Open Journal of Modern Hydrology Vol.05 No.02(2015), Article ID:55470,12
pages
10.4236/ojmh.2015.52003
Hydrological Controls on Nutrient Exportation from Old-Growth Evergreen Rainforests and Eucalyptus nitens Plantation in Headwater Catchments at Southern Chile
Carlos E. Oyarzun1, Pedro Hervé-Fernández2,3, Dries Huygens3,4,5, Pascal Boeckx3, Niko E. C. Verhoest2
1Instituto de Ciencias Ambientales y Evolutivas, Facultad de Ciencias, Universidad Austral de Chile, Valdivia, Chile
2Laboratory of Hydrology and Water Management, Faculty Bioscience Engineering, Ghent University, Ghent, Belgium
3Isotope Bioscience Laboratory (ISOFYS), Ghent University, Ghent, Belgium
4Institute of Agricultural Engineering and Soil Science, Faculty of Agricultural Sciences, Universidad Austral de Chile, Valdivia, Chile
5Instituto Multidisciplinario de Biología Vegetal (IMBIV), Consejo Nacional de Investigaciones Científicas y Técnicas de Argentina, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Córdoba, Argentina
Email: coyarzun@uach.cl
Copyright © 2015 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 1 December 2014; accepted 7 April 2015; published 9 April 2015
ABSTRACT
Soil cover disturbances have a direct effect on biogeochemistry, potentially enhancing
nutrient loss, land degradation and associated changes in ecosystem services and
livelihood support. The objective of this study was to assess how canopy affected
throughfall chemistry and how hydrology affected stream nutrient load responses
in two watersheds dominated by native old-growth evergreen rainforest (NF) and exotic
plantation of Eucalyptus nitens (EP), located at the Coastal mountain range of southern
Chile (40˚S). We measured nitrogen (NO3-N, NH4-N, Organic-N,
Total-N) and total phosphorus (Total-P) at catchment discharge, and δ18O
in throughfall precipitation and stream discharge in both catchments, in order to
separate throughfall (or new water) contributions during storm events. It was hypothesized
that all nutrients showed an increase in concentration as discharge increased (or
enhanced hydrological access), in EP; but not in NF. Our results indicated that
Organic-N, Total-N and Total-P concentrations were positively related to discharge.
However,
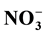 -N showed a negative correlation
with catchment discharge. Organic-N and Total-P showed a flush during storm events;
the opposite was observed for
-N showed a negative correlation
with catchment discharge. Organic-N and Total-P showed a flush during storm events;
the opposite was observed for
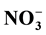 -N. However, this behavior
suggested that
-N. However, this behavior
suggested that
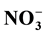 -N was being retained by
charged particles or soil micro biota, whether Organic-N was flushed as it was more
concentrated in big pore water that was not tightly attached, compared with
-N was being retained by
charged particles or soil micro biota, whether Organic-N was flushed as it was more
concentrated in big pore water that was not tightly attached, compared with
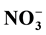 -N.
-N.
Keywords:
Native Rainforests, Exotic Plantations, Nutrient Fluxes, Hydrological Controls, Headwater Catchments

1. Introduction
Human disturbances have a great impact on native forest communities. This may lead to land degradation, causing changes in ecosystem services and livelihood support [1] . Stream nutrient loads are very sensitive to vegetation changes due human disturbances [2] -[4] , but also to variables related to ecosystem hydrology, including infiltration rates, rainfall, and surface runoff [5] . Human-induced alteration of forest canopies and their soils have a significant impact on the hydrological controls of the nutrients (nitrogen, phosphorous and base cations) that reach the stream water. In this sense, export during storm events, concentration of nutrients could exhibit one of three general trends with respect to stream discharge: 1) dilution, 2) hydrological constant, or 3) enhanced hydrological access [6] .
- Dilution occurs whenever stream discharge increases, but not the chemical concentration. This type of relationship is expected for chemicals with strong internal watershed source that does not increase in magnitude as a function of increased hydrologic throughput. We expect these elements to be diluted by enhanced through- put of water.
- Hydrological constant controls are characterized by a balance between water discharge and chemical concentration. This relationship is expected for elements that are delivered with precipitation water to the watershed, but that lack significant internal production or consumption processes. In the strictest sense, such idealized hydrological constancy may be rare since evapotranspiration provides a mechanism for concentrating solutes in deeper soil waters, and thus may impart differences in the delivery of chemicals and water depending on soil water flow paths. Some form of constancy may also be expected for elements that are chemically buffered within soils (e.g. via cation exchange reactions), but only if rates of hydrologic throughput remain low enough to maintain some form of equilibrium between soils and soil solutions. This is not likely to occur in most natural soils that experience variable hydrologic inputs over time.
- Enhanced hydrological access refers to controls that exhibit increasing chemical concentration with increasing discharge. The most common enhanced hydrological access is for chemicals found in areas of a watershed that are only active during periods of high flows. For example, in the case of elements produced in the surface soil horizons, as the region of subsurface flow deepens, i.e. as the saturated soil boundary approaches the soil surface, the flowing water increasingly accesses these elements. In recent years, this hydrological process has often been referred to as “piston flow”.
Native temperate rainforests of southern Chile covering an area of 13.5 million
ha, represent an important global reserve of temperate forests with an extraordinary
genetic, phytogeographic and ecological significance [7] . Native forests in the
Valdivian eco-region (36˚S through 48˚S) have suffered anthropic disturbances due
to fires, logging practices, or its conversion to agricultural land and exotic fast-growing
plantations. Temperate rain forest ecosystems of southern Chile have efficient mechanisms
of retention for essential nutrients, especially
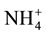 and
and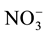 . [8] -[10]
described that the dominant form of N leaching was dissolved organic nitrogen (DON)
in unpolluted forests of southern Chile. [11] described that DIN inputs did not
end up in the soil water compartment, and gave evidence that Organic-N losses, originate
from bio-unavailable compounds leaching from slow-turnover soil organic matter pools.
While [3] reported that conversion from native forests to exotic fast- growing plantations
was likely to decrease catchment N retention.
. [8] -[10]
described that the dominant form of N leaching was dissolved organic nitrogen (DON)
in unpolluted forests of southern Chile. [11] described that DIN inputs did not
end up in the soil water compartment, and gave evidence that Organic-N losses, originate
from bio-unavailable compounds leaching from slow-turnover soil organic matter pools.
While [3] reported that conversion from native forests to exotic fast- growing plantations
was likely to decrease catchment N retention.
Besides the effects over N retention, [3] [5] described that soil water infiltration rates under eucalypts plantations were lower (6.7 ± 5.0 and 23.0 ± 19.7 mm∙hr−1) and higher under a second growth native evergreen forest (76.9 ± 56.7 and 703 ± 380 mm∙hr−1). In the former case, these would be large amounts of rainfall reaching the catchment after storm events, therefore affect N and P dynamics.
The temperate climate region in southern Chile still reflects undisturbed environmental conditions, with total nitrogen (Total-N) bulk precipitation inputs of less than 3 kg∙ha−1∙yr−1 at the Coastal mountain range [3] . This is in strong contrast with land cover, which has been altered significantly over the last decades and centuries. Only fragments of the original forest vegetation remain unaltered, and are located in the coastal and Andes mountain range. Agricultural areas dominate the central valley of southern Chile; however, exotic tree plantations are spreading fast over the coastal mountain range [3] [12] [13] . These observations make this region ideal to study land cover change effects on biogeochemical nutrient cycling, without biases due to increased atmospheric nutrient depositions.
The objectives of this study are: 1) to assess how canopy affected throughfall chemistry,
and 2) to compare how hydrological variability of “new” or event water, and “old”
or groundwater contributions during storm events affects nutrient loads under different
land covers of native evergreen rainforest (NF) and exotic plantation of Eucalyptus
nitens (EP). To find these differences we measured nutrients such as: nitrogen (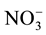 -N,
-N,
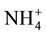 -N, Organic-N
and Total-N) and total phosphorus (Total-P) in bulk and throughfall precipitation
and catchment discharge, and δ18O in throughfall and storm discharge
in small headwater catchments located at coastal mountain range in southern Chile
(40˚S). Our hypothesis is that new water will have higher contribution in EP, but
not in NF, therefore controlling nutrient exportation.
-N, Organic-N
and Total-N) and total phosphorus (Total-P) in bulk and throughfall precipitation
and catchment discharge, and δ18O in throughfall and storm discharge
in small headwater catchments located at coastal mountain range in southern Chile
(40˚S). Our hypothesis is that new water will have higher contribution in EP, but
not in NF, therefore controlling nutrient exportation.
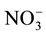 -N and
-N and
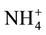 -N will show
a dilution in old-growth native evergreen (NF), but not in Eucalyptus nitens (EP);
while Organic-N and Total-P will show an enhanced hydrological access in EP, but
not in NF, due to the different water infiltration rates are higher in native forests
and lower in Eucalyptus plantation [12] .
-N will show
a dilution in old-growth native evergreen (NF), but not in Eucalyptus nitens (EP);
while Organic-N and Total-P will show an enhanced hydrological access in EP, but
not in NF, due to the different water infiltration rates are higher in native forests
and lower in Eucalyptus plantation [12] .
2. Study Sites
We selected two catchments with different land cover at the Coastal mountain range (40˚S), near the city of Valdivia, Chile. NF catchment, covered by old-growth native evergreen rainforest; and EP catchment covered with exotic fast growing Eucalyptus nitens (Figure 1). The drainage area of NF is 12.5 ha with an average alti- tude of 336 m above sea level (a.s.l.) and average slope of 15.4%. The main canopy species in this catchment are Eucryphia cordifolia, Aextoxicon punctatum and Laureliopsis philippiana. This last shows the highest density (718 tree ha−1) and basal area (37.2 m2∙ha−1). The understorey is dominated by Amormyrtus luma, Amomyrtus

Figure 1. Old growth native forest (NF) and Eucalyptus nitens plantation (EP) research sites near the city of Valdivia, south central Chile (40˚S).
meli, Drimys winteri and Myrceugenia planipes. On the other hand, EP catchment has an area of 54.8 ha, an average altitude of 35 m a.s.l. and average slope of 34%. This catchment is actually covered with 6 and 16 years old E. nitens plantations, established on soils that already had 5 E. nitens rotations. Tree density equals 2911 tree ha−1 and the basal area is 132 m2∙ha−1. The riparian vegetation is dominated by native species such as Aristotelia chilensis, Luma apiculata, Fuchsia magellanica, Podocarpus saligna and Embothrium coccineum covering not more than 15% of the whole catchment area. Both catchments are 13 km apart from each other.
The climate in the area of study is rainy temperate. In the meteorological station Isla Teja ( 25 m a.s.l.), 10 to 20 km away from the study sites, the mean annual temperature is 12.0˚C (January mean is 17˚C and July mean is 7.6˚C) and the mean annual precipitation is 2280 mm. Rainfall is concentrated in winter (May-August, 62%) and decreases strongly in summer (January-March, 9%). Soils at each catchment have approximately the same texture in the bottom of the 1 meter depth soil profile, but the top layers (0 to 15 cm; and 15 to 30 cm) have consistently 10% more clay, and 1% less sand in EP compared to NF soil profiles. Soils in these catchments are Andic Palehumult and Typic Paleudult, for NF and EP respectively. The main characteristic of these soils is that they are formed from volcanic ashes over a meteorized metamorphic complex [14] . In EP, soil clay content ranged between 37.2% - 45.1%, organic matter content ranges between 1.8% - 17.1%, inorganic-N (NO3-N + NH4-N) ranges between 9.8 - 21.0 mg∙kg−1, Ca2+ between 0.19 - 0.23 cmol∙kg−1 and Mg2+ ranges between 0.09 - 0.16 cmol∙kg−1. While, NF soil clay content ranges between 31.1% - 37.3% and organic matter content ranges between 5.9% - 17.8%, inorganic-N ranges between 11.2 - 57.4 mg∙kg−1, Ca2+ ranges between 0.23 - 1.32 cmol∙kg−1 and Mg2+ ranges between 0.10 - 0.71 cmol∙kg−1.
3. Methods
3.1. Sampling and Sample Analysis
Bulk precipitation was sampled using four plastic rain collectors attached to a 2.5-liter bottle. Bulk precipitation collectors (surface area 200 cm2) were installed in open areas (no trees were within 20 m of the sampling point), located between a distance of 100 - 500 m . Throughfall water was collected, using 4 collectors (surface area 254 cm2) which were installed under each type forest (evergreen and E. nitens plantation). All collectors were installed 1.2 m above the forest floor and installed inside opaque tubes in order to avoid light penetration that could promote algae growth. Throughfall collectors had a thin mesh at the beginning of the neck of the funnel, in order to prevent insects and leaves entering the collection bottles, and designed with a plastic ring in order to exclude bird droppings [15] . Soil water was sampled at two different depths (0.3 and 0.6 m) with low-tension porous-cup lysimeters (max 60 kPa of tension was applied) (Soil Moisture equipment corp. Santa Barbara, CA., USA).
Discharge from each catchment was constantly measured by a pressure transducer paired with a baro diver (Schlumberger Water Services). We sampled 5 rainfall events during the period March-November 2013 (Table 1). However, in this work we present detailed data from the events of April 4th (2nd event) and August 2nd (4th event) corresponding to events occurring at the end of dry season, and to mid rainy season respectively.
Water samples were taken directly from the streams with an ISCO-6712 automatic sampler
in each catchment. Stream samples were composed by two 250 mL aliquots taken each
30 minutes (1 h compound sample per bottle). Samples were filtered through a borosilicate
glass filter (Whatman) of 0.45 µm, and were determined for: NH4-N
using the phenate method (blue indophenol), and NO3-N as (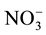 -N +
-N +
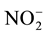 -N) using the
cadmium reduction method,
-N) using the
cadmium reduction method,
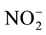 -N was always
below detection limit (DL), which was 1.5 μg∙L−1, for nitrate, nitrite
and ammonia. Dissolved Inorganic Nitrogen (DIN) was calculated as follows: DIN =
-N was always
below detection limit (DL), which was 1.5 μg∙L−1, for nitrate, nitrite
and ammonia. Dissolved Inorganic Nitrogen (DIN) was calculated as follows: DIN =
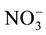 -N +
-N +
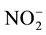 -N +
-N +
 -
N. Total dissolved nitrogen (Total-N) was determined by the sodium hydroxide and
persulfate digestion method (DL < 15 μg∙L−1). Organic nitrogen (Organic-N)
was calculated as follows: Organic-N = Total-N-DIN. Total phosphorous (Total-P)
was measured by the sodium hydroxide and persulfate digestion method (DL < 3
μg∙L−1) at LIMNOLAB (Limnology Laboratory, Universidad Austral de Chile).
-
N. Total dissolved nitrogen (Total-N) was determined by the sodium hydroxide and
persulfate digestion method (DL < 15 μg∙L−1). Organic nitrogen (Organic-N)
was calculated as follows: Organic-N = Total-N-DIN. Total phosphorous (Total-P)
was measured by the sodium hydroxide and persulfate digestion method (DL < 3
μg∙L−1) at LIMNOLAB (Limnology Laboratory, Universidad Austral de Chile).
In order to separate water fluxes in pre-event and event water during a storm event, we used stream samples sampled each hour with an ISCO-6512 automatic sampler; and 5 mm sequentially sampled throughfall using a modified version of the passive sequential sampler by [16] , during the events. If two end members have a distinct difference in their isotopic signature, the stormflow hydrograph can be separated in their contributions based on a mass balance approach [17] :

Table 1. Mean discharge (Q), concentrations of dissolved inorganic nitrogen (DIN), organic nitrogen (Organic-N), total nitrogen (Total-N) and total phosphorus (Total-P) for the rainfall events in old growth native forest (NF) and Eucalyptus nitens plantation (EP).
 (1)
(1)
 (2)
(2)
 (3)
(3)
where QT is the streamflow, QP is the contribution from pre-event water, QE is the contribution of event water, CT, CP and CE are the δ values of streamflow, pre-event water and event water, and FP is the fraction of pre- event water in the stream. Abundance of stable water isotopes is based on the isotopic ratios (18O/16O). The abundance is reported in the δ notation and often expressed as parts per thousand (‰ or per mil). δ18O values were determined on a Picarro Cavity Ring-Down Spectrometer (CRDS) L2120-i. Standard deviations were equal to or lower than 0.03‰ for δ18O. All isotopic analyses were made in ISOFYS Laboratory, Ghent University, Belgium.
The contributions of event and pre-event water can be determined based on Equation (3). The equation is constrained so that CT falls between CP and CE and that QP and QE are between zero and QT. Several assumptions underlie Equations (1) and (2):
1) The isotopic content of event and pre-event water are significantly different.
2) The event water maintains a constant isotopic signature in space and time, or any variations can be accounted for.
3) The isotopic signature of the pre-event water is constant in space and time, or any variations can be accounted for.
4) Contributions from the vadose zone must be negligible, or the isotopic signature of the soil water must be similar to that of groundwater.
5) Surface storage contributes minimally to the streamflow.
3.2. Data Analysis
We used Spearman correlations and also fitted models to determine whether catchment
discharge or new water contributions had an influence on nutrient concentration.
Then, catchment discharge and nutrient concentrations during the study period and
each event were plotted in order to observe the behavior in the increase, decrease
and peak flows. Sigmaplot 12.5 (Systat Software, inc.) was used for all regressions.
Statistical differences were considered if p ≤ 0.05. Since Total-N concentration
was almost 95% conformed by Organic-N, we decided to plot only
 -N,
DIN, Total-N and Total-P.
-N,
DIN, Total-N and Total-P.
4. Results and Discussion
4.1. Canopy Effects on Nutrient Concentration
There was a trend for Total-N and Total-P concentrations to be higher in throughfall than in bulk precipitation for both NF and EP (Figure 2). The enrichment concentration ratios of throughfall to precipitation were as follows: 3.03 and 4.48 for Total-N and Total-P, respectively, in NF; and 1.43 (Total-N) and 2.31 (Total-P) in EP. This enrichment is due to two processes: the washing off of the unquantified N input by dry deposition, on the one hand, and the N uptake from wet, dry particulate and gaseous deposition by leaves, twigs, stem surfaces, and lichens, on the other hand [18] . The old-growth evergreen forests are multi-stratified and have an understory of high diversity, resulting in a complex and diverse structure and species composition. The particular structural features of old-growth forests in southern Chile include the presence of old individuals of late-successional species, canopy gaps, and large-sized coarse woody detritus [19] .
Total-N concentrations from soil water at 30 and 60 cm were higher in NF (1125 and 1265 µg∙L−1, respectively), and EP plantation (399 µg∙L−1, for 30 cm only) than in bulk precipitation (169 µg∙L−1). Also, Total-P concentrations in soil water at 30 and 60 cm depth were higher in NF (172 and 157 µg∙L−1, respectively) and EP plantation (57 µg∙L−1, for 30 cm only) than in bulk precipitation (18 µg∙L−1). The same pattern was found in water soil infiltration at 10 cm depth in a native Nothofagus obliqua forest and in a Pinus radiata plantation located at the Central Valley of southern Chile (40˚S) [20] . Compared to bulk precipitation, the Total-N increased in both stands by passing through the humus and soil layers. This can be attributed to mineralization, nitrification and plant uptake occurring in the top soil [20] . There was a general trend for lower Total-N (127 µg∙L−1 and 100 µg∙L−1, for NF and EP respectively) and Total-P concentrations (11.1 µg∙L−1 and 11.0 µg∙L−1 for NF and EP,

Figure 2. Total-N and Total-P concentrations in rainfall, throughfall, soil water and streams for old growth native forest (NF) and Eucalyptus nitens plantation (EP).
respectively) in streams than in bulk precipitation (Total-N: 169 µg∙L−1; Total-P: 18.1 µg∙L−1) (Figure 2).
4.2. Relationships between Discharge and Nutrient Concentration
Table 1 summarizes measured concentrations for stream discharge and nutrient concentrations for the different rainfall events. In both catchments, the highest values of discharge and nutrients concentration were observed during the 3rd event (June 21-22, 2013). In NF, mean discharge was 17.8 ± 10.3 L seg−1. On average, Organic-N amounted to 392.0 ± 315.6 µg∙L−1, whereas DIN was only 19.3 ± 3.8 µg∙L−1 resulting in a Total-N concentration of 411.4 ± 313.7 µg∙L−1; while Total-P was 37.0 ± 37.0 µg∙L−1 (Table 1). In EP, mean discharge was 20.1 ± 15.5 L∙s−1. Organic-N was much higher than inorganic nitrogen (363.9 ± 560.7 µg∙L−1 and 28.7 ± 9.9 µg∙L−1, respectively) (Table 1). The lowest values of discharge and nutrient concentration were in the 2nd event (April 4-5, 2013). On Table 2, we summarized the Spearman correlation (r values) and model fitting analysis (adjusted r2) for the 2nd and 4th events, corresponding to the end of dry (April 4-5, 2013) and in middle of the rain seasons (August 2, 2013), respectively.
In general, Total-N (and Organic-N) and Total-P showed an increase in concentration with increasing discharge and were best described by exponential and linear models for 2nd and 4th event respectively. Catchment discharge showed higher correlation and r2 values compared to new water (see Table 2, for details). This means that Total-N and Total-P shows an enhanced hydrological access on 2nd and 4th events. Presumably this behavior is due to the fact that the majority of Organic-N reaching the stream is present in the mobile water compartment (or big pore water). The slope decrease from 2nd to 4th event could be the effect of several rainfall events previous the 4th event. We hypothesized that these relations were due to these nutrients were more concentrated in mobile (or big pore) water in soil, and not retained by soil particles or by biological activity.
According to the “piston flow” theory, during storm events, water reaching the soil pushes mobile soil water into the stream. This explains the stronger relations that Total-N (and Organic-N) and Total-P have with catch-
Table 2. Results of Spearman correlations (r); and of best fitted models adjusted r2 (Adj r2): Linear models (L, y = y0 + a × x); 2 parameter exponential decay (ED2, y = a × e(−b×x)), and 3 parameter exponential decay (ED3, y = y0 + a × e(−b×x)), for old growth native forest (NF) and Eucalyptus nitens plantation (EP).
ment discharge during the 2nd event, on both catchments. Nevertheless, the 4th event shows that Total-N and Total-P concentrations are highly related to both, catchment discharge (r = 0.86; p < 0.001 and r = 0.93; p < 0.001, for Total-N and Total-P respectively), and new water (r = 0.92; p < 0.001 and r = 0.95; p < 0.001, for Total-N and Total-P respectively) in EP. This was not observed in NF, where the relation between N and P was explained mainly by catchment discharge (r = 0.60; p < 0.01 and r = 0.74; p < 0.001, for Total-N and Total-P respectively), than new water (r = 0.37; ns and r = 0.46; p < 0.05, for Total-N and Total-P respectively). The higher relation and slopes of the model, shown by Total-P vs catchment discharge and new water in EP, could reflect higher erosion rates that are taking place in EP, but not in NF. Several studies have described high erosion rates in Eucalyptus spp. covered catchments [5] . [21] described that 85% and 79% of exported sediment was coming from the stream bed in an E. nitens (100% coverage), and an E. nitens and Pinus spp. (66% and 33% of catchment cover respectively) covered catchment in a nearby study site.
 -N
and DIN, on the other hand, showed a different behavior, being best fitted with
exponential decay models (Table 2), showing a clear
dilution behavior. During 2nd event
-N
and DIN, on the other hand, showed a different behavior, being best fitted with
exponential decay models (Table 2), showing a clear
dilution behavior. During 2nd event
 -N
and DIN concentrations were best explained by new water and by an exponential decay
models in both catchments. However, the model fitting was similar only for
-N
and DIN concentrations were best explained by new water and by an exponential decay
models in both catchments. However, the model fitting was similar only for
 -N
(adjusted r2 = 0.59; p < 0.001 and adjusted r2 = 0.57;
p < 0.001, for EP and NF respectively), but not for DIN (adjusted r2
= 0.43; p < 0.01 and adjusted r2 = 0.08; ns, for EP and NF catchments
respectively). During the 4th event, in EP,
-N
(adjusted r2 = 0.59; p < 0.001 and adjusted r2 = 0.57;
p < 0.001, for EP and NF respectively), but not for DIN (adjusted r2
= 0.43; p < 0.01 and adjusted r2 = 0.08; ns, for EP and NF catchments
respectively). During the 4th event, in EP,
 -N
and DIN concentrations were more related with catchment discharge (r = 0.51; p <
0.01 and r = 0.72; p < 0.001, respectively), rather new water (r = 0.14; ns and
r = 0.43; p < 0.05 for
-N
and DIN concentrations were more related with catchment discharge (r = 0.51; p <
0.01 and r = 0.72; p < 0.001, respectively), rather new water (r = 0.14; ns and
r = 0.43; p < 0.05 for
 -N
and DIN respectively). However, native evergreen forest showed the opposite behavior
-N
and DIN respectively). However, native evergreen forest showed the opposite behavior
 -N
was highly related to new water (r = 0.92; p < 0.001) than catchment discharge
(r = 0.68; p < 0.001). DIN concentrations showed the same behavior as
-N
was highly related to new water (r = 0.92; p < 0.001) than catchment discharge
(r = 0.68; p < 0.001). DIN concentrations showed the same behavior as
 -N,
being more related to new water than catchment discharge (r = 0.92; p < 0.001;
r = 0.67; p < 0.001, for new water and discharge respectively).
-N,
being more related to new water than catchment discharge (r = 0.92; p < 0.001;
r = 0.67; p < 0.001, for new water and discharge respectively).
The dilution observed for
 -N
and DIN, as catchment discharge and new water increases suggests that these nutrients
have a strong internal source. Even though, throughfall is highly enriched in
-N
and DIN, as catchment discharge and new water increases suggests that these nutrients
have a strong internal source. Even though, throughfall is highly enriched in
 -N
and DIN, it is known that inorganic forms of nitrogen are used in several biological
processes occurring at soil level. Also, [22] described that acid soils formed by
volcanic ashes had the capability of retaining anions, like
-N
and DIN, it is known that inorganic forms of nitrogen are used in several biological
processes occurring at soil level. Also, [22] described that acid soils formed by
volcanic ashes had the capability of retaining anions, like
 and
and .
On the other hand, [11] described that DIN inputs did not end up in the soil water
compartment (mobile water). If the rate of
.
On the other hand, [11] described that DIN inputs did not end up in the soil water
compartment (mobile water). If the rate of
 -N
and DIN supply remains relatively unchanged during precipitation events, we expect
these elements to be diluted by the enhanced throughput of water, therefore we could
say that
-N
and DIN supply remains relatively unchanged during precipitation events, we expect
these elements to be diluted by the enhanced throughput of water, therefore we could
say that
 -N
and DIN are either highly consumed by microorganisms or is being retained by soil
particles. This last suggests that the accessibility of moving water to
-N
and DIN are either highly consumed by microorganisms or is being retained by soil
particles. This last suggests that the accessibility of moving water to
 -N
and DIN is difficult, maybe due to the fact that is strongly attached to soil particles.
-N
and DIN is difficult, maybe due to the fact that is strongly attached to soil particles.
Figure 3 and Figure 4
show the relationship between catchment discharge and nutrient concentration for
the 2nd event and for the 4th event. The correlations between
nutrient fractions and catchments discharge were significant in both catchments
for most of the nutrients (see Table 2). In general,
when discharge increases also Total-N and Total-P concentrations increase and this
is generally observed during all events (data not shown). Total-N increases are
mostly due to Organic-N concentrations increase and for all N forms, except for
DIN in NF (Figure 3). On the other hand, [23] observed
both positive and negative correlations between stream discharge and NO3-N
in an old-growth evergreen rainforest located at Andean mountain range (40˚S) which
was attributed to differences in peak flow. Nitrate has different behaviors during
storm events because the streamflow can dilute the nitrate at the peak flows, but
when the streamflow increases slowly, usually the nitrate also increases [23] .
Our results show that when stream discharge increases,
 -N
concentrations decreases (Figure 3 and Figure 4). Organic-N flushes, along with the dilution of
-N
concentrations decreases (Figure 3 and Figure 4). Organic-N flushes, along with the dilution of
 -N,
have been previously described in literature [24] [25] .
-N,
have been previously described in literature [24] [25] .
4.3. Total-N and Total-P Concentrations in Stream Water for Forest Ecosystems of Southern Chile
Total-N and Total-P concentrations in stream water are variable in forest ecosystems of southern Chile (see Table 3). In general, the highest values of Total-N and Total-P concentrations are in Fitzroya cuppressoides forest (176.5 µg Total-N L−1) located in Coastal mountain range and in Nothofagus pumilio forest (67.3 µg Total-P L−1) located in Andean mountain range, while the lowest values are found in an evergreen forest (36.8 µg Total-N L−1), located in Coastal range and in Fitzroya cuppresoides forest (4.6 µg Total-P L−1) located in the Coastal

Figure 3. Nutrient concentrations
( -N,
DIN, Total-N and Total-P) vs catchment dicharge (left side) and new water (L seg−1)
(right side), for both studied catchments during the 2nd event: old growth
native forest (NF), in black round dots and Eucalyptus nitens plantation (EP) in
white inverted triangles.
-N,
DIN, Total-N and Total-P) vs catchment dicharge (left side) and new water (L seg−1)
(right side), for both studied catchments during the 2nd event: old growth
native forest (NF), in black round dots and Eucalyptus nitens plantation (EP) in
white inverted triangles.

Figure 4. Nutrient concentrations
( -N,
DIN, Total-N and Total-P) vs catchment dicharge (figures on the left side) and new
water (figures on the right side), both on L∙s−1 for the 4th
event. Old growth native forest (NF), in black round dots and Eucalyptus nitens
plantation (EP) in white inverted triangles.
-N,
DIN, Total-N and Total-P) vs catchment dicharge (figures on the left side) and new
water (figures on the right side), both on L∙s−1 for the 4th
event. Old growth native forest (NF), in black round dots and Eucalyptus nitens
plantation (EP) in white inverted triangles.
Table 3. Mean concentrations (µg∙L−1) of total nitrogen (Total-N) and total phosphorous (Total-P) in stream water for different forest ecosystems under a low-deposition climate, southern Chile.
nd = not determined.
range. Concentrations of inorganic-N were smaller than Organic-N concentration in NF (33.2 µg DIN L−1 versus 94.4 µg Organic-N L−1) and EP (33.6 µg DIN L−1 versus 67.0 µg Organic-N L−1), in agreement with previous research in southern Chile [9] [10] , demonstrating that dissolved organic nitrogen is responsible for the majority of nitrogen losses from unpolluted forest ecosystems. According to [3] , the DIN: DON ratio is smaller in evergreen native forests than in exotic plantation forests.
5. Conclusions
We conclude that the native evergreen forest shows higher enrichment concentration ratios of throughfall to precipitation with respect to Eucalyptus plantation. The differences in enrichment are attributed to the multi-strati- fied canopies and an understory of high diversity in the evergreen forests resulting in a complex and diverse structure and species composition. The Total-N and Total-P concentrations increased in both stands, especially in native evergreen forest, by passage through by the humus and upper soil layer (30 and 60 cm depth).
DIN and
 -N
showed a dilution during the 2nd event and hydrologically constant as
discharge and/or new water apportionment increased during the 4th event.
Since our study sites are in a region which is constantly under nutrient limitation,
with very low N inputs (<3 kg∙ha−1∙yr−1), forest ecosystems
in this region have developed strategies of high nutrient retention. In this case
-N
showed a dilution during the 2nd event and hydrologically constant as
discharge and/or new water apportionment increased during the 4th event.
Since our study sites are in a region which is constantly under nutrient limitation,
with very low N inputs (<3 kg∙ha−1∙yr−1), forest ecosystems
in this region have developed strategies of high nutrient retention. In this case
 -N
and DIN are rapidly retained (biotic or abiotic) by the soil ecosystem. Both catchments
showed very similar behaviors on all measured nutrients. However, it was clear that
during the 4th event, DIN (therefore
-N
and DIN are rapidly retained (biotic or abiotic) by the soil ecosystem. Both catchments
showed very similar behaviors on all measured nutrients. However, it was clear that
during the 4th event, DIN (therefore
 -N)
and Total-P showed higher concentrations in Eucalyptus nitens than native evergreen
forest. Nevertheless, the 2nd event showed similar concentrations at
catchment discharge for nutrients like Total-N, DIN and Total-P. These results support
what [11] described for
-N)
and Total-P showed higher concentrations in Eucalyptus nitens than native evergreen
forest. Nevertheless, the 2nd event showed similar concentrations at
catchment discharge for nutrients like Total-N, DIN and Total-P. These results support
what [11] described for
 -N
and Organic-N losses in volcanic soil catchments.
-N
and Organic-N losses in volcanic soil catchments.
Total-N and Total-P concentrations were more related to catchment discharge and not to new water. During the 4th event the pattern repeats only for NF. Whether in EP, Total-N and Total-P concentrations are more related to new water rather than catchment discharge. This is expected since Eucalyptus covered catchments are described and known to have low water infiltration rates, in addition P is attached to soil particles. This relation is sustained actually by the soil erosion during the 4th event.
We would like also to address that even though, we only measured throughfall nutrient concentrations, our results indicate that Total-N (DIN and Organic-N) are highly controlled by atmospheric inputs. However, we should start to observe whether other inputs (i.e. bacterial atmospheric fixation) of nitrogen are taking place in the understory of native forest and plantations (Pinus spp., and Eucalyptus spp.). Only then we will have a clearer picture of what is happening inside these forest ecosystems. So far, most of the differences between native forests and exotic plantations are exemplified by different water infiltration rates (higher in native forests), soil erosion (lower in native forests) and nutrients (higher exportation, especially for nitrate and Total-P, in exotic plantations). Further studies need to be done in order to unravel the different pathways and sources of nitrogen and phosphorous in this complex ecosystems in which one is decreasing and the other one is growing each year.
Acknowledgements
This study was financed by FONDECYT (Fondo Nacional de Ciencia y Tecnología) Project 1120188. Also Mr. Hervé-Fernandez would like to thank Mr. C. Brumbt, Programa de Personal Avanzado CONICYT, Becas Chile and Commissie Wetenschappelijk Onderzoek, Ghent University.
References
- Zhao, M., Zeng, C., Liu, Z.H. and Wang, S.J. (2010) Effect of Different Land Use/Land Cover on Karst Hydrogeochemistry: A Paired Catchment Study of Chenqi and Dengzhanhe, Puding, Guizhou, SW China. Journal of Hydrology, 388, 121-130. http://dx.doi.org/10.1016/j.jhydrol.2010.04.034
- Cannel, M. (1999) Environmental Impacts of Forest Monocultures: Water Use, Acidification, Wildlife Conservation, and Carbon Storage. New Forests, 17, 239-262. http://dx.doi.org/10.1023/A:1006551018221
- Oyarzún, C., Godoy, R., Aracena, A., Rutherford, P. and Deschrijver, A. (2007) Effects of Land Use Conversion from Native Forest to Exotic Plantations on Nitrogen and Phosphorus Retention in Catchments of Southern Chile. Water, Air and Soil Pollution, 179, 341-35. http://dx.doi.org/10.1007/s11270-006-9237-4
- Uyttendaele, G. and Iroumé, A. (2002) The Solute Budget of a Forest Catchment and Solute Fluxes within a Pinus radiata and a Secondary Native Forest Site, Southern Chile. Hydrological Processes, 16, 2521-2536. http://dx.doi.org/10.1002/hyp.1046
- Oyarzún, C., Frene, C., Lacrampe, G., Huber, A. and Hervé, P. (2011) Propiedades hidrológicas del suelo y exportación de sedimentos en dos microcuencas de la Cordillera de la Costa en el sur de Chile con diferente cobertura vegetal. Bosque, 32, 10-19. http://dx.doi.org/10.4067/S0717-92002011000100002
- Salmon, C., Water, M.T., Hedin, L. and Brown, M. (2001) Hydrological Controls on Chemical Export from and Undisturbed Old-Growth Chilean Forest. Journal of Hydrology, 253, 69-80. http://dx.doi.org/10.1016/S0022-1694(01)00447-4
- Armesto, J., Smith-Ramírez, C., Carmona, M., Celis-Diez, J.L., Díaz, I., Gaxiola, A., Gutiérrez, A.G., Nuñez-Avila, M., Pérez, C. and Rozzi, R. (2009) Old-Growth Temperate Rain Forests of South America: Conservation, Plant-Animal Interactions, and Baseline Biogeochemical Processes. In: Wirth, C., Gleixner, G. and Heiman, M., Eds., Old-Growth Forests. Function, Fate and Value. Ecological Studies, Vol. 207, Springer Berlin, Heidelberg, 367-390.
- Huygens, D., Rütting, T., Boeckx, P., Van Cleemput, O., Godoy, R. and Müller, C. (2007) Soil Nitrogen Conservation Mechanisms in a Pristine South Chilean Nothofagus Forest Ecosystem. Soil biology and Biogeochemistry, 39, 2448-2458. http://dx.doi.org/10.1016/j.soilbio.2007.04.013
- Oyarzún, C., Godoy, R., Deschrijver, A., Staelens, J. and Lust, N. (2004) Water Chemistry and Nutrient Budgets in an undisturbed Evergreen Rainforest of Southern Chile. Biogeochemistry, 71, 107-123. http://dx.doi.org/10.1007/s10533-004-4107-x
- Perakis, S. and Hedin, L. (2002) Nitrogen Loss from Unpolluted South American Forest Mainly via Dissolved Organic Compounds. Nature, 415, 416-419. http://dx.doi.org/10.1038/415416a
- Huygens, D., Boeckx, P., Templer, P., Paulino, L., Van Cleemput, O., Oyarzún, C., Müller, C. and Godoy, R. (2008) Mechanisms for Retention of Bioavailable Nitrogen in Volcanic Rainforest Soils. Nature Geosciences, 1, 543-548. http://dx.doi.org/10.1038/ngeo252
- Huber, A., Iroumé, A., Mohr, C. and Frêne, C. (2010) Effect of Pinus radiata and Eucalyptus globulus Plantations on Water Resource in the Coastal Range of Biobío Region, Chile. Bosque, 31, 219-230. http://dx.doi.org/10.4067/S0717-92002010000300006
- Iroumé, A. and Palacios, H. (2013) Afforestation and Changes in Forest Composition Affect Runoff in Large River Basins with Pluvial Regime and Mediterranean Climate, Chile. Journal of Hydrology, 505, 113-125. http://dx.doi.org/10.1016/j.jhydrol.2013.09.031
- Salazar, O., Casanova, M. and Luzio, W. (2005) Correlation between World Reference Base and Soil Taxonomy for the Soils from Xth Los Lagos Region of Chile. Soil Science and Plant Nutrition, 5, 35-45.
- Kleemola, S. and Soderman, G. (1998) Manual for Integrated Monitoring. International Co-Operative Programme on Integrated Monitoring on Air Pollution Effects. Environmental Report 5. Environment Data Centre, National Board of Waters and the Environment, Helsinki.
- Vermette, S. and Drake, J. (1986) Simplified Wet-Only and Sequential Fraction Rain Collector. Atmospheric Environment, 21, 715-716. http://dx.doi.org/10.1016/0004-6981(87)90053-9
- McDonnell, J., Bonnell, M., Stewart, M. and Pearce, A. (1990) Deuterium Variations in Storm Rainfall: Implications for Stream Hydrograph Separation. Water Resources Research, 26, 455-458. http://dx.doi.org/10.1029/WR026i003p00455
- Staelens, J., Godoy, R., Oyarzún, C., Thibo, K. and Verheyen, K. (2005) Nitrogen Fluxes in Throughfall and Litterfall in Two Nothofagus Forests in Southern Chile. Gayana Botánica, 62, 63-71. http://dx.doi.org/10.4067/S0717-66432005000200002
- Donoso, P., Frêne, C., Flores, M., Moorman, M., Oyarzún, C. and Zavaleta, J. (2009) Balancing Water Supply and Old-Growth Forest Conservation in the Lowlands of South-Central Chile through Adaptive Co-Management. Land- scape Ecology, 29, 245-260. http://dx.doi.org/10.1007/s10980-013-9969-7
- Oyarzún, C., Godoy, R., Staelens, J., Aracena, C. and Proschle, J. (2005) Nitrogen Fluxes in a Nothofagus obliqua Forest and a Pinus radiata Plantation in the Central Valley of Southern Chile. Gayana Botánica, 62, 88-97. http://dx.doi.org/10.4067/S0717-66432005000200004
- Schüller, P., Walling, D., Iroumé, A., Quilodrán, C., Castillo, A. and Navas, A. (2013) Using Cs-137 and Pb-210ex and Other Sediment Source Fingerprints to Document Suspended Sediment Sources in Small Forested Catchments in South-Central Chile. Journal of Environmental Radioactivity, 124, 147-159. http://dx.doi.org/10.1016/j.jenvrad.2013.05.002
- Strahm, B. and Harrison, R. (2006) Nitrate Sorption in a Variable-Charge Forest Soil of the Pacific Northwest. Soil Science, 171, 313-321. http://dx.doi.org/10.1097/01.ss.0000209355.76407.16
- Lozano, A. (2011) Relación entre el caudal y la exportación de nutrientes durante otoño en un bosque templado lluvioso en la Cordillera de los Andes, sur de Chile. Master’s Thesis in Water Resources, Universidad Austral de Chile, Valdivia, 51 p.
- Cirmo, C. and McDonnell, J. (1997) Linking the Hydrologic and Biogeochemical Controls of Nitrogen Transport in Near-Stream Zones of Temperate-Forested Catchments: A Review. Journal of Hydrology, 199, 88-120. http://dx.doi.org/10.1016/S0022-1694(96)03286-6
- Inamdar, S. and Mitchell, M. (2007) Storm Event Exports of Dissolved Organic Nitrogen (DON) across Multiple Catchments in a Glaciated Forested Watershed. Journal of Geophysical Research, 112, 1-18. http://dx.doi.org/10.1029/2006JG000309
- Godoy, R., Oyarzún, C. and Gerding, V. (2001) Precipitation Chemistry in Deciduous and Evergreen Nothofagus Forest of Southern Chile under a Low-Deposition Climate. Basic and Applied Ecology, 2, 65-72. http://dx.doi.org/10.1078/1439-1791-00037
- Oyarzún, C., Godoy, R. and Sepulveda, A. (1998) Water and Nutrient Fluxes in a Cool Temperate Rainforest at the Cordillera de la Costa in Southern Chile. Hydrological Processes, 12, 1067-1077. http://dx.doi.org/10.1002/(SICI)1099-1085(19980615)12:7<1067::AID-HYP641>3.0.CO;2-O
- Oyarzún, C.E., Campos, H. and Huber, A. (1997) Exportación de nutrientes en microcuencas con distinto uso del suelo en el sur de Chile (Lago Rupanco, X Región). Revista Chilena de Historia Natural, 70, 507-519.
- Oyarzún, C. and Huber, A. (2003) Nitrogen Export from Forested and Agricultural Watersheds of Southern Chile. Gayana. Botánica, 60, 63-68. http://dx.doi.org/10.4067/S0717-66432003000100010
- Little, C., Soto, D., Lara, A. and Cuevas, J. (2008) Nitrogen Exports at Multiple-Scales in a Southern Chilean Water- shed (Patagonian Lakes District). Biogeochemistry, 87, 297-309. http://dx.doi.org/10.1007/s10533-008-9185-8