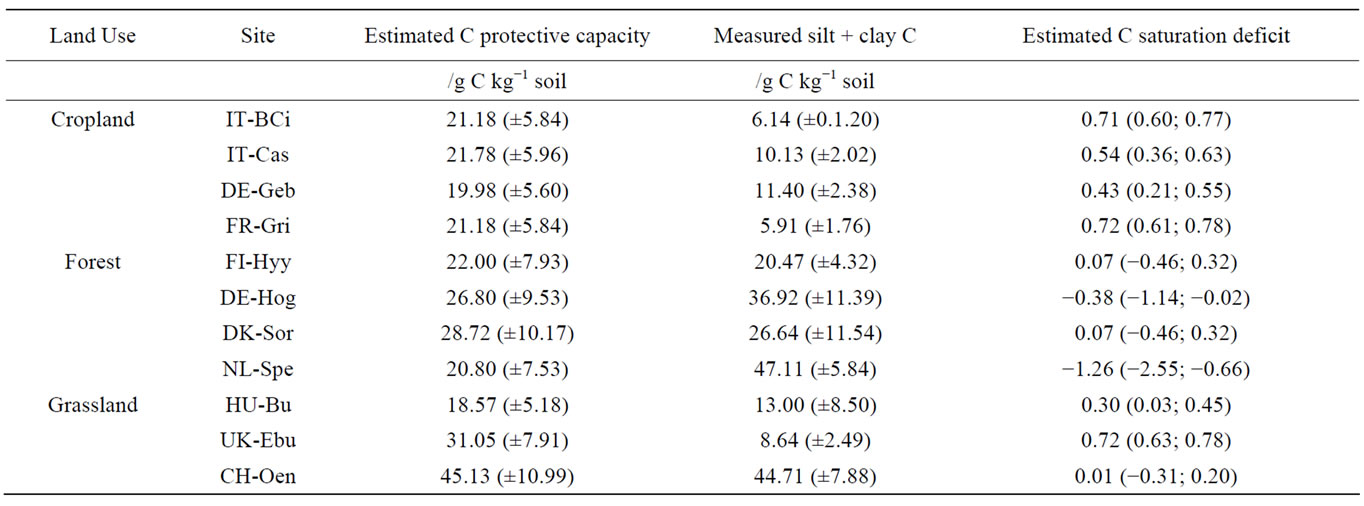
Table 7. Estimates of (1) soil C protective capacity (g C kg−1 soil) using relationships between texture (% silt + clay) and mineral (silt + clay) C content (g C kg−1 soil) developed by Six et al. [54] and (2) C saturation deficit (g C kg−1 soil) based on Stewart et al. [55], for the 0 - 5 depth of the different soils. Values in between brackets represent 95% confidence interval.
 (a)
(a)  (b)
(b)
Figure 6. Linear regression of total particulate organic matter and total fraction organic carbon (a) and total nitrogen (b), for the 0 - 5 cm depth.
The lower concentrations of C and N in the silt and clay fractions (Figure 5) as well as the greater calculated saturation deficit for the croplands (Table 7) indicate that the croplands are further from mineral C saturation compared to the grasslands and forests. These croplands have therefore the potential to sequester additional C and N in mineral fractions if appropriately managed. The continuous physical disturbance in tilled croplands has been shown to stimulate POM decomposition due to enhanced turnover of aggregates, which in turn decreases mineral C and N stabilization compared to minimally disturbed soils [17,19]. We postulate that when these systems are managed in an improved way (less disturbance, e.g. notillage, cover crops, etc.), there could be an actual increase in SOC, due to physical protection of POM through aggregate formation and subsequent mineral C association, that could be stabilized in the long term.
We conclude that the applied fractionation scheme was successful in providing insights into how and to what amount C and N are stored and stabilized in the different soils and land uses included in this study. A simple chemical dispersion followed by a mechanical sieving was sufficient to obtain a highly diagnostic fraction for total soil C and N variability among widely varying ecosystems, and supports the idea that the total POM fraction is a good predictor of total SOM in these investigated ecosystems.
5. Conclusion
This study shows that the investigated Nitro Europe network sites store a large fraction of C and N in POM pools that are particularly vulnerable to soil disturbance such as the one caused by land use change. Best management practices that minimize soil disturbance and increase OM inputs are therefore recommended to avoid rapid losses of SOM in these systems. Rough estimates of saturation deficits suggested most forest and grassland sites to be near mineral C and N saturation, explaining the accumulation of soil C and N in the more labile POM pools, and the strong correlation between POM-C and –N and total SOC and TN. The cropland soils, on the other hand, being further from saturation, appeared to have greater potential to sequester C and N in the mineral fractions, which may be promoted under appropriate management. The observed lower C and N concentration in the silt and clay fractions points to the lower C and N stabilization efficiency at these cropland sites, likely due to the continuous physical disturbance and enhanced aggregate turnover.
6. Acknowledgements
Publication of this article was sponsored by the Colorado State University Libraries Open Access Research and Scholarship Fund. We thank Dr. Catherine Stewart for her insightful comments on this work. The study was funded by the European Union, Nitro Europe IP #017841. We are grateful to all the Nitro Europe level 3 site managers for their assistance during sampling.
REFERENCES
- N. H. Batjes, “Total Carbon and Nitrogen in the Soils of the World,” European Journal of Soil Science, Vol. 47, No. 2, 1996, pp. 151-163. http://dx.doi.org/10.1111/j.1365-2389.1996.tb01386.x
- R. F. Follett, C. E. Stewart, E. G. Pruessner and J. M. Kim- ble, “Effects of Climate Change on Soil Carbon and Nitrogen Storage in the US Great Plains,” Journal of Soil and Water Conservation, Vol. 67, No. 5, 2012, pp. 331- 342. http://dx.doi.org/10.2489/jswc.67.5.331
- E. A. Davidson, S. E. Trumbore and R. Amundson, “Biogeochemistry—Soil Warming and Organic Carbon Content,” Nature, Vol. 408, No. 6814, 2000, pp. 789-790. http://dx.doi.org/10.1038/35048672
- R. T. Conant, S. M. Ogle, E. A. Paul and K. Paustian, “Measuring and Monitoring Soil Organic Carbon Stocks in Agricultural Lands for Climate Mitigation,” Frontiers in Ecology and the Environment, Vol. 9, No. 3, 2011, pp. 169-173. http://dx.doi.org/10.1890/090153
- S. Sleutel, S. De Neve, G. Hofman, P. Boeckx, D. Beheydt, O. Van Cleemput, I. Mestdagh, P. Lootens, L. Carlier, N. Van Camp, H. Verbeeck, I. Vande Walle, R. Samson, N. Lust and R. Lemeur, “Carbon Stock Changes and Carbon Sequestration Potential of Flemish Cropland Soils,” Global Change Biology, Vol. 9, No. 8, 2003, pp. 1193- 1203. http://dx.doi.org/10.1046/j.1365-2486.2003.00651.x
- S. Spencer, S. M. Ogle, F. J. Breidt, J. Goebel and K. Pau- stian, “Designing a National Soil Carbon Monitoring Network to Support Climate Change Policy: A Case Example for US Agricultural Lands,” Greenhouse Gas Management & Measurement, Vol. 1, No. 3-4, 2011, pp. 167-178. http://dx.doi.org/10.1080/20430779.2011.637696
- W. J. Parton, D. S. Ojima, C. V. Cole and D. S. Schimel, “A General Model for Soil Organic Matter Dynamics: Sensitivity to Litter Chemistry, Texture and Management,” Soil Science Society of America Journal, Vol. 39, 1994, pp. 147-167.
- W. J. Parton, M. Hartman, D. Ojima and D. Schimel, “Daycent and Its Land Surface Submodel: Description and Testing,” Global and Planetary Change, Vol. 19, No. 1-4, 1998, pp. 35-48. http://dx.doi.org/10.1016/S0921-8181(98)00040-X
- K. Coleman, D. S. Jenkinson, G. J. Crocker, P. R. Grace, J. Klir, M. Korschens, P. R. Poulton and D. D. Richter, “Simulating Trends in Soil Organic Carbon in Long-Term Experiments Using RothC-26.3,” Geoderma, Vol. 81, No. 1-2, 1997, pp. 29-44. http://dx.doi.org/10.1016/S0016-7061(97)00079-7
- C. S. Li, “Modeling Trace Gas Emissions from Agricultural Ecosystems,” Nutrient Cycling in Agroecosystems, Vol. 58, No. 1-3, 2000, pp. 259-276. http://dx.doi.org/10.1023/A:1009859006242
- S. M. Ogle, F. J. Breidt, M. Easter, S. Williams, K. Killian and K. Paustian, “Scale and Uncertainty in Modeled Soil Organic Carbon Stock Changes for US Croplands Using a Process-Based Model,” Global Change Biology, Vol. 16, No. 2, 2010, pp. 810-822. http://dx.doi.org/10.1111/j.1365-2486.2009.01951.x
- E. D. Schulze, S. Luyssaert, P. Ciais, A. Freibauer, I. A. Janssens, J. F. Soussana, P. Smith, J. Grace, I. Levin, B. Thiruchittampalam, M. Heimann, A. J. Dolman, R. Valentini, P. Bousquet, P. Peylin, W. Peters, C. Rodenbeck, G. Etiope, N. Vuichard, M. Wattenbach, G. J. Nabuurs, Z. Poussi, J. Nieschulze, J. H. Gash and T. Carbo Europe, “Importance of Methane and Nitrous Oxide for Europe’s Terrestrial Greenhouse-Gas Balance,” Nature Geoscience, Vol. 2, No. 12, 2009, pp. 842-850. http://dx.doi.org/10.1038/ngeo686
- M. W. I. Schmidt, M. S. Torn, S. Abiven, T. Dittmar, G. Guggenberger, I. A. Janssens, M. Kleber, I. Kogel-Knabner, J. Lehmann, D. A. C. Manning, P. Nannipieri, D. P. Rasse, S. Weiner and S. E. Trumbore, “Persistence of Soil Organic Matter as an Ecosystem Property,” Nature, Vol. 478, No. 7367, 2011, pp. 49-56. http://dx.doi.org/10.1038/nature10386
- J. Six and J. D. Jastrow, “Organic Matter Turnover,” In: R. Lal, Ed., Encyclopedia of Soil Science, Marcel Dekker, New York, 2002, pp. 936-942.
- P. Smith, “How Long before a Change in Soil Organic Carbon Can Be Detected?” Global Change Biology, Vol. 10, No. 11, 2004, pp. 1878-1883. http://dx.doi.org/10.1111/j.1365-2486.2004.00854.x
- I. Del Galdo, J. Six, A. Peressotti and M. F. Cotrufo, “Assessing the Impact of Land-Use Change on Soil C Sequestration in Agricultural Soils by Means of Organic Matter Fractionation and Stable C Isotopes,” Global Change Biology, Vol. 9, No. 8, 2003, pp. 1204-1213. http://dx.doi.org/10.1046/j.1365-2486.2003.00657.x
- K. Denef, J. Six, R. Merckx and K. Paustian, “Carbon Sequestration in Microaggregates of No-Tillage Soils with Different Clay Mineralogy,” Soil Science Society of America Journal, Vol. 68, No. 6, 2004, pp. 1935-1944. http://dx.doi.org/10.2136/sssaj2004.1935
- J. Leifeld and I. Kogel-Knabner, “Soil Organic Matter Fractions as Early Indicators for Carbon Stock Changes under Different Land-Use?” Geoderma, Vol. 124, No. 1-2, 2005, pp. 143-155. http://dx.doi.org/10.1016/j.geoderma.2004.04.009
- J. Six, E. T. Elliott and K. Paustian, “Soil Macroaggregate Turnover and Microaggregate Formation: A Mechanism for C Sequestration under No-Tillage Agriculture,” Soil Biology & Biochemistry, Vol. 32, No. 14, 2000, pp. 2099- 2103. http://dx.doi.org/10.1016/S0038-0717(00)00179-6
- E. G. Gregorich and M. H. Beare, “Physically Uncomplexed Organic Matter,” In: M. R. Carter and E. G. Gregorich, Eds., Soil Sampling and Methods of Analysis, CRC Press/Taylor & Francis, Boca Raton, 2008, pp. 607- 616.
- C. E. Stewart, R. F. Follett, J. Wallace and E. G. Pruessner, “Impact of Biosolids and Tillage on Soil Organic Matter Fractions: Implications of Carbon Saturation for Conservation Management in the Virginia Coastal Plain,” Soil Science Society of America Journal, Vol. 76, No. 4, 2012, pp. 1257-1267. http://dx.doi.org/10.2136/sssaj2011.0165
- C. A. Cambardella and E. T. Elliott, “Particulate Soil Organic-Matter Changes across a Grassland Cultivation Sequence,” Soil Science Society of America Journal, Vol. 56, No. 3, 1992, pp. 777-783. http://dx.doi.org/10.2136/sssaj1992.03615995005600030017x
- A. S. Grandy and J. C. Neff, “Molecular C Dynamics Downstream: The Biochemical Decomposition Sequence and Its Impact on Soil Organic Matter Structure and Function,” Science of the Total Environment, Vol. 404, No. 2-3, 2008, pp. 297-307. http://dx.doi.org/10.1016/j.scitotenv.2007.11.013
- M. Helfrich, B. Ludwig, P. Buurman and H. Flessa, “Effect of Land Use on the Composition of Soil Organic Matter in Density and Aggregate Fractions as Revealed by Solid-State C-13 NMR Spectroscopy,” Geoderma, Vol. 136, No. 1-2, 2006, pp. 331-341. http://dx.doi.org/10.1016/j.geoderma.2006.03.048
- P. Sollins, C. Swanston, M. Kleber, T. Filley, M. Kramer, S. Crow, B. A. Caldwell, K. Lajtha and R. Bowden, “Organic C and N Stabilization in a Forest Soil: Evidence from Sequential Density Fractionation,” Soil Biology & Biochemistry, Vol. 38, No. 11, 2006, pp. 3313-3324. http://dx.doi.org/10.1016/j.soilbio.2006.04.014
- J. S. Clemente, A. J. Simpson and M. J. Simpson, “Association of Specific Organic Matter Compounds in Size Fractions of Soils under Different Environmental Controls,” Organic Geochemistry, Vol. 42, No. 10, 2011, pp. 1169-1180. http://dx.doi.org/10.1016/j.orggeochem.2011.08.010
- K. F. Nadelhoffer and B. Fry, “Controls on Natural N-15 and C-13 Abundances in Forest Soil Organic Matter,” Soil Science Society of America Journal, Vol. 52, No. 6, 1988, pp. 1633-1640. http://dx.doi.org/10.2136/sssaj1988.03615995005200060024x
- J. Balesdent, A. Mariotti and B. Guillet, “Natural 13C Abundance as a Tracer for Studies of Soil Organic Matter Dynamics,” Soil Biology & Biochemistry, Vol. 19, No. 1, 1987, pp. 25-30. http://dx.doi.org/10.1016/0038-0717(87)90120-9
- A. Diochon and L. Kellman, “Natural Abundance Measurements of (13)C Indicate Increased Deep Soil Carbon Mineralization after Forest Disturbance,” Geophysical Research Letters, Vol. 35, No. 14, 2008. http://dx.doi.org/10.1029/2008GL034795
- J. R. Ehleringer, N. Buchmann and L. B. Flanagan, “Carbon Isotope Ratios in Belowground Carbon Cycle Processes,” Ecological Applications, Vol. 10, No. 2, 2000, pp. 412-422. http://dx.doi.org/10.1890/1051-0761(2000)010[0412:CIRIBC]2.0.CO;2
- P. Hogberg, L. Hogbom, H. Schinkel, M. Hogberg, C. Johannisson and H. Wallmark, “N-15 Abundance of Surface Soils, Roots and Mycorrhizas in Profiles of European Forest Soils,” Oecologia, Vol. 108, No. 2, 1996, pp. 207- 214.
- K. J. Nadelhoffer and B. Fry, “Nitrogen Isotope Studies in Forest Ecosystems,” In: K. Lajtha and R. Michener, Eds., Stable Isotopes in Ecology and Environmental Science, Blackwell Scientific Publications, Boston, 1994, pp. 23-44.
- P. Hogberg, “Tansley Review No 95 - N-15 Natural Abun- dance in Soil-Plant Systems,” New Phytologist, Vol. 137, No. 2, 1997, pp. 179-203. http://dx.doi.org/10.1046/j.1469-8137.1997.00808.x
- M. A. Sutton, E. Nemitz, J. W. Erisman, C. Beier, K. B. Bahl, P. Cellier, W. de Vries, F. Cotrufo, U. Skiba, C. Di Marco, S. Jones, P. Laville, J.F. Soussana, B. Loubet, M. Twigg, D. Famulari, J. Whitehead, M.W. Gallagher, A. Neftel, C. R. Flechard, B. Herrmann, P. L. Calanca, J. K. Schjoerring, U. Daemmgen, L. Horvath, Y. S. Tang, B. A. Emmett, A. Tietema, J. Penuelas, M. Kesik, N. Brueggemann, K. Pilegaard, T. Vesala, C. L. Campbell, J. E. Olesen, U. Dragosits, M. R. Theobald, P. Levy, D. C. Mobbs, R. Milne, N. Viovy, N. Vuichard, J. U. Smith, P. Smith, P. Bergamaschi, D. Fowler and S. Reis, “Challenges in Quantifying Biosphere-Atmosphere Exchange of Nitrogen Species,” Environmental Pollution, Vol. 150, No. 1, 2007, pp. 125-139. http://dx.doi.org/10.1016/j.envpol.2007.04.014
- U. Skiba, J. Drewer, Y. S. Tang, N. van Dijk, C. Helfter, E. Nemitz, D. Famulari, J. N. Cape, S. K. Jones, M. Twigg, M. Pihlatie, T. Vesala, K. S. Larsen, M. S. Carter, P. Ambus, A. Ibrom, C. Beier, A. Hensen, A. Frumau, J. W. Erisman, N. Bruggemann, R. Gasche, K. ButterbachBahl, A. Neftel, C. Spirig, L. Horvath, A. Freibauer, P. Cellier, P. Laville, B. Loubet, E. Magliulo, T. Bertolini, G. Seufert, M. Andersson, G. Manca, T. Laurila, M. Aurela, A. Lohila, S. Zechmeister-Boltenstern, B. Kitzler, G. Schaufler, J. Siemens, R. Kindler, C. Flechard and M. A. Sutton, “Biosphere-Atmosphere Exchange of Reactive Nitrogen and Greenhouse Gases at the NitroEurope Core Flux Measurement Sites: Measurement Strategy and First Data Sets,” Agriculture Ecosystems & Environment, Vol. 133, No. 3-4, 2009, pp. 139-149. http://dx.doi.org/10.1016/j.agee.2009.05.018
- G. Schaufler, B. Kitzler, A. Schindlbacher, U. Skiba, M. A. Sutton and S. Zechmeister-Boltenstern, “Greenhouse Gas Emissions from European Soils under Different Land Use: Effects of Soil Moisture and Temperature,” European Journal of Soil Science, Vol. 61, No. 5, 2010, pp. 683-696. http://dx.doi.org/10.1111/j.1365-2389.2010.01277.x
- K. Pilegaard, T. N. Mikkelsen, C. Beier, N. O. Jensen, P. Ambus and H. Ro-Poulsen, “Field Measurements of Atmosphere-Biosphere Interactions in a Danish Beech Forest,” Boreal Environment Research, Vol. 8, No. 4, 2003, pp. 315-333.
- D. Harris, W. R. Horwath and C. van Kessel, “Acid Fumigation of Soils to Remove Carbonates Prior to Total Organic Carbon or Carbon-13 Isotopic Analysis,” Soil Science Society of America Journal, Vol. 65, No. 6, 2001, pp. 1853-1856. http://dx.doi.org/10.2136/sssaj2001.1853
- F. Marzaioli, C. Lubritto, I. Del Galdo, A. D’Onofrio, M. F. Cotrufo and F. Terrasi, “Comparison of Different Soil Organic Matter Fractionation Methodologies: Evidences from Ultrasensitive (14)C Measurements,” Nuclear Instruments & Methods in Physics Research Section BBeam Interactions with Materials and Atoms, Vol. 268, No. 7-8, 2010, pp. 1062-1066. http://dx.doi.org/10.1016/j.nimb.2009.10.098
- S. De Gryze, J. Six, K. Paustian, S. J. Morris, E. A. Paul and R. Merckx, “Soil Organic Carbon Pool Changes Following Land-Use Conversions,” Global Change Biology, Vol. 10, No. 7, 2004, pp. 1120-1132. http://dx.doi.org/10.1111/j.1529-8817.2003.00786.x
- A. J. Franzluebbers. “Depth Distribution of Soil Organic Carbon as a Signature of Soil Quality,” Proceedings of the 19th World Congress of Soil Science: Soil Solutions for a Changing World. Symposium 4.1.1 Valuing the Soil’s Natural Capital, Brisbane, 2010, pp. 1-4.
- R. B. Harrison, P. W. Footen and B. D. Strahm, “Deep Soil Horizons: Contribution and Importance to Soil Carbon Pools and in Assessing Whole-Ecosystem Response to Management and Global Change,” Forest Science, Vol. 57, No. 1, 2011, pp. 67-76.
- M. Wiesmeier, P. Sporlein, U. Geuss, E. Hangen, S. Haug, A. Reischl, B. Schilling, M. von Lutzow and I. KogelKnabner, “Soil Organic Carbon Stocks in Southeast Germany (Bavaria) as Affected by Land Use, Soil Type and Sampling Depth,” Global Change Biology, Vol. 18, No. 7, 2012, pp. 2233-2245. http://dx.doi.org/10.1111/j.1365-2486.2012.02699.x
- K. Pilegaard, U. Skiba, P. Ambus, C. Beier, N. Bruggemann, K. Butterbach-Bahl, J. Dick, J. Dorsey, J. Duyzer, M. Gallagher, R. Gasche, L. Horvath, B. Kitzler, A. Leip, M. K. Pihlatie, P. Rosenkranz, G. Seufert, T. Vesala, H. Westrate and S. Zechmeister-Boltenstern, “Factors Controlling Regional Differences in Forest Soil Emission of Nitrogen Oxides (NO and N2O),” Biogeosciences, Vol. 3, No. 4, 2006, pp. 651-661. http://dx.doi.org/10.5194/bg-3-651-2006
- A. S. Bateman, S. D. Kelly and T. D. Jickells, “Nitrogen Isotope Relationships between Crops and Fertilizer: Implications for Using Nitrogen Isotope Analysis as an Indicator of Agricultural Regime,” Journal of Agricultural and Food Chemistry, Vol. 53, No. 14, 2005, pp. 5760- 5765. http://dx.doi.org/10.1021/jf050374h
- S. Ranucci, T. Bertolini, L. Vitale, P. Di Tommasi, L. Ottaiano, M. Oliva, U. Amato, A. Fierro and V. Magliulo, “The Influence of Management and Environmental Variables on Soil N2O Emissions in a Crop System in Southern Italy,” Plant and Soil, Vol. 343, No. 1-2, 2011, pp. 83-96. http://dx.doi.org/10.1007/s11104-010-0674-x
- D. Huygens, K. Denef, R. Vandeweyer, R. Godoy, O. Van Cleemput and P. Boeckx, “Do Nitrogen Isotope Patterns Reflect Microbial Colonization of Soil Organic Matter Fractions?” Biology and Fertility of Soils, Vol. 44, No. 7, 2008, pp. 955-964. http://dx.doi.org/10.1007/s00374-008-0280-8
- K. T. Hubick, G. D. Farquhar and R. Shorter, “Correlation between Water Use Efficiency and Carbon Isotope Discrimination in Diverse Peanut (Arachis) Germplasm,” Australian Journal of Plant Physiology, Vol. 13, No. 6, 1986, pp. 803-816. http://dx.doi.org/10.1071/PP9860803
- K. Denef, A. F. Plante and J. Six, “Characterization of Soil Organic Matter,” In: W. L. Kutsch, M. Bahn and A. Heinemeyer, Eds., Soil Carbon Dynamics: An Integrated Methodology, Cambridge University Press, Cambridge, 2009, pp. 91-126.
- A. J. Franzluebbers and J. A. Stuedemann, “Particulate and Non-Particulate Fractions of Soil Organic Carbon under Pastures in the Southern Piedmont USA,” Environmental Pollution, Vol. 116, No. S1, 2002, pp. S53-S62. http://dx.doi.org/10.1016/S0269-7491(01)00247-0
- C. Ammann, C. Spirig, J. Leifeld and A. Neftel, “Assessment of the Nitrogen and Carbon Budget of Two Managed Temperate Grassland Fields,” Agriculture Ecosystems & Environment, Vol. 133, No. 3-4, 2009, pp. 150- 162. http://dx.doi.org/10.1016/j.agee.2009.05.006
- M. R. Carter and E. G. Gregorich, “Carbon and Nitrogen Storage by Deep-Rooted Tall Fescue (Lolium Arundinaceum) in the Surface and Subsurface Soil of a Fine Sandy Loam in Eastern Canada,” Agriculture Ecosystems & Environment, Vol. 136, No. 1-2, 2010, pp. 125-132. http://dx.doi.org/10.1016/j.agee.2009.12.005
- J. Hassink, “The Capacity of Soils to Preserve Organic C and N by Their Association with Clay and Silt Particles,” Plant and Soil, Vol. 191, No. 1, 1997, pp. 77-87. http://dx.doi.org/10.1023/A:1004213929699
- J. Six, R. T. Conant, E. A. Paul and K. Paustian, “Stabilization Mechanisms of Soil Organic Matter: Implications for C-Saturation of Soils,” Plant and Soil, Vol. 241, No. 2, 2002, pp. 155-176. http://dx.doi.org/10.1023/A:1016125726789
- C. E. Stewart, K. Paustian, R. T. Conant, A. F. Plante and J. Six, “Soil Carbon Saturation: Evaluation and Corroboration by Long-Term Incubations,” Soil Biology & Biochemistry, Vol. 40, No. 7, 2008, pp. 1741-1750. http://dx.doi.org/10.1016/j.soilbio.2008.02.014
- H. Chung, K. J. Ngo, A. F. Plante and J. Six, “Evidence for Carbon Saturation in a Highly Structured and Organic-Matter-Rich Soil,” Soil Science Society of America Journal, Vol. 74, No. 1, 2010, pp. 130-138. http://dx.doi.org/10.2136/sssaj2009.0097
NOTES
*Corresponding author.