Open Journal of Ecology
Vol.06 No.12(2016), Article ID:71945,22 pages
10.4236/oje.2016.612065
Cyperus papyrus L. Growth Rate and Mortality in Relation to Water Quantity, Quality and Soil Characteristics in Nyando Floodplain Wetland, Kenya
P. J. K. Rongoei1*, N. O. Outa2
1Department of Environmental Science, Egerton University, Njoro, Kenya
2Department of Biological Sciences, Egerton University, Njoro, Kenya

Copyright © 2016 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: July 7, 2016; Accepted: November 8, 2016; Published: November 11, 2016
ABSTRACT
Cyperus papyrus (L.) growth rate and mortality is influenced by environmental conditions prevailing in the wetland. To assess growth dynamics of C. papyrus in relation to water depth and anthropogenic (exploitation) pressures, monthly and bi-monthly measurement of culm length and girth were done between June and December 2010 (period 1) and April to June 2011 (period 2). Three study sites were selected based on the water levels and livelihood-driven exploitation pressures. Surrogate measurements of individual culm height and girth were done in three 1 m2 quadrats in each site to determine the growth rate of papyrus. Water depth was lowest in period 2 (dry) and highest in period 1 (wet) which was related to the livelihood activities being highest in period two and lowest in period one. Culm mortality occurred throughout the study period with 64% due to natural senescing while insect/rodent accounted for 19%. Papyrus growth was higher in Singida (2.5 ± 0.2 cm/day) representing less disturbed site and least in Wasare (1.4 ± 0.1 cm/day) which was highly disturbed. Multiple regression models for culm length showed culm density, mean length and NH4 negatively influenced growth rate while site as a dummy variable, water depth, SRP and TP had positive effects on papyrus growth rate. Understanding growth rate and causes of mortality in papyrus is important to establish sustainable management strategies of this ecosystem to maintain its integrity.
Keywords:
Water Depth, Density, Multiple Regression Models, Lake Victoria Wetlands, Kenya

1. Introduction
Characteristics and conditions of papyrus marshes fringing Lake Victoria are affected by human activities as they play an important role in meeting the needs of the poor majority [1] [2] . Increase in human population has resulted in overexploitation of resources, intensive farming activities [3] that threaten the productivity of the wetland and affect its integrity. Understanding its growth in the face of hydrological and exploitation pressures is crucial to better estimate papyrus wetland value for proper management.
Nyando floodplain wetland experience periodic flooding and drying which influences biomass production and the health of the wetland. A slight change in hydrologic conditions may result in large changes in species richness and in turn lead to changes in ecosystem productivity [4] . Higher productivities can be achieved in areas with stable water regime where there is low density but with robust individual plants [5] . Other environmental conditions, such as wave exposure, nutrient availability and soil moisture, are some of the important factors determining occurrence and growth of papyrus [6] [7] [8] . For example, [9] showed that receding water levels can expose mud where seeds can germinate and sprout. Other studies have shown that water level in the wetland is closely related to the high production of papyrus [7] [10] .
Studies on primary productivity of papyrus-dominated wetlands have focused on destructive method for determining biomass production in natural intact papyrus marshes of Lake Naivasha [11] , Rubondo Island, Lake Victoria, Tanzania [10] and Winam Gulf, Lake Victoria, Kenya [12] . Knowledge of primary productivity of papyrus in Nyando is still inadequate. Therefore, un-destructive method to determine specific papyrus productivity is crucial in this wetland where hydrological regimes keep changing and exploitation pressures from livelihood activities increase. Papyrus productivity in relation to differences in exploitation activities are represented by the three study sites with different exploitation pressures and water levels [2] [4] . This study focused on papyrus aerial biomass (culm density and growth rate) which is important in determining the overall productivity of the system as they contribute more than 50% of the plant biomass [10] [13] [14] .
It is hypothesized that density and growth rate are important parameters to determine productivity of papyrus wetland. These characteristics are relied upon by the ecosystem functions such as nutrient regulation [15] , habitat availability [16] , and ecosystem services available to people who depend on wetland resources for their needs [2] . Understanding the growth rate of papyrus and density in relation to exploitation and environmental conditions prevailing in the wetland is important to give better quantitative estimates of regulating services of papyrus wetlands. These estimates will assist in planning for management and sustainable exploitation strategies of wetland resources. The specific objectives were to: (i) determine the water level, water quality and soil characteristics of the three transects of Nyando wetland; (ii) compare culm density, mortality and new culm shoots of papyrus among sites; (iii) determine the growth rate of C. papyrus L. in three transects of Nyando wetland in period 1 and period 2; (iv) evaluate the relationship between C. papyrus L. growth rate and environmental factors using multiple regression models.
2. Methods
2.1. Study Area
The study was carried out in a papyrus-dominated Nyando floodplain wetland in the north eastern part of Lake Victoria, Kenya (0˚11'S - 0˚19'S to 34˚47'É - 34˚57'É) (Figure 1). Due to lack of delineation coupled by encroachment and hydrological variability in

Figure 1. Location of Nyando wetland in relation to Lake Victoria showing the study sites.
the floodplain, the wetland area is not clearly known but has been estimated to fluctuate between 3000 to 5000 hectares [1] [17] .
The annual rainfall received at the Nyando basin is between 1000 and 1600 mm with long rains between March and May and short rains in October to December with an estimated mean temperature of 23˚C [18] . This results to seasonal fluctuation in water levels of the wetland with the peak during the wet season causing inundation and the lowest water level in the dry season. The wetland is dominated by papyrus vegetation with patches of other emergent macrophytes such as Vossia cuspidata, Typha sp., Ipomoea aquatica and Phragmites sp. [4] .
Three transects were identified based on differences on water levels which drive livelihood exploitation levels. The sites were located in Ogenya which was influenced by the lake and inundated throughout the study period. Singida was influenced by both lake and river water and represented undisturbed site. However, Wasare site was dry most of the monitoring period and influenced by river over-flow during the wet season. This site was highly disturbed as receded water opened the area for livelihood exploitation activities.
2.2. Water Sample Collection and Water Quality Analysis
In each study site, three quadrats of 1 m2 each were established in areas with less interference. The study was carried in two periods (period 1 and period 2). Period 1 covered June-December 2010 which was wet with dense, untouched papyrus stands and period 2 covered April-June 2011 which was dry with newly growing papyrus stands after disturbance. Three measurements of surface water depths were done in each quadrat using portable staff gauge. Physical parameters of surface water quality pH, dissolved oxygen (DO), temperature (T), and conductivity (EC) were measured insitu using WTW meters and probes (pH 3210, OXi 3210, Cond 3210), respectively. Water samples were collected in duplicates once every month for 13 months from the three quadrats. However, water level was zero in some quadrats during some months (Figure 2). Collected samples were labeled and placed in cool box with ice blocks and transported to Biological Sciences laboratory of Egerton University, Kenya.
While in the laboratory, 250 mls of surface water sample was immediately filtered on arrival through 0.45 μm glass-fiber filters. This was then used to analyze ammonium nitrogen (NH4-N) using sodium salicylate method, nitrite nitrogen (NO2-N) using colorimetric method, nitrate nitrogen (NO3-N) using sodium-salicylate method, and soluble reactive phosphorus (SRP) using ascorbic acid method. For non-filtered samples, total nitrogen (TN) and total phosphorus (TP) were analyzed using cadmium reduction and per-sulphate digestion methods, respectively [19] .
2.3. Soil Sample Collection and Analysis
In the field, soil samples were collected from about 5 cm below soil surface in the same 1 m2 quadrats using pre-weighed stainless steel ring core with determined dimensions which were labeled and covered with their caps. The soil samples in the stainless steel

Figure 2. Mean monthly water levels relative to the soil surface overtime in the intact wetland zones of the three study sites in Nyando wetland. Numbers are means of six measurements ± standard error in Wasare and nine measurements ± standard error in Ogenya and Singida. The study covered the period from May 2010 to June 2011. Water depth was measured using graduated portable staff gauge. Period 1 represents the wet season and period 2 represents dry season.
ring core were used to determine bulk density and percent soil moisture content. The dimensions of the core (diameter and height) were used to calculate the bulk density using modified formula from [20] :
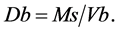 (1)
(1)
where Db is the bulk density while Ms is the mass of dry soil (g) and Vb are the dimensions of the core given as the total volume.
Soil moisture content was determined by weighing the fresh soil samples in the cores to get the wet weight of soil. The weight cores were placed in the oven at 105˚C for 24 hours to a constant weight. The cores were removed and re-weight to get the weight of dry soil. This was used to calculate % moisture content of the soil using the formula:
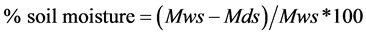 (2)
(2)
where Mws is the mass of wet soil (g), and Mds is the mass of dry soil (g).
A composite of five soil samples were collected from about 0 - 15 cm depth below soil surface around each quadrat using soil auger. Samples were collected at an interval of two months between June 2010 and June 2011. The five composite samples were mixed in a bucket and a sub-sample was put in a sealable plastic bag which was labeled and placed in a cool box for transportation to Egerton University, Biological Sciences laboratory for drying and Soil Sciences laboratory for analysis.
In the laboratory, sub-sample of composite soil was sun-dried for three days and oven dried at 60˚C for 48 hours to constant weight. The soil was homogenized with a pestle and a mortar and passed through 2 mm sieve. These samples were used to determine soil particle size using Bouyous Method and pH using electric pH meter. Total nitrogen and total phosphorus was determined using kjeltec auto distillation unit and molybdate-vanadate method, respectively [20] [21] .
2.4. Determining Papyrus Culm Density, New Culm Recruits and Mortality
The same three quadrats of 1 m2 were used to determine culm density, new shoots and dead culms. In each of the three quadrats established in each site, individual papyrus culms were counted to get the total number as culm density per unit area. New culm shoots were identified and recorded in each quadrat in every visit. Causes of mortality were monitored by recording observed changes on papyrus culm and umbel in period 1 and 2. Causes of mortality were classified into four categories as: (i) culm with no marks but senesced from the umbel was termed as natural cause (N); (ii) culm with no umbel but with sharp edge represented a cut umbel (C); (iii) culm with no umbel or with umbel but broken was categorized as grazed (G); (iv) culm with marks just above the rhizome or along the culm represented the group that were damaged by insects or rodents (IR). At the end of each study period the numbers of live culms were recorded.
2.5. Measuring Papyrus Growth Rate
Measuring of papyrus culm length and girth was carried out in period 1 and 2. Papyrus was measured in period 1 on a monthly basis with an interval of days ranging from 27- 36 (July to December 2010) and 14 days interval in period 2 (April to June 2011). In period 2, regeneration experiment was conducted in Singida and Ogenya while Wasare site was excluded due to human interference (burning, harvesting and animal grazing).
In each quadrat where water samples were collected, individual culms were marked with a unique number (identification code) using permanent marker. The length of each culm was measured above the rhizome from end of scale leaf to the apex of the umbel while the girth was measured just above the scale leaf according to [22] . The difference between the final measurement and the initial in each visit was divided by the number of days to get specific growth of each culm in cm/day. New shoot was marked with a different number and their growth was followed from July to December 2010 in period 1 and from April to June 2011 for period 2.
The relationship between papyrus culm length and absolute growth was calculated using the formula:
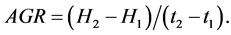 (3)
(3)
where AGR is the absolute growth rate in cm/day, H2 is the culm length at time t2 and H1 is the initial culm length at time t1. The average length was determined by adding initial length at t1 to length at t2 and dividing it by two.
Relative growth rate (RGR) is the growth rate relative to the size of the plant and was determined using the formula:
 (4)
(4)
where AGR is the absolute growth given in Equation (3) and Hav is the average culm length (cm).
To estimate the parameters of logistic equation, data was sorted by excluding negative growth from a total of 953 to 723. This was then used to determine the maximum culm length in each study site from the logistic model. From the remaining data, linear regression analysis was done for data from each site focusing on maximum growth rate. The slope and the intercept with the known average length were used to calculate estimated maximum culm length (k) of the logistic model in each site using the formula;
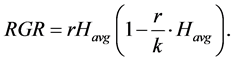 (5)
(5)
where RGR is the relative growth rate per day, r is a constant which is equal to relative growth rate at lower values of Havg (intercept), k is the maximum measured value of Havg (maximum length) and r/k is the slope. The same was done for culm girth by modifying the above formula to;
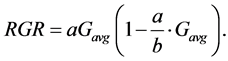 (6)
(6)
where G is the papyrus culm girth, a is a constant while b is the maximum measured value of Gavg (maximum girth) and a/b is the slope.
2.6. Data analysis
Prior to parametric tests, data was checked for conformity to normal distribution using Shapiro-Wilk test. As most data were not normally distributed, Kruskal-Wallis one way non-parametric ANOVA (KW-ANOVA) was used to determine differences between sites and period of study. This was followed by Kruskal-Wallis multiple comparison tests for post-hoc evaluation. Descriptive statistics was used to analyze the physico- chemical parameters in water and soil. The relationship between population dynamics (culm density, new culms and dead culms per square meter) and water depth in the quadrats was assessed using Spearman's rho rank correlation. To explore variations among sites, a comparison of relative growth rate of papyrus was done. Furthermore, relationship between culm length and girth was determined by a linear regression analysis after data was log-transformed.
The relationship between papyrus growth rate and habitat conditions was determined using a data set of 723 for papyrus culm length and 733 for papyrus girth. Correlation matrix was used for all data set from each transect in order to determine if there was a significant relationship between predictor variables (r > 0.50). The variables that were not significantly related (r < 0.50) were then used for five multiple linear regression models that were divided according to their average length (~0 - 200 cm and > 201 cm ) in each period. This resulted in three models for culm length and two models for culm girth (culm girth data not included here). Soil characteristics, surface water depth and water quality parameters were used as independent variables for explaining papyrus growth using multiple linear regression models (
3. Results
3.1. Water Depth, Water Quality and Soil Characteristics in the Study Sites
Wetland surface water levels differed among the study sites. Mean monthly variation in surface water depths showed Singida to be highest with a mean of 76.7 ± 4.8 cm, followed by Ogenya (44.1 ± 2.5 cm) and Wasare (38.7 ± 4.8 cm). In Ogenya, water levels remained above the soil surface for most part of the study period (Figure 2). However, water depth in Singida declined to 3 cm in December 2010 and 94 cm by March 2011 below soil surface while Wasare declined to below soil surface by 24 cm in December 2010 to 253 cm by June 2011. Therefore, Wasare surface water level was lowest as compared to Singida and Ogenya (Х2 = 40.2, df = 2, P < 0.001).
Figure 3 summarizes the water quality characteristics of the three study sites. Surface water pH, dissolved oxygen and conductivity did not show differences among sites. However, water temperature was generally higher in Ogenya compared to Wasare and Singida (X2 = 17.4, df = 2, P < 0.001).
Figure 4 shows the nutrient concentrations of wetland surface water quality. Nutrients (NO2, NO3, TN, SRP and TP) showed the same trend in all the sites although different in the months of measurements. However, TSS in Wasare was high compared to Singida (X2 = 11.4, df = 2, P < 0.01) while NH4 was higher in Wasare than Ogenya (X2 = 7.74, df = 2, P = 0.02).
Wetland soil pH ranged between 4.6 and 4.8 in Wasare and was lower than in Ogenya

Figure 3. Monthly changes in physical quality of surface water in the three sites of Nyando wetland with different water levels covering the period of June 2010 to June 2011. Values are means ± standard error (SE) (n = 3).

Figure 4. Monthly changes in chemical quality of surface water in the three sites of Nyando wetland with different water levels covering the period of June 2010 to June 2011. Values are means ± standard error (SE) (n = 2).
and Singida (X2 = 47.4, df = 2, P < 0.001). Proportion of clay in Singida wetland soil was higher and lowest in Wasare although no significant differences were observed among study sites (Table 1). Also, the proportion of sand was different among the sites (X2 = 26.1, df = 2, P < 0.001) and was higher in Ogenya as compared to Singida and Wasare. Silt proportion was higher in Wasare compared to Ogenya and Singida (X2 = 11.7, df = 2, P < 0.01) (Table 1). Total phosphorus in soil was lowest in Wasare (X2 = 16.8, df = 2, P < 0.001) but not the case with Ogenya and Singida.
3.2. Density, New Culm Shoot Recruitment and Mortality in Papyrus Stands
Papyrus culm densities differed among sites with Wasare having the highest density of 37 ± 13 culms/m2 compared to 26 ± 11 and 15 ± 4.5 culms/m2 in Ogenya and Singida, respectively. Kruskal-Wallis test showed that there was a significant difference between papyrus culm density in Wasare and Ogenya and Wasare and Singida (X2 = 19.5, df = 2, P < 0.001) but no differences observed between Singida and Ogenya.
The recruitment of new culm shoots was more in the month of September in all the sites than the other months with Wasare having the highest number (8 ± 2 culms/m2) followed by Singida (5 ± 1.6 culms/m2) and Ogenya (3 ± 2 culms/m2) (Figure 5). Culm density of new shoot recruits was lowest in the month of December in all the sites with Wasare and Singida sites having an average of one each while Ogenya had 2 ± 0.3 culms/m2. However, variations was not observed among transects.
Papyrus culm mortality was monitored throughout the study period. Senescing culms were observed to be higher in Wasare in the month of November at an average of 12.5 ± 4.5 culms/m2 (X2 = 8.24; df = 2; P < 0.05). However, Ogenya had the highest number of dead culms in September (3.7 ± 0.4 culms/m2) while Singida (6.0 ± 3.2 dead culms/m2) was high in October although there was no difference between Ogenya and Singida in period 1 (Figure 5). Furthermore, no difference in number of dead culms observed among months of measurements and between Singida and Ogenya in period 2.
Several factors were identified for the cause of culm mortality in Nyando wetland. Natural causes of culm mortality dominated in all the three sites with Ogenya (71%), Singida (60%) and Wasare (59%) as depicted in Figure 6. Culms that died as a result of cutting accounted for 24% in Wasare while it was 3% in Ogenya and none was observed in Singida. Mortality associated with insects/rodents accounted for 40% in Singida while Ogenya and Wasare had 10% and 14%, respectively. Herbivory related mortality due to grazing by livestock, Sitatunga and probably Hippos accounted for 16% in Ogenya and 3% in Wasare while this type of mortality was not observed in Singida.

Table 1. Physical and chemical soil characteristics from three sites of Nyando wetland. Data was from the months of June, July, September and November 2010. The values are means ± standard error (n = 4) except for bulk density (n = 2). Ranges are given in brackets.

Figure 5. Monthly variations in culm density, dead and new shoots in the three study sites of Nyando wetland for period 1 and 2 in Singida and Ogenya. Data for Wasare site depicts monthly results obtained only for period 1. Values are means ± standard error (SE) (n = 3).

Figure 6. Percentage of culm mortality caused by natural dead (N), cut (C), insect/rodent (IR), and grazed (G) in Ogenya, Singida and Wasare sites of Nyando wetland. Data is the total number of dead culms per mortality cause per site combining period 1 and 2 including the average for all the sites.
Overall, average culm mortality caused by natural senescing was 64% while insect/rodent infestation accounted for 19% mortality. In addition, grazing and cutting accounted for 7% and 10% mortality, respectively (Figure 6). The density of culms that survived by the end of period 1 were 5 ± 0.9, 3 ± 1.5, and 4.5 ± 0.5 culms/m2 in Ogenya, Singida and Wasare, respectively. These culms were below 200 cm in length at the start of measurements and can be categorized between growth stage I and II (closed and just opening) as described by [22] .
The effect of surface water depth on density, number of new shoots and number of dead culms was explored. According to Spearman's coefficient of correlation, water depth did not have a significant influence on culm density and dying culms in all the sites. However, water depth had a positive and significant relationship with the number of new shoot recruits in Wasare site (rho = 0.80; P = 0.01) and not in Ogenya (rho = 0.34; P > 0.05) and Singida (rho = 0.48; P > 0.05). Furthermore, it was noted that culm density had an effect on the number of senesced (rho = 0.89; P < 0.001) and new shoot recruits (rho = 0.77; P = 0.01) in Ogenya and not in Singida and Wasare.
3.3. Papyrus Growth Rate and Relationship between Culm Length and Culm Girth
Papyrus growth rate is a valuable parameter that is related to nutrient regulation process with optimal hydrological regime. The growth rate of papyrus culm differed between the study sites and period of study. Singida had a higher average absolute culm length growth rate of 2.5 ± 0.2 cm/day and 2.4 ± 0.2 cm/day in period 1 and 2, respectively. This was followed by Ogenya with 1.5 ± 0.2 and 2.0 ± 0.2 cm/day in period 1 and 2, respectively and lowest in Wasare 1.4 ± 0.1 cm/day period 1. Figure 7 shows relative growth rate (
Likewise, culm girth growth rate was observed to be different in Singida as compared to Wasare and Ogenya. Girth relative growth rate was higher in period 2 than in period 1 (KW-ANOVA, P < 0.05; Figures 8(a)-(e). There was a significant difference among transects and between periods (KW-ANOVA, P < 0.05) but no interaction effect was observed between transect and period of measurements (KW-ANOVA, P > 0.05; Figure 8).
Linear regression model was used to determine the relationship between culm length and culm girth using log-transformed data (Table 2). A strong culm length-girth relationship was observed for Singida period 1 (R2 = 0.64; P < 0.001) followed by Ogenya (R2 = 0.50; P < 0.001) and Wasare (R2 = 0.34; P < 0.001). The relationship in period 2 was weak although significant for Singida (R2 = 0.15; P < 0.001) and not in Ogenya.
3.4. The Effect of Water Quality and Soil Type on Papyrus Growth Rate
Papyrus culms growing in wetland quadrats are affected by different factors. Average culm size, density, water quality and soil characteristics have relationship with growth rate of culms in quadrats. Correlation matrices between 21 variables showed significant correlations between total culm density with new and dead culms; water depth and EC; pH and
Table 3 shows three multiple linear regression models for culm length period 1 and 2. All models were significant (P < 0.05). The highest correlation coefficient between predictor variables for culm length in model 1 was 0.414 (between SRP and water depth) which explains a variance of 17% in one variable hence, can be included as the variance is low and can be tolerated. In model one, relative growth rate (>200 cm) in period 1 against average culm length, total culm density, water depth, NH4, and site as dummy showed that growth was significantly and negatively affected by average length, total density and NH4. On the other hand, growth was significantly and positively affected by surface water depth and site. This was also the case with culms < 200 cm length (model 2) for period 1. However, growth rate of culms < 200 cm for period 2 was additionally influenced positively but not significant by SRP and not dummy site. In model three (period 2), the highest correlation coefficient between predictor variables was 0.426 (between average height and number of dead culms) which explains 18% and

Figure 7. Relative growth rate of C. papyrus L. in relation to average culm length in (a) Ogenya P1 (b) Ogenya P2 (c) Singida P1 (d) Singida P2 and (e) Wasare P1 showing fitted linear regression of growth rate versus maximum culm length. P1 and P2 stands for period 1 and 2.

Figure 8. Relative growth rate of C. papyrus L. in relation to average culm girth in (a) Ogenya P1 (b) Ogenya P2 (c) Singida P1 (d) Singida P2 and (e) Wasare P1 showing fitted linear regression of growth rate versus maximum culm girth. P1 and P2 stands for period 1 and 2.
Table 2. Linear regression coefficient relationship between papyrus culm girth and culm length at three study sites of Nyando wetland in period 1 and 2 showing the intercept, slope and adjusted R2 for log-transformed data.
+Wasare period 2 was not measured as the site had been previously burned; *Significant at 5%; **Significant at 1%
Table 3. Multiple regression models for growth of papyrus culms in two categories of growth stages relating with water quality and soil parameters as predictor variables and length relative growth rate as dependent variable. The significance of partial regression coefficient (b) are shown by stars (***: 0.1%, **: 1%, *: 5%), and s.e. is the standard error of the bs while beta shows the beta-weights for variables in the model.
+Represents data for period 2 with culm length below 200 cm.
can be tolerated. Relative growth rate against average height, number of new culms, and TP presented average height as significant and negatively affecting growth. Also number of new culms and TP significantly and positively affected growth rate of culm length (R2 = 0.27; P < 0.001; Table 3).
Durbin-Watson statistics showed positive serial correlations in two models of culm height. For example, model 1 and 3, the Durbin-Watson statistics was 1.7876 and 1.7710, respectively while for model 2 was 2.17. All models are greater than the significance level dL = 1.49 from the Durbin-Watson table [24] .
4. Discussion
Papyrus growth relies on wetland water depth, water quality and soil characteristics. Variation in water depth is a common feature in tropical papyrus wetlands which is influenced by seasonality [2] [7] . According to [2] , there is a link between hydrological regime and livelihood exploitation activities. The three sites studied in Nyando represent different hydrological regimes which influence the kind of livelihood activities that were observed in the wetland
Water depth differed among the three sites that led to the variations of activities and papyrus productivity observed in Nyando. High water depth in Singida was characterized by low culm density, low biomass, but thick and tall culms. However, Ogenya with water throughout the study period was characterized by moderate culm density, low growth rate, high biomass, but thin and moderately tall culms. Wasare site was dry for at least six months during the study period and was characterized by low growth rate, high culm density, moderate biomass and moderate thick and tall culms. Although biomass and culm density varied among sites in Nyando, they can be compared with other papyrus-dominated wetlands in East Africa (Table 4). Other researchers (e.g. [25] [26] [27] have shown that increase in water depth can lead to increase in shoot length and a decrease in shoot density. Their results are comparable to what was observed for papyrus in less disturbed site, Singida. Likewise, some studies have shown that shoot density is higher in drier than in wet areas [28] [29] which agrees with our results of the disturbed area in Wasare.
Water depth and ecosystem characteristics can influence papyrus production (growth rate, new shoots, density) including mortality. Recruitment of new shoots, culm density, senescing and mortality of papyrus shoots was observed to be a continuous process in Nyando wetland. More so, mortality was observed to occur in all the sites in the month of November (Figure 5). The reason could be due to water level that dropped, low pH, SRP and TP in surface water, while NO2, NO3, NH4 and TN were highest in November. According to [30] and [31] , papyrus growth requires a pH range of 6 - 8.5. However, in this study, the pH in surface water was below 6 in Wasare and Singida in November while the soil pH was below 6 in all the sites which may have caused culms to die.
Productivity of papyrus in natural ecosystems varies in density and growth rate due to environmental conditions prevailing at the site [32] . Due to disturbances such as cutting, grazing, insect/rodent infestation and hydrological modification, mortality rate may increase to synergize the natural senescing. Some studies have shown that harvesting and burning increase reed density and the amount of dead shoots while it decreases relative growth rate, shoot length and diameter [33] [34] . More research is required to isolate factors causing mortality of papyrus and to understand whether they support each other in affecting papyrus productivity in the face of climate uncertainty.
Table 4. Comparison of Cyperus papyrus L. mean (±SE) culm density and aerial dry biomass at different study sites in East Africa.
*This site was relatively undisturbed by human activities. +This site was dry for a longer period and was accessible to human exploitation activities.
Papyrus productivity depends on the availability of nutrients and the habitat conditions which enables faster rate of nutrient uptake [13] . However, this changes when the conditions are poor. As papyrus is harvested and hydrological conditions are altered, provisioning and regulation services diminish. Since flood water brings nutrients and deposit sediments in the wetland, it is likely that the wetland nutrients are seasonally restored so as to maintain continuous growth. More nutrients are released to the wetland as the various plants of the wetland die and decompose [35] [36] . These nutrients are made available mainly during the wet season to be taken up by plants. [37] showed that for plants to absorb nutrients, wet conditions are required. This meant that nutrients were availed to plants in high water levels hence high growth rate that was observed in Singida. In dry season in Wasare, nutrients became limiting in the substrate as the site was disturbed through channelization, burning, grazed by livestock and converted to crop lands. These activities accelerated the drying that formed cracks in dry season which resulted in destabilizing the substrate and making nutrients unavailable for plants to take-up. This has been observed in Lake Chilwa where growth of potential plants was inhibited due to drying [38] .
The relative growth rate of papyrus in Nyando wetland did not differ among the sites but at different periods. Period 1 had an average of 0.06% while it increased to 0.14% in period 2. These results were slightly higher as compared to papyrus growth rate constant of 0.04/day growing in low strength domestic wastewater in Thailand [39] . The growth rate and the concentration of nutrients in the plant tissue may limit the potential rate of nutrient uptake [40] . In addition, [41] listed rapid growth, high nutrient content in plant tissue and capability to attain high standing crop yield as characteristics of a good plant to be used for nutrient assimilation and storage. However, these characteristics depend on the prevailing conditions and the site status in terms of disturbances and hydrological regime patterns. Papyrus high productivity enables it to have potential to remove nutrients in wetlands receiving more loads of nutrients [42] [15] . Papyrus plants in Wasare site had a brief connection with flowing water that lasted 2 - 3 weeks and therefore may not buffer the lake from nutrient loads. However, Singida natural undisturbed vegetation may buffer the lake from receiving nutrients by incorporating nutrients in plant tissues.
Multiple regression models showed that water depth influenced growth rate positively for both mature and young plants. However, SRP was an important variable for young plants (model 2) than for mature plants (model 1). The influence of SRP has been seen in other wetland plants (Phragmites sp.) which showed that increased nutrients led to increase in length and diameter [43] . For mature plants (model 1), site condition influenced papyrus growth rate positively since there was no disturbance, had stable water levels and nutrients. Likewise, NH4, culm density and mean culm length influenced negatively the growth rate in model 1 and 2. Although [44] found out that nitrogen can increase leaf area, a decrease in plant growth due to NH4 in papyrus-dominated wetland was observed. This agrees with results of [45] who reported a decrease in the area of individual leaves with decreasing N fertility. This decrease in length and girth may be attributed to reduction in cell expansion which was proposed by [46] . There is need for more research to understand why there is stunted growth with high N fertility in the papyrus wetland.
The regression models used to explain factors that influence growth rate of papyrus was low (R2 = 37%) but was significant. This may be explained by long intervals (monthly) that was used to monitor papyrus growth. This resulted in obtaining zero growth of some culms in young and mature plants alike. Since plant growth is continuous and physical and chemical characteristics of water and soil frequently change, there may be loss of information while averaging length and environmental parameters in a month. Therefore, to improve on this study, there is need to have daily measurements of growth rate of papyrus under certain conditions. Also all plants monitored for growth rate need to start at the initial growth stage and be monitored at close intervals to capture local changes within the study sites.
Clearly, there are distinct seasonal changes in Nyando wetland which may occasionally be prolonged to have negative or positive effects on the growth of papyrus. Water level fluctuation is the main driving factor of wetland productivity in papyrus-dominated wetland. High water levels will maintain the growth of papyrus and allow other processes such as decomposition, nutrient uptake, and habitat provisioning for other organisms to occur. Low water level on the other hand will lead to stunted papyrus growth, less nutrient availability and easy accessibility for grazing livestock, harvesting of biomass, burning, and conversion to agricultural crops. When these factors are coupled with natural water fluctuations in the wetland, it will impact negatively on the growth of papyrus and may lead to stressed ecosystem that threatens its integrity.
This study used exploratory data analysis using multiple linear regression technique. This technique is important as it analyzes and summarizes large datasets which would otherwise be impossible to obtain. Though this technique resulted in low explanation of factors influencing growth of papyrus, the models can stimulate thinking about the papyrus-dominated system being studied. The technique has suggested various relationships that occur among different variables that may not have been established. This has opened another area to be studied in terms of setting and designing experiments to understand the various theories that the models have shown relating to papyrus. This implies that the information obtained can assist in early warning signs of a stressed ecosystem so as to plan for response activities that will curb the problem before it goes out of hand. In addition, variation among the study sites is a good way of localizing effects that affect plant growth. This will inform the decision-makers and ecosystem managers on where to start the remedy early.
5. Conclusion and Recommendations
Papyrus-dominated wetland in Nyando exhibit varying hydrological regimes that influence livelihood activities affecting papyrus growth dynamics. These conditions are important in determining the growth attributes of C. papyrus and for maintaining ecosystem healthy. Surface water depth in the wetland had a significant relationship with papyrus growth rate which was supported by the site condition. Mortality of culms in areas with low human activities was attributed to natural senescing which may have been influenced by changes in water levels. However, it is important to note that insects/rodents were responsible too for mortality of culms. Higher papyrus growth can be achieved in areas with stable water regimes, undisturbed substrate, and availability of nutrients combined with minimal disturbances from human activities at the site. Low growth rate of papyrus was observed in disturbed floodplain of Wasare which was characterized by high culm density and stunted growth. Estimated length-girth linear regression relationship could be used to determine the maturity of papyrus vegetation in areas with low human interference. Based on multiple regression models, water depth, SRP and TP positively influenced relative growth rate while culm density, mean length and NH4 negatively influenced papyrus relative growth rate.
Although wetland environmental conditions explained little in papyrus growth, other factors that play a role in influencing papyrus growth need to be explored. Factors speculated to have influenced growth of papyrus and need to be explored may include among others; interstitial water; substrate characteristic; water exchange between wetland and lake, and type of human activities in the wetland. Natural wetlands are characterized by high variability leaving an option for observational studies rather than controlled experiments. In Nyando wetland, human activities and seasonal events complicate experimental studies. Therefore, controlled experimental quadrats could be carried out in intact wetland and those that have been cleared to understand the effect of specific nutrients and human disturbances on growth indicators. For sustainable management of papyrus-dominated wetlands, there is need to understand the differences among the sites as climatic change effects are experienced over a long-term period.
Acknowledgements
This work was conducted in collaboration with the Departments of Water Science and Engineering Technology of UNESCO-IHE Institute for Water Education, Delft, Biological Sciences of Egerton University, Kenya and Victoria Institute for Environment and Development (VIRED International), Kenya. This project was funded by DGIS through UNESCO-IHE Partnership Research Fund (UPaRF) to “Ecology of Livelihoods in East African Papyrus Swamps” (ECOLIVE) project. Thanks to Joel Onyango, research assistant, MSc. students Patteson Chula and Patrick Orwa who assisted in the field work. We appreciate the community members at the study sites for allowing access and participating in the project activities.
Cite this paper
Rongoei, P.J.K. and Outa, N.O. (2016) Cyperus papyrus L. Growth Rate and Mortality in Relation to Water Quantity, Quality and Soil Characteristics in Nyando Floodplain Wetland, Kenya. Open Journal of Ecology, 6, 714-735. http://dx.doi.org/10.4236/oje.2016.612065
References
- 1. Kipkemboi, J., van Dam, A.A. and Denny, P. (2006) Biophysical Suitability of Smallholder Integrated Aquaculture-Agriculture Systems (Fingerponds) in East Africa’s Lake Victoria Freshwater Wetlands. International Journal of Ecology and Environmental Science, 32, 75-83.
- 2. Rongoei, P.J.K., Kipkemboi, J., Okeyo-Owuor, J.B. and van Dam, A.A. (2013) Ecosystem Services and Drivers of Change in Nyando Floodplain Wetland, Kenya. African Journal of Environmental Science and Technology, 7, 274-291.
- 3. Kinaro, Z.O. (2008) Wetland Conversion to Large-Scale Agricultural Production. Implications on the Livelihoods of Rural Communities, Yalla Swamp, Lake Victoria Basin, Department of Water and Environmental Studies, University of Linköping, Linköping.
- 4. Rongoei, P.J.K., Kipkemboi, J., Kariuki, S.T. and van Dam, A.A. (2014) Effects of Water Depth and Livelihood Activities on Plant Species Composition and Diversity in Nyando Floodplain Wetland, Kenya. Wetlands Ecology and Management, 22, 177-189.
http://dx.doi.org/10.1007/s11273-013-9313-7 - 5. Denny, P. (1985) Wetland Vegetation and Associated Plant Life Forms. In: Denny, P., Ed., The Ecology and Management of African Wetland Vegetation, Dr W. Junk Publishers, Dordrecht, 1-18.
http://dx.doi.org/10.1007/978-94-009-5504-2_1 - 6. Azza, N.G.T., Kansiime, F., Nalubega, M. and Denny, P. (2000) Differential Permeability of Papyrus and Miscanthidium Root Mats in Nakivubo Swamp, Uganda. Aquatic Botany, 67, 169-178.
http://dx.doi.org/10.1016/S0304-3770(00)00093-0 - 7. Boar, R.R. (2006) Responses of a Fringing Cyperus papyrus L. Swamp to Changes in Water Level. Aquatic Botany, 84, 85-92.
http://dx.doi.org/10.1016/j.aquabot.2005.07.008 - 8. Kansiime, F., Saunders, M. and Loiselle, S. (2007) Functioning and Dynamics of Wetland Vegetation of Lake Victoria: An Overview. Wetlands Ecology and Management, 15, 443-451.
http://dx.doi.org/10.1007/s11273-007-9043-9 - 9. Gaudet, J.J. (1977) Natural Drawdown on Lake Naivasha, Kenya and the Formation of Papyrus Swamps. Aquatic Botany, 3, 1-47.
http://dx.doi.org/10.1016/0304-3770(77)90002-X - 10. Mnaya, B., Asaeda, T., Kiwango, Y. and Ayubu, E. (2007) Primary Production in Papyrus (Cyperus papyrus L.) of Ru-Bondo Island, Lake Victoria, Tanzania. Wetlands Ecology and Management, 15, 269-275.
http://dx.doi.org/10.1007/s11273-006-9027-1 - 11. Terer, T., Triest, L. and Muthama, M.A. (2012) Effects of Harvesting Cyperus papyrus in Undisturbed Wetland, Lake Naivasha, Kenya. Hydrobiology, 680, 135-148.
http://dx.doi.org/10.1007/s10750-011-0910-2 - 12. Osumba, J.J.L., Okeyo-Owuor, J.B. and Raburu, P.O. (2010) Effect of Harvesting on Temporal Papyrus (Cyperus papyrus) Biomass Regeneration Potential among Swamps in Winam Gulf Wetlands of Lake Victoria Basin, Kenya. Wetlands Ecology Management, 18, 333-341.
http://dx.doi.org/10.1007/s11273-010-9174-2 - 13. Thompson, K., Shewry, P.R. and Woolhouse, H.W. (1979) Papyrus Swamp Development in the Upemba Basin, Zaire: Studies of Population Structure in Cyperus papyrus Stands. Botanical Journal of the Linnean Society, 78, 299-316.
http://dx.doi.org/10.1111/j.1095-8339.1979.tb02573.x - 14. Muthuri, F.M., Jones, M.B. and Imbamba, S.K. (1989) Primary Productivity of Papyrus (Cyperus papyrus) in a Tropical Swamp: Lake Naivasha, Biomass, 18, 1-14.
http://dx.doi.org/10.1016/0144-4565(89)90077-2 - 15. Kyambadde, J., Kansiime, F. and Dalhammar, G. (2005) Nitrogen and Phosphorus Removal in Substrate-Free Pilot Constructed Wetlands with Horizontal Surface Flow in Uganda. Water, Air and Soil Pollution, 165, 37-59.
http://dx.doi.org/10.1007/s11270-005-4643-6 - 16. Owino, A.O. and Oyugi, J.O. (2008) Some Conservation Aspects of Papyrus Endemic Passerines around Lake Victoria, Kenya. Scopus, 28, 25-30.
- 17. Mwakubo, S.M. and Obare, G.A. (2009) Vulnerability, Livelihood Assets and Institutional Dynamics in the Management of Wetlands in Lake Victoria Watershed Basin. Wetlands Ecology Management, 17, 613-626.
http://dx.doi.org/10.1007/s11273-009-9138-6 - 18. World Agroforestry Centre (2006) Improved Land Management in the Lake Victoria Basin: Final Report on the TransVic Project. World Agroforestry Centre, Nairobi.
- 19. APHA (1998) Standard Methods for the Examination of Water and Wastewater. 20th Edition, American Public Health Association, American Water Works Association and Water Environmental Federation, Washington DC.
- 20. Okalebo, J.R., Gathua, K.W. and Woomer, P.L. (2002) Laboratory Methods of Soil and Plant Analysis: A Working Manual. 2nd Edition, SACRED Africa, Nairobi, 128 p.
- 21. Gee, G.W. and Bauder, J.W. (1986) Particle Size Analysis. In: Klute, A., Ed., Methods of Soil Analysis, Part 1: Physical and Minerological Methods, 2nd Edition, American Society of Agronomy, Madison, 383-408.
- 22. Muthuri, F.M. and Jones, M.B. (1997) Nutrient Distribution in a Papyrus Swamp: Lake Naivasha, Kenya. Aquatic Botany, 56, 35-50.
http://dx.doi.org/10.1016/S0304-3770(96)01093-5 - 23. R Core Team (2014) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
http://www.R-Project.org/ - 24. Yamane, T. (1973) Statistics: An Introductory Analysis. 3rd Edition, Harper and Row, New York.
- 25. Edwards, A.L., Lee, D.W. and Richards, J.H. (2003) Responses to a Fluctuating Environment: Effects of Water Depth on Growth and Biomass Allocation in Eleocharis cellulose Torr (Cyperaceae). Canadian Journal of Botany, 81, 964-975.
http://dx.doi.org/10.1139/b03-091 - 26. Eid, E.M., Shaltout, K.H., Al-Sodany, Y.M. and Jensen, K. (2010) Effects of Abiotic Conditions on Phragmites australis along Geographical Gradients in Lake Burullus, Egypt. Aquatic Botany, 92, 86-92.
http://dx.doi.org/10.1016/j.aquabot.2009.10.010 - 27. Webb, J.A., Wallis, E.M. and Stewardson, M.J. (2012) A Systematic Review of Published Evidence Linking Wetland Plants to Water Regime Components. Aquatic Botany, 103, 1-14. http://dx.doi.org/10.1016/j.aquabot.2012.06.003
- 28. Bodensteiner, L.R. and Gabriel, A.O. (2003) Response of Mid-Water Common Reed Stands to Water Level Variation and Winter Conditions in Lake Poygan, Wisconsin, USA. Aquatic Botany, 76, 49-64.
http://dx.doi.org/10.1016/S0304-3770(03)00013-5 - 29. Engloner, A.I. (2004) Annual Growth Dynamics and Morphological Differences of Reed (Phragmites australis [Cov.] Trin. ex Steudel) in Relation to Water Supply. Flora, 199, 256-262.
http://dx.doi.org/10.1078/0367-2530-00153 - 30. Duke, J.A. (1983) Cyperus papyrus L. In: Duke, J.A., Ed., Handbook of Energy Crops, Purdue University, West Lafayette, E-Book.
- 31. Serag, M.S. (2003) Ecology and Biomass Production of Cyperus papyrus L. on the Nile Bank at Damietta, Egypt. Journal of Mediterranean Ecology, 4, 15-24.
- 32. Okurut, T.O. (2000) A Pilot Study on Municipal Wastewater Treatment Using a Constructed Wetland in Uganda. PhD Dissertation, UNESCO-IHE, Institute of Water Education, Delft.
- 33. Ostendorp, W. (1999) Management Impacts on Stand Structure of Lakeshore Phragmites Reeds. International Review of Hydrobiology, 84, 33-47.
- 34. Warren, R.S., Feel, P.E., Grimsby, J.L., Buck, E.L., Rilling, G.C. and Fertik, R.A. (2001) Rates, Patterns and Impacts of Phragmites australis Expansion and Effects of Experimental Phragmites Control on Vegetation, Macroinvertebrates and Fish within Tidelands of the Lower Connecticut River. Estuaries, 24, 90-107.
http://dx.doi.org/10.2307/1352816 - 35. Pratima, S., Asaeda, T., Manatunge, J. and Fujino, T. (2006) Nutrient Cycling in Natural Stand of Typha angustifolia. Journal of Freshwater Ecology, 3, 431-438.
- 36. Rongoei, P.J.K., Kipkemboi, J. and Van Dam, A.A. (2015) Decomposition and Nutrient Dynamics of Plant Litter in Nyando Floodplain Wetland, Kenya. Egerton Journal of Science and Technology, 15, 163-186.
- 37. Dodds, W. and Whiles, M. (2010) Freshwater Ecology: Concepts and Environmental Applications of Limnology. 2nd Edition, Elsevier, Amsterdam, 330-333.
- 38. Osborne, P.L. (2012) Tropical Ecosystems and Ecological Concepts. Cambridge University Press, Cambridge, 349-353.
http://dx.doi.org/10.1017/cbo9781139057868 - 39. Perbangkhem, T. and Polprasert, C. (2010) Biomass Production of Papyrus (Cyperus papyrus) in Constructed Wetland Treating Low-Strength Domestic Wastewater. Bioresource and Technology, 101, 833-835.
http://dx.doi.org/10.1016/j.biortech.2009.08.062 - 40. Vyamazal, J. (2007) Removal of Nutrients in Various Types of Constructed Wetlands. Science of the Total Environment, 380, 48-65.
http://dx.doi.org/10.1016/j.scitotenv.2006.09.014 - 41. Reddy, K.R. and DeBusk, W.F. (1987) Nutrient Storage Capabilities of Aquatic and Wetland Plants. In: Reddy, K.R. and Smith, W.H., Eds., Aquatic Plants for Water Treatment and Resource Recovery, Magnolia Publishing, Orlando, 337-357.
- 42. Kansiime, F. and Nalubega, M. (1999) Wastewater Treatment by a Natural Wetland: The Nakivubo Swamp Uganda-Processes and Implications. PhD Thesis, UNESCO-IHE, Delft.
- 43. Lovelock, C.E., Ball, M.C., Martin, K.C. and Feller, I.C. (2009) Nutrient Enrichment Increases Mortality of Mangroves. PLoS ONE, 4, e5600.
http://dx.doi.org/10.1371/journal.pone.0005600 - 44. Oberbauer, S.F., Hastings, S.J., Beyers, J.L. and Oechel, W.C. (2006) Comparative Effects of Downslope Water and Nutrient Movement on Plant Nutrition, Photosynthesis and Growth in Alaskan Tundra. Ecography, 12, 324-334.
http://dx.doi.org/10.1111/j.1600-0587.1989.tb00853.x - 45. Vos, J. and van de Putten, P.E.L. (1998) Effect of Nitrogen Supply on Leaf Growth, Leaf Nitrogen Economy and Photo-Synthetic Capacity of Potato. Field Crops Resource, 59, 63-72.
http://dx.doi.org/10.1016/S0378-4290(98)00107-5 - 46. Del Amor, F.M. (2006) Growth, Photosynthesis and Chlorophyll Fluorescence of Sweet Pepper Plants as Affected by the Cultivation Method. Annals of Applied Biology, 148, 133-139.
http://dx.doi.org/10.1111/j.1744-7348.2006.00048.x