 Advances in Anthropology 2012. Vol.2, No.2, 87-105 Published Online May 2012 in SciRes (http://www.SciRP.org/journal/aa) http://dx.doi.org/10.4236/aa.2012.22010 Copyright © 2012 SciRes. 87 Ancient History of the Arbins, Bearers of Haplogroup R1b, from Central Asia to Europe, 16,000 to 1500 Years before Present Anatole A. Klyosov The Academy of DNA Genealogy, Newton, USA Email: aklyosov@comcast.net Received March 14th, 2012; revised April 13th, 2012; accepted April 23rd, 2012 This article aims at reconstructing the history of R1b ancient migrations between 16,000 and 1500 years before present (ybp). Four thousand four hundred eight (4408) haplotypes of haplogroup R1b (with sub- clades) were considered in terms of base (ancestral) haplotypes of R1b populations and the calculated time to their common ancestors. The regions considered are from South Siberia/Central Asia in the east (where R1b haplogroup arose ~16,000 ybp) via the North Kazakhstan, South Ural to the Russian Plain and further west to Europe (the northern route entering Europe around 4500 ybp); from the Russian Plain south to the Caucasus (6000 ybp), Asia Minor (6000 ybp) and the Middle East (6000 - 5500 ybp) to the Balkans in Europe (the southern route, entering Europe around 4500 ybp); along North Africa and the Mediterranean Sea (5500 - 5000 ybp) via Egypt to the Atlantic, north to Iberia (the North African route with arrival to the Pyrenees 4800 ybp). The Arbins (bearers of R1b haplogroup) along their migration route to the Middle East and South Mesopotamia apparently have established the Sumer culture (and the state), moving westward to Europe (5000 - 4500 ybp) carrying mainly the R-M269 subclade and its downstream L23 subclade. This last subclade was nearly absent along the North African route, and/or did not survive the migration to Iberia or evidenced later. At the arrival to Iberia (4800 ybp) the M269 sub- clade split off M51 and soon thereafter the L11 downstream subclades. These populations became known as the Bell Beakers and moved north, along with the newly arisen subclades of P312 and L21 (which split off within a few centuries after P312). Those subclades and their downstream clades have effectively, without major interruptions, populated Europe (the smooth haplotype trees demonstrate the near non-stop proliferation of R1b haplotypes in Europe). They are evidenced from the Atlantic eastward to the Balkans, Carpathian Mountains, present day Poland to the western border of the Russian Plain and up to the Baltic Sea. The Isles had a different history of R1b migrations. The bearers of L11, P312 and L21 moved to the Isles by land and sea concurrently with those Arbins who were populating Europe between 4000 and 2500 ybp and formed the respective “local” subclades of P314, M222, L226, which largely populated the Isles. As a result, a significant part of the Isles is populated almost exclusively by the Arbins, whose frequency reaches 85% - 95% among the current population. In general, the frequency of Arbins in Western and Central Europe, reaches—albeit not uniformly—some 60% of the population. This study essentially pre- sents an example of application of DNA genealogy in studying the history of mankind. Keywords: Y Chromosome; Mutations; Haplotypes; Haplogroups; TMRCA; STR; SNP; Arbins; Aryans; R1b Introduction The origin and history of haplogroup R1b, bearers I refer to as the Arbins (see explanation below), currently populate nearly 60% of Western and Central Europe. Additionally Arbins populate significant parts of the Caucasus, Anatolia and Asia Minor, Middle East, and many locations in Central Asia, in- cluding South Siberia, Altay, Tuva, North-Western China, Middle Asia, some Ural and Middle Volga regions with ethnic groups and populations such as the Bashkirs, Tatars, Chuvash, and other. History of R1b was significantly distorted from the begin- ning of “genetic genealogy” at the end of 1990-s, when it was claimed, quite groundlessly, that R1b arose in Europe some 30,000 years before present (ybp). Groundlessly because, in- deed, the claims were based on no data. Such data never existed. Nevertheless, statements and claims such as “Around 30,000 years ago, a descendant of the clan making its way into Europe gave rise to marker M343, the defining marker of haplogroup R1b. These travelers are direct descendants of the people who dominated the human expansion into Europe, the Cro-Magnon” (Spencer Wells, “Deep Ancestry”, 2006). This and similar claims, such as R1b (and its M269 subclades) were “well established throughout Paleolithic Europe”, “contemporaneous with Aurig- nacian culture”, “the earliest expansion into Europe, during the Upper Paleolithic ~30,000 years ago” by Wells, Semino, Un- derhill, Cavalli-Sforza, Cinnioglu, Kivisild, Wiik and many others (e.g., Semino et al., 2000; Wells et al., 2001; Cinnioglu et al., 2004; Wiik, 2008) were essentially based on “thoughts” that if people lived in Europe some 30,000 years ago, they nec- essarily were of the R1b haplogroup, and not of I, G, J, E, F or any other haplogroups. Were any haplotypes analyzed? Their mutations counted? Any chronological evaluations? There was nothing of the above. “Paleolithic origin” of R1b in Europe, or their “Paleolithic migrations” to Europe around 10,000 - 8000 ybp are still claimed 87  A. A. KLYOSOV in recent academic papers, such as (Myres et al., 2010; Balares- que et al., 2010; Morelli et al., 2010). These assumptions and resulting calculations are based, typically, on “population muta- tion rates” (Zhivotovsky et al., 2004; Hammer et al., 2009; Underhill et al., 2009), which commonly exaggerate the chro- nological estimates of migrations and events by 200% - 400% since they are based on crude, artificial and unrealistically naïve and generalized reasoning (for critique, see Klyosov, 2009a, 2009b, 2009c; Rozhanskii & Klyosov, 2011). The “academic papers”, placing origin of R1b in Asia Minor or nearby, have not considered regions east of Asia Minor as well as R1b hap- lotypes of those eastern regions. In short, the whole story of R1b migrations and their history is in disarray, and the “popu- lation geneticists” continue to advance misleading conclusions due to their methodology. This study’s methodology/analysis includes considerations of Y chromosome extended 67 and 111 marker haplotypes, when available. The methodology was described in detail in the pre- ceding papers in this journal (Rozhanskii & Klyosov, 2011; Klyosov & Rozhanskii, 2012a) and elsewhere (e.g., Klyosov, 2009a, 2009c, 2009d), and in Materials and Methods section of this article. As described (Klyosov & Rozhanskii, 2012b), Europeoids (Caucasoids) appeared ~58,000 ybp. They gradually branched to downstream haplogroups and their subclades, and migrated to the north, west, south and east. Haplogroup NOP, which was among them, arose ~48,000 ybp, and moved eastward, pre- sumably towards South Siberia and/or adjacent regions. Hap- logroup P split off ~38,000 ybp, presumably in South Siberia, and gave rise to haplogroup R and then R1 ~30,000 - 26,000 ybp (see the diagram in Klyosov and Rozhanskii, 2012b). Hap- logroup R1b arose ~16,000 ybp, as it will be shown further in this paper. The timing may be reconstructed from a series of R1b hap- lotypes, made available from the databases (see Appendix). The most distant R1b haplotypes (those exhibiting the greatest mu- tational differences) from European R1b haplotypes, were found in Siberia and Middle Asia (a part of Central Asia) populations. Central Asian R1b haplotype bearers have the most ancient common ancestors with European R1b bearers, and those ancient common ancestors lived ~16,000 ybp in Cen- tral Asia. We do not know as yet whether in South Siberia or Middle Asia; however, the evidence will demonstrate that it was somewhere in that vast region. In this endeavor both terms, “haplogroup” and “subclade”, are employed as near equivalents because all haplogroups are essentially subclades of other upstream haplogroups, and usage of one or the other of these terms is suggested by the context. This is shown in the following diagram (ISOGG-2012, a frag- ment, http://www.isogg.org/tree/ISOGG_HapgrpR.html) which is related to the most ancient subclades of R1b haplogroup: R1b M343 R1b1 L278, M415, P25_1, P25_2, P25_3 R1b1* - R1b1a P297, L320 R1b1a1 M73, M478 R1b1a2 L265, M269, M520, S3, S10 R1b1b M335 R1b1c V88 R1b1c* - R1b1c1 M18 R1b1c4 V69 Due to lack of a common name for bearers of R1b hap- logroup (with subclades) and their languages in ancient times, which were carried for millennia and eventually brought to Europe as non-Indo European languages, I refer to them as the Arbins (from R1b), both the people and their original languages, on the analogy with the Aryans, who essentially belonged to haplogroup R1a (Klyosov & Rozhanskii, 2012a). A Bird’s-Eye View at R1b Haplotypes, and the Most Ancient R1b Populations Figure 1 presenting an overview of an R1b haplotype tree includes 338 haplotypes in a short 8 marker format. The pur- pose of the presentation is to show the complex pattern of the R1b haplogroup and identify the most ancient branch on the tree. The haplotype tree as shown next is arranged by a computer program which combines branches based on the similarity of their alleles in the respective markers (or loci) in Y chromo- some, and dynamics of their alleles (Klyosov, 2009c and refer- ences therein). Haplotypes identical to each other and prevail- Figure 1. The 8-marker 338 haplotype tree for R1b1 haplotypes, composed from data listed in (Zhong et al., 2010). The tree includes haplotypes of sub- clade R1b* (8 haplotypes), R1b1* (2 haplotypes), R1b1c (2 haplotypes), R1b (140 haplotypes), R1b1a1 (12 haplotypes), R1b2b (21 haplotypes), R1b1a2 (153 haplotypes). Regarding ethnicity and/or location, the haplotypes belong to 40 Western Europeans, 17 Iberians, 84 Turks, 48 Albanians, Macedonians and Romanians, 40 Italians, 30 Cretans, 13 Greeks and Macedonians, 9 Romanians from a different region, 13 Pakistanis, 4 Indians, 2 Japanese, 2 Mongolians (one of them, No. 176, belongs to R1b1b1-M73 subclade and stands out in the lower part of the tree), 2 Tibetans, 1 Kyrgyz from Xinjiang, 7 Han Chinese, 16 Uighur (12 from the Xinjiang Province, six of them belong to R1b* subclade, three R1b1b1 and three R1b1b2); four Chinese (three R1b2b and one R1b1b2), 2 Chinese of the Hui tribe, and 6 Chinese of other ethnicities (Hazak R1b1b1, Tu R1b2b, and four Naxi R1b2b). Copyright © 2012 SciRes. 88 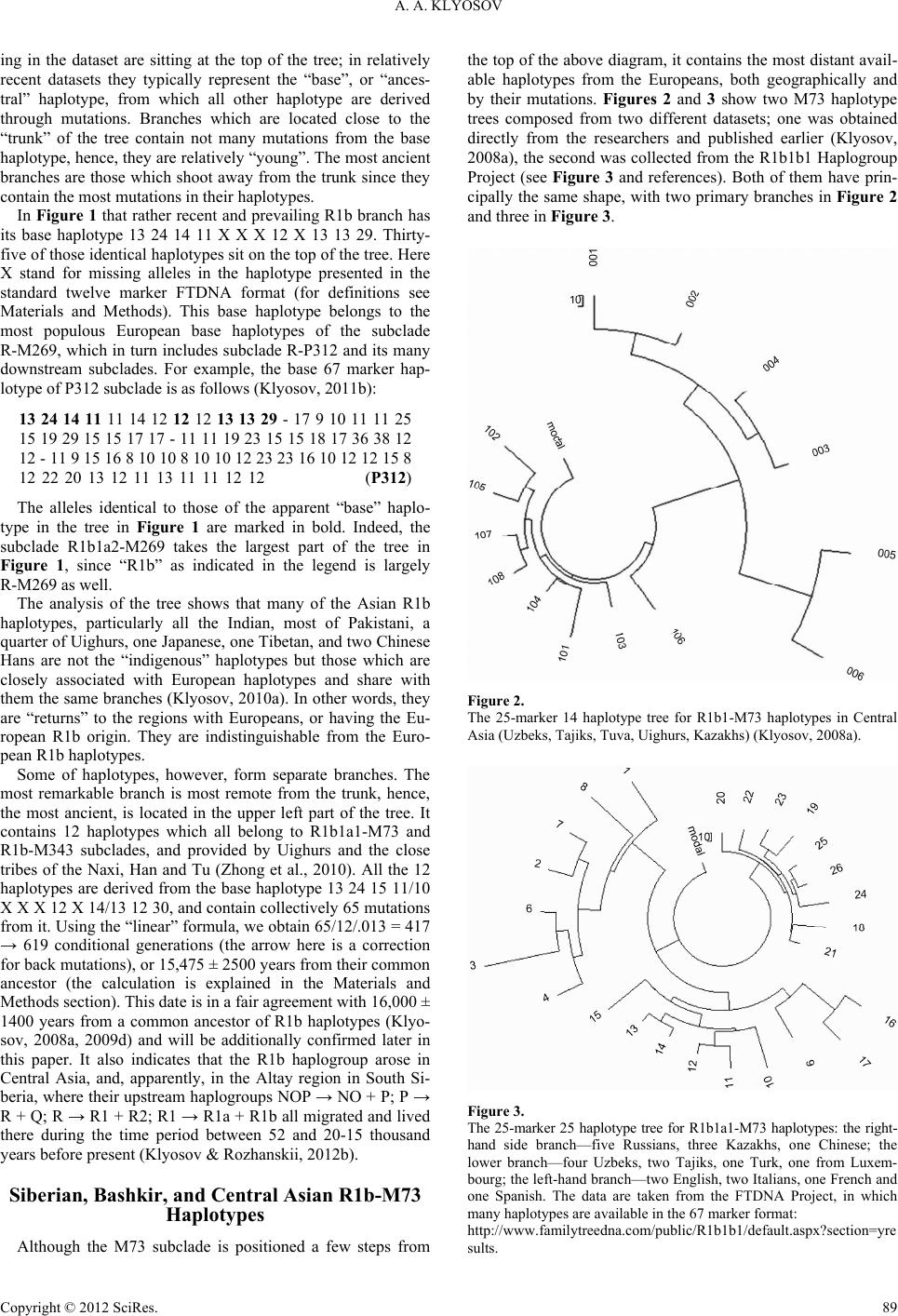 A. A. KLYOSOV ing in the dataset are sitting at the top of the tree; in relatively 1b branch has its ty the tree shows that many of the Asian R1b ha however, form separate branches. The m Although the M73 suned a few steps from th recent datasets they typically represent the “base”, or “ances- tral” haplotype, from which all other haplotype are derived through mutations. Branches which are located close to the “trunk” of the tree contain not many mutations from the base haplotype, hence, they are relatively “young”. The most ancient branches are those which shoot away from the trunk since they contain the most mutations in their haplotypes. In Figure 1 that rather recent and prevailing R base haplotype 13 24 14 11 X X X 12 X 13 13 29. Thirty- five of those identical haplotypes sit on the top of the tree. Here X stand for missing alleles in the haplotype presented in the standard twelve marker FTDNA format (for definitions see Materials and Methods). This base haplotype belongs to the most populous European base haplotypes of the subclade R-M269, which in turn includes subclade R-P312 and its many downstream subclades. For example, the base 67 marker hap- lotype of P312 subclade is as follows (Klyosov, 2011b): 13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 17 17 - 11 11 19 23 15 15 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12 (P312) The alleles identical to those of the apparent “base” haplo- pe in the tree in Figure 1 are marked in bold. Indeed, the subclade R1b1a2-M269 takes the largest part of the tree in Figure 1, since “R1b” as indicated in the legend is largely R-M269 as well. The analysis of plotypes, particularly all the Indian, most of Pakistani, a quarter of Uighurs, one Japanese, one Tibetan, and two Chinese Hans are not the “indigenous” haplotypes but those which are closely associated with European haplotypes and share with them the same branches (Klyosov, 2010a). In other words, they are “returns” to the regions with Europeans, or having the Eu- ropean R1b origin. They are indistinguishable from the Euro- pean R1b haplotypes. Some of haplotypes, ost remarkable branch is most remote from the trunk, hence, the most ancient, is located in the upper left part of the tree. It contains 12 haplotypes which all belong to R1b1a1-M73 and R1b-M343 subclades, and provided by Uighurs and the close tribes of the Naxi, Han and Tu (Zhong et al., 2010). All the 12 haplotypes are derived from the base haplotype 13 24 15 11/10 X X X 12 X 14/13 12 30, and contain collectively 65 mutations from it. Using the “linear” formula, we obtain 65/12/.013 = 417 → 619 conditional generations (the arrow here is a correction for back mutations), or 15,475 ± 2500 years from their common ancestor (the calculation is explained in the Materials and Methods section). This date is in a fair agreement with 16,000 ± 1400 years from a common ancestor of R1b haplotypes (Klyo- sov, 2008a, 2009d) and will be additionally confirmed later in this paper. It also indicates that the R1b haplogroup arose in Central Asia, and, apparently, in the Altay region in South Si- beria, where their upstream haplogroups NOP → NO + P; P → R + Q; R → R1 + R2; R1 → R1a + R1b all migrated and lived there during the time period between 52 and 20-15 thousand years before present (Klyosov & Rozhanskii, 2012b). Siberian, Bashkir, and Central Asian R1b-M73 Haplotypes bclade is positio e top of the above diagram, it contains the most distant avail- able haplotypes from the Europeans, both geographically and by their mutations. Figures 2 and 3 show two M73 haplotype trees composed from two different datasets; one was obtained directly from the researchers and published earlier (Klyosov, 2008a), the second was collected from the R1b1b1 Haplogroup Project (see Figure 3 and references). Both of them have prin- cipally the same shape, with two primary branches in Figure 2 and three in Figure 3. Figure 2. er 14 haplotype tree for R1b1-M73 haplotypes in Central The 25-mark Asia (Uzbeks, Tajiks, Tuva, Uighurs, Kazakhs) (Klyosov, 2008a). Figure 3. er 25 haplotype tree for R1b1a1-M73 haplotypes: the right- ?section=yre The 25-mark hand side branch—five Russians, three Kazakhs, one Chinese; the lower branch—four Uzbeks, two Tajiks, one Turk, one from Luxem- bourg; the left-hand branch—two English, two Italians, one French and one Spanish. The data are taken from the FTDNA Project, in which many haplotypes are available in the 67 marker format: http://www.familytreedna.com/public/R1b1b1/default.aspx sults. Copyright © 2012 SciRes. 89  A. A. KLYOSOV The tight 8-haplotype branch close to the “trunk” of the tree in 9 11 11 23 15 y ha imilar series of ten R haplo- ty e tree ha 11 13 17 12 12 12 13 13 30 - 15 9 9 11 11 23 15 and ure 3 has the following base ha 14 12 12 12 14 13 29 - 16 9 10 11 11 22 an , they all are X 13 14 13 30 - 15 10 d Siberian-Ural- C Figure 2 and the 9-haplotype right-hand branch in Figure 3 both have the same base haplotype as follows: 13 19 14 11 13 13 12 12 13 14 13 30 - 17 9 19 33 12 15 15 16 - 10 10 19 25 15 16 16 17 30 36 12 10 - 11 8 16 16 8 10 10 8 11 10 12 23 23 16 10 12 12 16 8 12 24 21 13 12 11 13 11 11 12 11 (M73, branch) The remarkable feature of this base haplotype (and ever plotype in the both branches) is the second allele, DYS390 = 19. In the 25 marker format, the branches contain 15 and 17 mutations respectively, which gives the same 1075 ± 280 years to the common ancestor in both instances. It should be mentioned here that a s -M73 haplotypes (with DYS 390 = 19) were obtained from the Bashkir population (Myres et al., 2010) near the border between Europe and Asia, on both sides of the Ural Mountains. Their base haplotype was 13 19 14 10 X X X 12 13 14 13 29 - 10 (the last marker is DYS461), and all hundred alleles in the ten haplotypes contained only three mutations; this gives 3/10/.018 = 17 generations, or 425 ± 250 years to their common ancestor. However, as it will be shown below, a common an- cestor of these and other R-M73 haplotypes lived more that 10 thousand years ago. What we see in these haplotypes are essen- tially “tips of the iceberg;” they are tremendously distant from each other and must be very remote from their common ances- tor in time. It seems that the “base” of the proverbial iceberg vanished without leaving their ancient haplotypes behind. In other words, we see here a series of present-day haplotypes which either arrived from elsewhere a few centuries earlier, or went through a severe bottleneck(s) of the population. The Bashkir’s (similar or slightly mutated) R-M73 pes were identified among Tatars and Mari populations, in the adjacent regions (ibid.). In the same study, the R-M73 were found also among Kabardino-Balkars, Mengrels, Turks, with all speaking Turkic or Uralic languages, which belong to the Altay language family. Another name for Mari people is Cheremis, meaning “people from the East” in a neighboring Komi language. Their base haplotype 13 22 14 11 X X X 12 13 14 13 32 - 10 differs by 7 mutations (!) in ten markers from the Bashkir base L23 haplotype, which places their common an- cestor by 9700 ybp (Klyosov, 2011a), and if haplotypes with DYS390 = 19 are included, their common ancestor moves deeper in time, to at least 16,000 years before present. Revisiting the tree in Figure 3, the lower branch in th s the following base haplotype (when extended to 67 mark- ers) 13 22 14 20 30 12 15 15 17 - 11 10 19 24 15 14 17 17 35 39 12 10 - 11 8 16 16 8 10 10 8 10 10 12 23 23 15 10 12 12 18 9 12 25 20 13 12 11 13 11 11 12 12 (M73, branch) its common ancestor lived 3525 ± 600 ybp. It corresponds to the right branch in Figure 2. The left-hand side branch in Fig plotype: 12 25 14 11 13 15 20 32 12 15 16 17 - 11 10 19 25 15 17 17 18 34 36 14 10 - 11 8 16 16 8 11 10 8 11 10 12 22 23 17 10 12 12 15 8 13 22 20 14 12 11 14 11 11 12 12 (M73, branch) d its common ancestor lived 3050 ± 600 ybp. All three branches are rather “young”, however very distant from each other. Their pair-wise mutational differ- ences are 38, 38 and 42, which translates into the separation time between their common ancestors as 11,500 - 13,200 years of the mutational evolution in their haplotypes (we call it the “lateral time” between two base haplotypes). All three M73 base haplotypes differ from each other collectively by 62 muta- tions, which place their common ancestor at ~7750 ybp. How- ever, their mutational difference is even more pronounced when compared with the base haplotype of R1b1a2-P312 subclade, ancestral to many European subclades of R1b1a2 haplotypes. For the above M73 base haplotypes it amounts to 48, 35 and 42 mutations, or up to 16,000 years of the mutational difference between the two base haplotypes. This places their common ancestor at 10,600 ybp. Another series of 26 of 12 marker M73 haplotypes was listed in (Malyarchuk et al., 2011), and were collected among the Siberian populations: Shors, Teleuts, Kalmyks, Khakass, Tuva, and Altayans. Those haplotypes split into two branches. The first one fits exactly to the first 67 marker base haplotypes with their available 12 markers; the second deviates from the base haplotype of the respective right-hand branch in Figure 2 in two alleles (marked): 13 19 14 11 13 13 X 13 22 14 11 13 16 X X 13 13 13 30 - 15 10 The first 12 marker base haplotype is a mixe aucasian population (if to consider their current bearers—Bash- kirs, Kalmyks, Tuva, Tatar, Kabardin, Russian, and Altayan). All 11 haplotypes of this branch have collectively 20 mutations, which gives 20/11/.024 = 76 → 83 conditional generations, or, 2075 ± 510 years to a common ancestor. The logarithmic method gives [ln(11/2)]/.024 = 71 → 77 generations, or 1925 years, which is nearly the same (2 haplotypes are identical in their base haplotypes among 11 haplotypes in the dataset. .024 is the mutation rate constant for the 12 marker haplotypes). The same but in 67 markers, Middle Asian M73 haplotypes showed 1075 ± 280 years to the common ancestor. Thus, the Siberia- Ural-Caucasian dataset is somewhat “older” compared to the Middle Asian dataset. The second 12 marker base haplotype was largely from Sibe- rian populations (Shors, Teleuts, Khakasses), plus a singular Mari, Turk and Tatar each. All 15 haplotypes of this branch had 22 mutations, which gives 22/15/.024 = 61 → 65 conditional generations; or 1625 ± 380 years to a common ancestor. The logarithmic method gives [ln(15/5)]/.024 = 46 → 48 genera- tions, or 1200 ± 580 years, which is the same within the margin of error. The 67 marker base haplotype, however showed 3525 ± 600 years to the common ancestor. Hence, now the Middle- Asian M73 branch is “older” compared to the Siberian. Overall, the Siberian and Middle Asian haplotypes were in about the same estimate of “age”. The two 12 marker haplotypes differ by 8 mutations, which sets their two common ancestors apart by 8/.024 = 333 → 492 generations, or 12,300 years. It gives (12,300+2075+1625)/2 = 7985 ybp to their common ancestor. One can see that the data obtained with the 67 and 12 marker haplotypes of the M73 subclade from Siberia and Middle Asia are fairly reproducible, and point to a common ancestor of Central Asian M73 haplo- types nearly 8000 ybp. It should be kept in mind that these data Copyright © 2012 SciRes. 90  A. A. KLYOSOV are related to currently living descendants of those common ancestors. We also need to be mindful that the 67 marker M73 Central Asian haplotype examples a distance up to 48 muta- tions from the European R1b1a2 haplotypes, which is up to 16,000 years of the mutational difference between the two base haplotypes. This places their (R-M73 in Central Asia and R-M269 in Europe) common ancestor at 10,600 years before present. R1 * b1-L278/R1b1 Subclade Haplotypes The the Projec M73 an are represented in th most ancient subclades of R1b are considered by t R1b1 (xP297); xP297 does not include subclades d M269, discussed in the preceding section of this article. All 68 haplotypes of the Project which are available in the 67 marker format are shown in the tree in Figure 4. The tree in- cludes haplotypes of subclade R1b1*, a para-group of R1b1-L278 (see the R1b diagram exampled previously), V88, its down- stream subclade V69, and a series of haplotypes R1b1a2-M269 which are in the Project apparently by mistake (however, useful in the tree as a reference branch). A detailed analysis of the tree was performed in (Klyosov, 2011a). Haplotypes of the R1b1* para-group e Project by either Ashkenazi Jews (from Hungary, Russia, Belarus) with their very recent common ancestors a few centu- ries ago, or by very assorted geography—from Italy, Puerto-Rico, Germany, Armenia, Turkey. Formally, a triple branch on the right of Figure 4 (haplotypes 1 - 10) has a common ancestor who lived about 5800 ybp. It does not carry any certain histori- cal meaning judging from their composition except, the time span corresponds to that of R-L23 subclade in the Caucasus and Anatolia (Klyosov, 2010d); R-L23 was certainly not alone there. Those assorted haplotypes could have been brought by their ancestors from Central Asia along with R-M269 followed by Figure 4. 67 marker haplotype tree of 68 haplotypes of subclades R1b1 1c-V88 (18 - 41), R1b1c4-V88-V69 (42 - 47), R1b1a2 L23 9 9 11 11 in the first 255 -V88 Subclade in Europe and Africa Hig- ure 4 16/17 9 10 11 12 an es * (1 - 17), R1b (48 - 68). The tree was composed from data of the FTDNA Project http://www.familytreedna.com/public/R1b1Asterisk/default.asp x?section=yresults. haplotypes, and eventually dispersed across Europe. he next small braTnch of six haplotypes (12 - 17) includes those from Central Asia (Uzbek), Armenia, Middle East (Iraq and Bahrain), India, and Germany (the last being relatively close to the Uzbek haplotype, within 4900 years of the muta- tional evolution). The Indian haplotype has DYS390 = 18, which brings it closer to the Central Asian haplotypes with DYS390 = 19, considered above. Others have DYS390 = 24 or 25. Their mutational deviations (without inclusion of the very distant haplotype) from base R1b1* haplotype 13 24 15 10 12 13/14 12 13 11/12 13 13 29 - 14 26 14/15 18 30 13/14 14 14/15 16 - 11 11 18 23 15 16 18 16 36 36 13 11 - 11 8 15 16 8 10 10 8 11 10 12 19 21 16 10 12 12 14/15 8 12 24 20/21 14 11 11 13 12 11 12 12 (R1b1*, branch) , 37, and 67 markers suggests 238, 217, and 23 generations from their common ancestor (without a correction for back mutations), that is 230 ± 11 generations, a fairly re- producible value. The most reliable value, for the 67 marker format, gives ~8400 years to their common ancestor. However, the Indian haplotype differs from the above base haplotype by 62.5 mutations (!), with its DYS390 = 18 contributing only 6 mutations into the difference. This places a timespan to the R1b1* common ancestor (of the available haplotypes) to 16,600 years before present. It certainly points to Central Asia, since the European-Middle Eastern R1b1* haplotypes are only ~8400 years “old” R1b1c aplotypes V88 occupy the left-hand side of the tree in F . Again, a number of their sub-branches are quite “young”. For example, a part of the branch belongs to Ashkenazi Jews (Russia, Ukraine, Hungary, Germany, France) with a common ancestor of only 350 ybp. Haplotypes of England and Scotland have a common ancestor who lived 650 ybp. However, a com- mon ancestor of the two sub-branches (Jews and the Isles) lived 6875 ybp, at the very bottom of the V88 subclade. Another series of V88 haplotypes identifies their route from Saudi Ara- bia (6225 ybp) via Spain to the Jewish community (1525 ybp, in the middle of the 1st millennium CE). The base haplotype of the V88 branch in Figure 4 is as follows: 13 24 16 10 13 14 12 12 12 14 13 29 - 26 14 19 29 12 12 15 15 - 11 12 21 23 15 15 18 19 33 35/36 12 11 - 12 8 15 16 8 10 10 8 9 11 12 22 23 15 11 12 12 15 8 12/13 23 20 13 12 11 13 11 12 12 12 (V88) d its bearer lived 6575 ± 700 ybp. V88 haplotypA rather extended series of 72 of 11 marker from Africa was provided in (Cruciani et al., 2010). They split into two rather distinct branches and an assorted series of hap- lotypes (Klyosov, 2010b). One branch of 37 haplotypes has the following base haplotype (in the format—413a,b 460, 461, GATA A10, YCAIIa,b after the first standard 12 markers, some of them are missing): 13 X 15 11 X X X X 12 X X X - 23 21 11 10 13 21 23 All of them contain 111 mutations, which gives 111/37/.02 = 150 → 176 conditional generations, or 4400 ± 610 years to their common ancestor. This African V88 base haplo- type does not fit with either of the Eurasian V88 base haplo- types, however, it fits rather well to the R1b1c4-V69 base hap- lotype (see the next section), whose common ancestor lived Copyright © 2012 SciRes. 91  A. A. KLYOSOV 4300 ± 600 ybp. The paper (Cruciani et al, 2010) suggested one more subclade named R1b1a4, with 29 haplotypes. Their base haplotype is identical with that shown above; all 29 haplotypes contain 76 mutations, and gives 76/29/.02 = 131 → 150 genera- tions, or 3750 ± 570 ybp, which is within the margin of error with the above date. Another branch of V88 African haplotypes is recent with a common ancestor of 750 ± 290 ybp. All 72 African haplotypes ha stward along the M This subclade es at the bottom of the tree in Fig 16 17 19 33 e ha r 43 ison with the entire R1b, and R-V69 is “y Subclade The last brane lower right- hand side has th 23 15 15 18 17 37 38 12 R- A ublications (Klyosov, 2008a, 2009d, 20 ting from Central Asia westward by tw e trees co 15 15 19 17 35 38 12 ve the same base haplotype as shown above with 201 muta- tions from it, which gives 201/72/.020 = 140 → 163 genera- tions, or 4075 ± 500 ybp, within the margin of error of that for the main V88 branch; however, the first is more accurate since it was calculated for the distinct branch. This date, 4300 ± 600 ybp, fits well with that for the migration route of R1b bearers from the Middle East we editerranean Sea coast to the Atlantic, which took place between 5500 and 4800 ybp (see below). It seems that the R1b-V88-V69 tribe may have split altering migration direction south, to Camer- oon and Chad, where R1b-V88 bearers live today. R1b1c4-V69 Subclade is represented by six haplotyp ure 4, with the base haplotype 13 23 15 11 13 15 12 12 13 13 13 29 - 16/17 9 10 11 12 27 14 19 29 12 12 14 15 - 11 12 21 23 15 36/37 12 12 - 12 8 15 16 8 10 10 8 10 11 12 21 24 15 11 12 12 14 8 12/13 22 20 13 12 11 13 11 12 12 13 (V69) All of them collectively have 106 mutations from the bas plotype, which gives 106/6/.12 = 147 → 172 generations, o 00 ± 600 years from a common ancestor. This is in agree- ment with the “age” of the upstream V88 subclade, of ~6575 ybp. The V69 base haplotype differs by 16 mutations from the base V88 haplotype (see above; some mutations are fractional), which sets apart their common ancestors by 16/.12 = 133 → 154 generations, or 3850 years, and places their common an- cestor at (3850 + 6575 + 4300)/2 = 7400 ± 800 ybp. This is within the margin of error with 6575 ± 700 ybp for the V88 upstream subclade. The analysis demonstrates that R-V88 is a rather “young” subclade in compar ounger”. It may have arisen on the R1b general migration route to the Middle East, for example, north-east or east of the Caspian Sea, in the Western Iran. R1b1a2-M269 ch on the tree in Figure 4 in th e base haplotype 13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 17 17 - 11 11 19 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12 (M269, a mix) It is exactly the base haplotype of subclades R-P312 and L21. They are typical and widespread European haplotypes. ll 20 haplotypes of the branch contain 365 mutations, which gives 365/20/.12 = 152 → 179 generations, or 4475 ± 505 years to their common ancestor. This, again, is a typical “age” for a mix of haplotypes of subclades R-P312 and R-L21 and their downstream subclades. The M269 subclade along with its downstream subclades were analyzed in many p 10c, 2010d, 2010e, 2011a, 2011b). In short, this subclade arose around 7000 ybp in Central Asia or in the eastern area of the Russian Plain—its immediate downstream subclade R-L23 (arose ~6000 ybp) is widespread among the Bashkirs in the South Urals, North Kazakhstan and adjacent regions, and can be seen in Russia. It migrated south to the Caucasus and be- yond, to Anatolia and the Middle East. Nearly all principal European and Middle Eastern subclades of the R1b haplogroup (currently more than 80 subclades) are derived from the R1b1a2-M269 subclade. It should be mentioned here, that two brother subclades, R1a and R1b, have been migra o quite different routes. While R1a were moving along the southern route from the Altay region across the Himalayas, Hindustan, the Iranian plateau, Anatolia and the balance of Asia Minor to the Balkans (Klyosov & Rozhanskii, 2012), R1b were moving along the northern route, from the same region across South Urals, Middle Asia, North Kazakhstan, Middle Volga, the Caucasus, and then split between the southward and west- ward directions. This migration pattern also explains why R1a, but not R1b bearers were found in the Eastern Himalayas (Kang et al., 2011; see also Klyosov & Rozhanskii, 2012). 37 and 67 marker haplotype trees of the R1b1a2*-M269 sub- clade are shown in Figures 5 and 6, respectively. Th ntain two distinct branches, one rather complex and obvi- ously “older”, with the base haplotype 12 25 14 11 11 14 11 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 16 17 - 10 11 19 23 Figure 5. The 37 marker 25-haplotype tree for R1b1a2*-M269 haplotype 6225 ybp) branch on the left and the bottom pre- s. The older (~ sent haplotypes from Turkey, Armenia, Italy, France, England. The younger (1000 ybp) branch in the upper right side repre- sents Ukraine, Belarus, Poland, and Kazakhstan. http://www.familytreedna.com/public/ht35new/default.aspx Copyright © 2012 SciRes. 92  A. A. KLYOSOV Figure 6. The 67 marker haplotype tree for R1b1a2*-M269 haplotypes, the sam own in Figure 5, however, with fewer haplotypes avail- - 11 9 15 16 8 10 10 8 11 10 12 23 23 16 10 12 12 15 8 an 12 12 13 14 29 - 17 9 10 11 11 25 younger mong R-L23 appar Plain, where Eur irs, a Turkic-language people who live la entral Asia, apparently from the So stic DYS393 = 12). In a re 20 15 16 17 - 11 11 19 23 16 15 17 17 36 37 12 act bo 11 19 23 16 16 18 17 36 37 12 e dataset as sh able. 12 13 22 20 13 12 11 13 11 11 12 12 (M269) d another, a tight, flat, “younger”, and largely Jewish branch, with the base haplotype. 12 24 14 10 11 14 11 15 19 30 15 15 16 16 - 11 10 19 23 17 16 17 17 38 39 13 12 - 11 9 15 16 8 10 10 8 11 10 12 21 23 16 10 12 12 14 8 12 22 20 13 12 11 13 10 11 12 12 (M269, Jewish) Mutation deviations between the two are shown in bold. The branch has 26 mutations in all six 67 marker haplo- types, which gives 26/6/.12 = 36 → 37 conditional generations, or 925 ± 200 years to a common ancestor. The 37 marker branch contains 31 mutation in nine haplotypes, which gives 31/9/.09 = 38 → 40 generations, or 1000 ± 200 years—essen- tially the same. This Jewish population exemplified with 14 of 37 marker haplotypes was analyzed earlier (Klyosov, 2008b) and determined that their common ancestor lived 1100 ± 250 ybp; the base haplotype is exactly as that of the “young” branch listed above. The older branch has a common ancestor who lived 6200 ± 900 ybp. The two base haplotypes differ by 22 mutations which separate them by 22/.12 = 183 → 224 generations, or 5600 years, and a common ancestor of both branches lived 6400 ± 900 ybp. This description gives us a general picture of migration of the Arbins from Central Asia (~16,000 ybp) westward during the next 10 - 9 thousand years, to about 7000 - 6000 ybp. Haplogroup R1b (mainly R1b1a2-L23) a Bashkirs, and in the Caucasus, Anatolia, Middle East ently arose on the eastern side of the Russian ope meets Asia, ~6200 ybp and migrated to the Caucasus and further South, to Anatolia and the Middle East. Another branch of L23 went westward, to Europe, approxi- mately 4500 ybp. The most eastern population with the prevailing R-L23 sub- clade is the Bashk rgely on both sides of the Ural Mountains and in North Ka- zakhstan. Frequency of R1b haplogroup reaches 84% in the Perm Bashkirs, 81% among Baimak Bashkirs, and lower fig- ures in other Bashkir tribes (Lobov, 2009). 29 of 10 marker haplotypes of subclade R-L23 of the Bashkirs were published (Myres et al., 2010), and their base haplotype is 12 24 14 10 X X X 12 12 13 13 30 - 10. This is a typical albeit slightly mu- tated L23 haplotype with its characteristic first allele DYS393 = 12. 26 haplotypes of those 29 were identical, as shown above, and the whole Bashkir L23 branch has a common ancestor who lived only 575 ± 175 ybp. However, this base haplotype differs from the European R-L23 base haplotype 12 24 14 10 X X X 12 12 13 14 29 - 11 by three mutations, which sets these two base haplotypes apart by 3/.018 = 167 → 200 conditional gen- erations—5000 years, and places their common ancestor at 5500 ybp. Other Bashkir haplotypes belonged to R-M269 (one haplotype), R-M73 (10 haplotypes), and R-U152 (8 haplotypes). The last series of haplotypes were all identical to each other, and thus are derived from a very recent common ancestor, who certainly had a European origin [U152 arose in Europe 4125 ± 450 ybp (Klyosov, 2011b)]. We therefore see today’s reflection of ancient migrations of the Arbins westward from C uth Siberian region, across the South Urals and further to the Russian Plain and then the Caucasus. Almost all R1b1a2 haplotypes in the Caucasus region belong to the subclade L23 (with its characteri cent paper (Balanovsky et al., 2012) 90 Caucasian haplotypes of R1b haplogroup were listed, and with exception of five R1b* haplotypes and a relatively “young” Abkhazian branch (Figure 7) 79 of 81 haplotypes (97.5%) in the dataset were of the L23 subclade (Note: the cited paper did not consider haplotype trees nor has analyzed the haplotypes in the manner presented here). The same pattern is observed with Armenian R1b haplotypes, and with most of Anatolian R1b haplotypes (Klyosov, 2010c, 11c). The 67 marker base R-L23 haplotype, obtained from an extended haplotype dataset from the world over (tree Figure 8) is as follows: 12 24 14 11 11 14 12 12 12 13 13 29 - 16 9 10 11 11 25 15 19 29 15 12 - 11 9 15 16 8 10 10 8 10 11 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12 (L23) The L23 base short Caucasian haplotype in Figure 7 fits ex- ly to the above haplotype (the matching alleles are marked in ld). A common ancestor of the L23 subclade lived ~6200 ybp (Klyosov, 2010d, 2011a). 81 Caucasian L23 haplotypes con- taining 425 mutations from their base haplotype, give 425/ 81/.035 = 150 → 176 conditional generations, or 4400 ± 490 years to their common ancestor. The “younger” date (compared with the “age” of L23 of about 6200 ybp) can be explained by a detailed consideration of an extended series of 107 of R-L23 haplotypes listed in the FTDNA Project (see the legend in Fig- ure 8). The tree in Figure 8 splits into two parts. On the left are 38 haplotypes, with the base 12 24 14 11 12 15 12 12 12 13 13 29 - 16 9 10 11 11 25 15 19 30 15 15 16 18 - 11 12 - 11 9 15 16 8 10 10 8 11 11 12 23 23 15 10 12 12 16 8 12 22 20 13 12 11 13 11 11 12 12 (L23, branch) Copyright © 2012 SciRes. 93 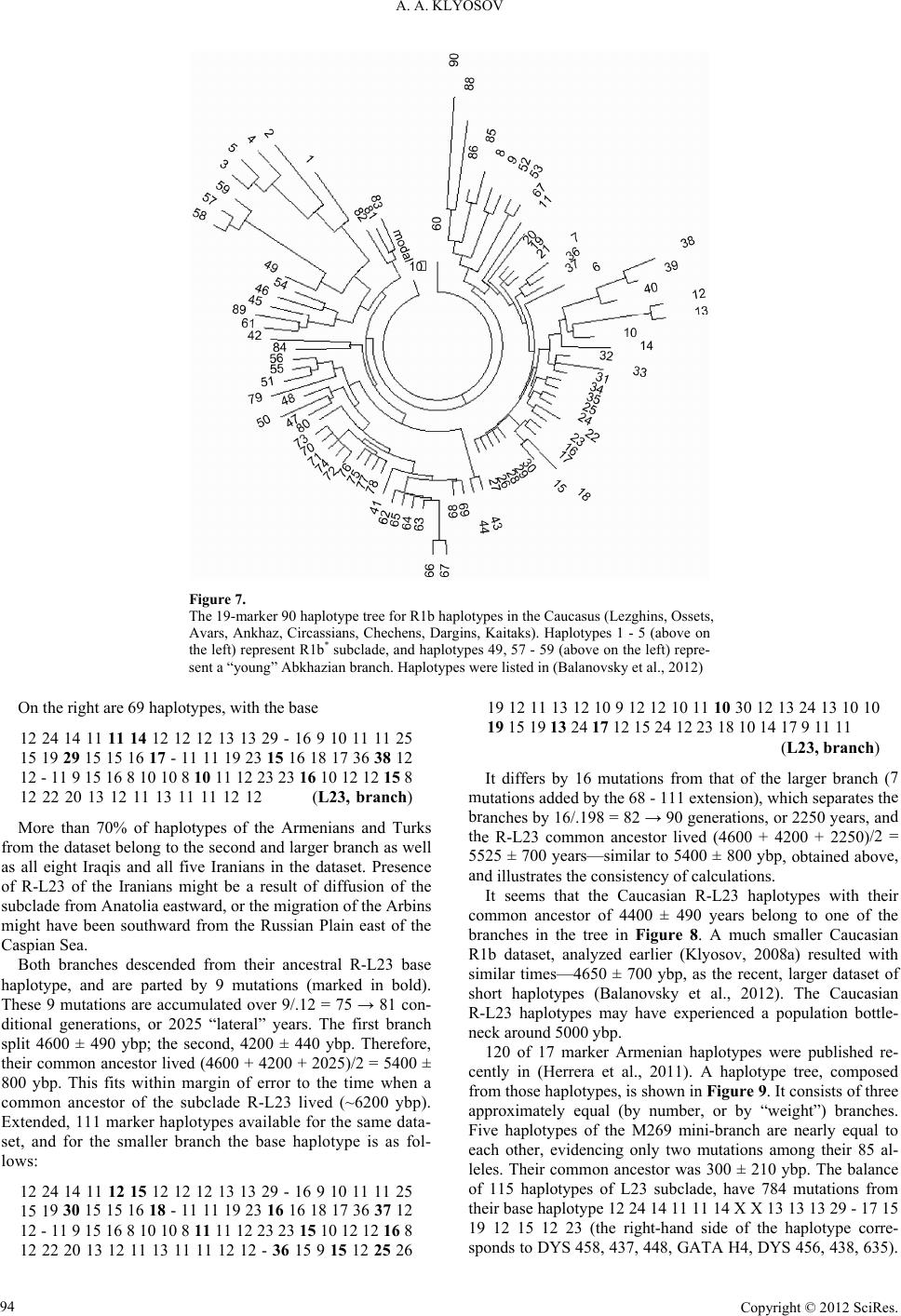 A. A. KLYOSOV Copyright © 2012 SciRes. 94 Figure 7. The 19-marker 90 haplotype tree for R1b haplotypes in the Caucasus (Lezghins, Osset Circassians, Chechens, Dargins, Kaitaks). Haplotypes 1 - 5 (above on * On the right are 69 haplo 12 24 14 11 11 14 12 12 12 13 13 29 - 16 9 10 11 11 25 17 36 38 12 s fro as 9 30 15 15 16 18 - 11 11 19 23 16 16 18 17 36 37 12 10 30 12 13 24 13 10 10 18 10 14 17 9 11 11 7 me br d th/2 = 5525 ± 700 years—similar to 5400 ± 800 ybpe, an s, Avars, Ankhaz, the left) represent R1b subclade, and haplotypes 49, 57 - 59 (above on the left) repre- sent a “young” Abkhazian branch. Haplotypes were listed in (Balanovsky et al., 2012) types, with the base 19 12 11 13 12 10 9 12 12 10 11 19 15 19 13 24 17 12 15 24 12 23 15 19 29 15 15 16 17 - 11 11 19 23 15 16 18 12 - 11 9 15 16 8 10 10 8 10 11 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12 (L23, branch) More than 70% of haplotypes of the Armenians and Turk m the dataset belong to the second and larger branch as well all eight Iraqis and all five Iranians in the dataset. Presence of R-L23 of the Iranians might be a result of diffusion of the subclade from Anatolia eastward, or the migration of the Arbins might have been southward from the Russian Plain east of the Caspian Sea. Both branches descended from their ancestral R-L23 base haplotype, and are parted by 9 mutations (marked in bold). These 9 mutations are accumulated over 9/.12 = 75 → 81 con- ditional generations, or 2025 “lateral” years. The first branch split 4600 ± 490 ybp; the second, 4200 ± 440 ybp. Therefore, their common ancestor lived (4600 + 4200 + 2025)/2 = 5400 ± 800 ybp. This fits within margin of error to the time when a common ancestor of the subclade R-L23 lived (~6200 ybp). Extended, 111 marker haplotypes available for the same data- set, and for the smaller branch the base haplotype is as fol- lows: 12 24 14 11 12 15 12 12 12 13 13 29 - 16 9 10 11 11 25 15 1 12 - 11 9 15 16 8 10 10 8 11 11 12 23 23 15 10 12 12 16 8 12 22 20 13 12 11 13 11 11 12 12 - 36 15 9 15 12 25 26 (L23, branch) It differs by 16 mutations from that of the larger branch ( utations added by the 68 - 111 extension), which separates th anches by 16/.198 = 82 → 90 generations, or 2250 years, an e R-L23 common ancestor lived (4600 + 4200 + 2250) , obtained abov d illustrates the consistency of calculations. It seems that the Caucasian R-L23 haplotypes with their common ancestor of 4400 ± 490 years belong to one of the branches in the tree in Figure 8. A much smaller Caucasian R1b dataset, analyzed earlier (Klyosov, 2008a) resulted with similar times—4650 ± 700 ybp, as the recent, larger dataset of short haplotypes (Balanovsky et al., 2012). The Caucasian R-L23 haplotypes may have experienced a population bottle- neck around 5000 ybp. 120 of 17 marker Armenian haplotypes were published re- cently in (Herrera et al., 2011). A haplotype tree, composed from those haplotypes, is shown in Figure 9. It consists of three approximately equal (by number, or by “weight”) branches. Five haplotypes of the M269 mini-branch are nearly equal to each other, evidencing only two mutations among their 85 al- leles. Their common ancestor was 300 ± 210 ybp. The balance of 115 haplotypes of L23 subclade, have 784 mutations from their base haplotype 12 24 14 11 11 14 X X 13 13 13 29 - 17 15 19 12 15 12 23 (the right-hand side of the haplotype corre- sponds to DYS 458, 437, 448, GATA H4, DYS 456, 438, 635).  A. A. KLYOSOV Figure 8. The 67-marker tree composed with 107 haplotypes of the subclade R-L23. 40% of the haplotypes belong to Armenians and Turks, followed by Iraqis and Italians (5% each). The haplotypes were roject http://www.familytreedna.com/public/ht35new/default.aspx It gives 784/115/.034 6250 ± 660 years fro typical timespan to a common ancestor of R-L23 subclade, and ped for subclades when tested several years ago as the typing was not 2/.09 = 164 → 196 generations, or 4900 ± 530 years from u- ta firy an of ce with 11% and others are in singular percentages (Lashgary et al., 2011). Such collected in the FTDNA P = 201 → 250 conditional generations, or m the common ancestor. This again is a al., 2008]. Among Jewish R1b haplotypes (most were not ty is in an agreement with the “age” of its upstream R-M269 sub- clade of ~7000 ybp. The same base haplotype, as shown in the preceding para- graph, was found in a dataset of 238 Armenian six-marker R1b haplotypes published earlier (Weale et al., 2001) and analyzed in (Klyosov, 2008a). It included haplotypes from six regions of Armenia, Karabakh, Iran, and other areas of the Armenian di- aspora. It can be presented as 12 24 14 11 X X X 12 X X 13 X. An average “age” of the common ancestor of R1b haplotypes in all the six regions was 5750 ± 1500 years, which is in line with other estimates for the R-L23 subclade. Considering other populations, containing a rather high share of R-L23 haplotypes, which in turn could be a tracer of ancient migration of the Arbins, one can mention Russia with their 37% of available R1b haplotypes having DYS393 = 12 (Roewer et al., 2008; Klyosov, 2010c and references therein) and the base haplotype 12 24 14 11 11 14 X X 12 13 13 29/30 - 16 15 19 (the last three alleles are of DYS 458, 437, 448), the same as R-L23, shown above, and with a common ancestor of the Rus- sian R1b of 6775 ± 830 ybp (Klyosov, 2009d); Anatolia and Lebanon with their 12 24 14 11 X X X 12 13 13 29 and 12 24 14 10 X X X 12 13 13 29 (mutated DYS391 1110 is marked) and with the respective common ancestors of 6000 ± 820 ybp (Anatolia) and 5200 ± 670 ybp (Lebanon) [Klyosov, 2010c; haplotypes were listed in Cinnioglu et al., 2004, and Zalloua et available), one branch remarkably resembled R-L23 haplotypes, and its 37 marker base haplotypes was as follows (Klyosov, 2008b): 12 24 14 10 11 14 12 12 12 13 14 29 - 17 9 10 11 11 25 15 19 29 15 15 16 17 - 11 11 19 23 16 16 18 17 37 38 12 12 (L23, Jewish) In this particular instance the dataset contained 42 of 37 marker haplotypes, which collectively had 620 mutations; this gives 620/4 a common ancestor. The base haplotype differs by 9 m tions from the “young” Jewish base haplotype above in its st 37 markers, which makes the 4900 ybp haplotype likel cestral. The oldest identified Jewish R1b base haplotype, is 5400 ± 500 ybp (Klyosov, 2008b). It has certainly appeared in the Middle East, during the Sumerian era. Sumers, the likely Bearers of R1b1a2 Haplotypes, and Their Descendant Assyrians Assyrians are one of the oldest surviving groups descended, as it is believed, from the historic Sumers. Among Assyrians, R1b is the major haplogroup, reaching 40% of the studied population; haplogroup J takes second pla Copyright © 2012 SciRes. 95  A. A. KLYOSOV Figure 9. The 17-marker tree composed with 120 haplotypes of the R-L23 haplogroup, except five haplotypes of the M269 subclade (numbers 1 - 5). The tree is composed based on data in (Herrera et al., 2011). a high percentage of R1b Mesopotamia. According t Appendix) half of their R1 subclade with DYS393 = 12, and all nine 12 marker haplotypes have 35 een 6000 an from the Pontic dle East Westward In the descriptions above we left bearers of R1b haplogroup ndants serve lished in (Barac et al., 2ericic et al., 2005, and pr haplogroup is very unusual in South o the Assyrian FTDNA Project (see b bearers belong to R-L23 Migrations of the Arbins Steppes, Asia Minor and Mid to Europe mutations, which gives 35/9/.02 = 194 → 240 conditional gen- erations, or 6000 ± 1200 years to their common ancestor. Only 6 haplotypes were available in the 25 marker format, and they have 48 mutations; they yield 48/6/.046 = 174 → 211 genera- tions, or 5275 ± 930 years. Those are indeed Sumerian times (e.g., Kramer, 1971). Most of the bearers of those haplotypes now live in Iraq and Turkey. Another half of the Assyrian hap- lotypes in the Project, mostly from Iran, have a slightly mutated “classical” European R1b1a2 base haplotype (with DYS464 = 15 15 17 17), and a common ancestor of 850 ± 360 ybp calcu- lated from the first 12 markers, and 1100 ± 280 ybp from the first 37 markers. This R1b evidence is clearly a relatively recent event in Assyrian population, brought from Europe. The R-L23 subclade is clearly traced from the Russian Plain south via the Caucasus, where it prevails among R1b haplo- types, and via Anatolia, where it is very pronounced; down to South Mesopotamia, where Sumers had lived betw d 4500 ybp (Kramer, 1971). Since timespans to common ancestors of those R-L23 haplotypes are around 6000 - 5000 ybp, it is quite likely that those common ancestors lived among the Sumers and their ancestors (in the Caucasus and Anatolia). It might be an additional feature for linguists, some of whom consider Sumerian as a remnant of a subgroup of the Dene- Caucasian language superfamily (e.g., Bengtson, 1997). in the Caucasus/Russian Plain/Pontic steppes, Anatolia, and Lebanon/Southern Mesopotamia around 6000 - 5000 ybp. All three areas were the relay regions for the Arbins to move fur- ther westward. Haplotypes of their present day desce as the tracers of those ancient migrations. A dataset of R1b1a2 haplotypes on the Balkans was pub- 003a, 2003b; P ivate communications with M. Pericic). It contains a series of obviously R-L23 haplotypes with a base 12 24 14 11 11 15 X X X 13 13 29, and a common ancestor of 4050 ± 890 ybp (Klyosov, 2010c). Another series of the Balkan R1b1a2 has the base 13 24 14 11 11 11 X X X 14 13 29, with a common an- cestor of 4975 ± 1300 ybp (ibid). Four mutations between them (including multi-copy mutations counted differently) place their common ancestor at ~7000 ybp to the time of ancient R-M269 haplotypes of the Russian Plain’s (or further to the east). Mi- grations westward from the Pontic steppes are supported also by multiple archaeological evidences, which include those dat- ing to the 3rd millennium BC (e.g., Mallory, 1989). The haplotype tree shown in Figure 8 provides some clues regarding possible directions of those ancient migrations. 40% of all 107 haplotypes of the tree belong to the Armenians and Turks; of those, 70% are haplotypes on the right-hand (larger) part of the tree. The upper left branch in the tree contains hap- lotypes from Russia, Lithuania, Poland, Croatia and Ireland. Copyright © 2012 SciRes. 96  A. A. KLYOSOV They all have the same pattern of mutations, with the base hap- lotype 12 24 14 11 11 14 12 12 12 12 13 29 - 16 9 10 11 11 25 15 19 31 14 15 16 18 (L23, mainly Eastern Europe) which differs from the base haplotype of the left-hand side 38-haplotype branch by three mutations (marked; some alleles are fractional). This places their common ancestor with the entire branch at 4600 ybp. This indicates that the above largely East-European branch split from the sizeable branch at the Rus- sian Plain in the middle of the 3rd millennium BC and appar- enf se - peared → 122 generations, or by 3050 “lateral” years (the 25w a fork, w only one mutation 12 → 13 in the first allele (D ese. In fact, the entire right-hand large branch in Figure 8 consists as R-P312, R-L21, R-U152, R-L2, R-L20, etc. The semon ancestor of 3550 ± 79a common ancestor who livestor of all of them lived 5025 ± 630 ybp, which fits the time and direction of ancient m ed by the linear method, and 43 tly migrated westward. That is one migration route (out o veral described) which explains how R-L23 haplotypes ap in Europe during those times. Another migration route took place from the Russian Plain southward, as indicated by another quite distinct branch on the opposite side (at 5 o’clock) of the haplotype tree. The branch includes haplotypes from Russia, Lithuania, Armenia (two hap- lotypes), Turkey (three haplotypes), Syria. The base haplotype is as follows: 12 24 14 11 11 14 12 12 12 13 13 30 - 16 9 10 11 11 25 15 19 28 15 15 16 17 (L23, mainly South) There are five full mutations between these two base haplo- types of the branches, both which includes Russian and Lithua- nian haplotypes, albeit, with a principally different history. One group of L23 bearers went west to Europe; another went south to the Caucasus and the Middle East. They are separated by 5/.046 = 109 is time is required on average to make five mutations in th marker haplotype). In other words, these two branches sho in migration routes of R-L23 from the Russian Plain estward and southward. One additional branch of R-L23 includes haplotypes from Armenia, Lebanon, Bulgaria, Italy, France, Spain, Germany. It is located at 7 o’clock on the haplotype tree in Figure 8. Their base haplotype 12 24 14 11 11 14 12 12 12 13 13 29 - 16 9 10 11 11 25 15 19 29 15 16 16 18 (L23, South and South-West) differs from the preceding discussed branch by only 2.6 muta- tions or by 1550 ”lateral” years. This points to R-L23 move- ment from the Middle East westward to Europe. Its neighboring branch, including haplotypes from Turkey (three haplotypes), Greece and Albania evidences nearly the same base haplotype as the above, with YS393) and 18 → 17 (17.3 on average) in the last allele. It is sentially the same migration route from Asia Minor to Europ of sub-branches with predominantly Armenian and Turkish (some Iraqi and Iranian) R-L23 haplotypes with singular inclu- sions of haplotypes from Greece, Germany, Netherlands, Eng- land, Scotland. This reflects migration routes of the Arbins from Asia Minor and the Middle East to Europe mainly be- tween 4500 and 3500 ybp. It may be expected that some migrations from the Middle East to Europe are associated with this mutation of DYS393 12 → 13 in the subclade R-L23. However, other, later migrations, could have occurred from the European continent eastward, and belong to more recent subclades, such as L51, L11, P312, U106, L21, U152, etc. (Klyosov, 2011b). Indeed, most Sardinian hap- lotypes have DYS393=13, such as the following base haplo- types on Sardinia (calculated from data provided in [Contu et al., 2008]): 13 24 14 11 11 14 X X X 13 X 29 13 24 14 10 11 14 X X X 13 X 29 13 24 14 10 11 15 X X X 13 X 29 The first one was presented on the tree by a series of identi- cal haplotypes which are obviously derived from the very re- cent common ancestor (Klyosov, 2008a, 2010c). They are in- distinguishable from common European R1b1a2- M269 sub- clades, such cond one has a timespan to its com 0 ybp, the third descended from ed 2900 ± 620 ybp. A common anc igration of the Arbins from the Middle East to Europe (how- ever, DYS393 = 13 would be unusual for them), and from Iberia up north to the continent, and subsequently populating Europe in all possible directions. One of the most common R1b base haplotypes in Sicily is as follows (Di Gaetano et al., 2009): 13 24 14 11 11 14 X X 12 13 13 29, which is identical to one of the Sardinian base R1b hap- lotypes above, and indistinguishable from common R1b sub- clades in Europe. The timespan to a common ancestor for that haplotype in Sicily calculated from the cited study is 4550 ± 1020 years. The same base haplotype is the most common in Italy (Capelli et al., 2007), with a timespan to the common an- cestor of 4125 ± 500 years calculat 00 ± 1160 years calculated by the logarithmic method (Kl- yosov, 2008a). There is one more, but very important migration route of the Arbins which does not practically include R-L23 haplotypes (only 5% of DYS393 = 12 among Iberian R1b haplotypes, it corresponds to a random mutation from the parent DYS393 = 13). It is a route from the Middle East westward along North African Mediterranean seacoast. R-L23 was either not repre- sented or did not survive along this route. It seems that bearers of R-V88 were part of the journey, however, split and went southward and settled in Central Africa (mainly Cameroon and Chad), mentioned previously. The migration of the Arbins from the Middle East to the Atlantic, then across the Strait of Gibral- tar to the Pyrenees took place from ~ 5500 - 5200 ybp to 4800 ybp when the Arbins landed in Iberia (see supportive references to archaeological data below). Part of the way from Egypt to Iberia could have been made by sea, details are not known as yet. There are some historical reports of arrival of the Egyptian military fleet to Iberia some 5000 ybp; there are some allegedly Egyptian mummies and fragments of Egyptian tombs exhibited in the Royal Academy of History in Madrid and in the Tar- ragona City Museum. Their status, however, is rather vague. It seems, nevertheless, that bearers of R1b subclades, mainly R-M269, and newly formed L51 and L11 (see below) had ar- rived in Iberia and this was the beginning of the archaeological Bell Beaker culture. Previous to discussing the Bell Beakers and history of the Arbins in Europe, it is worth mention of one more rather vague evidence of the R1b journey via ancient Egypt between 5500 and 5200 ybp. It concerns the alleged R1b haplotype of Phar- aoh Tutankhamun. Copyright © 2012 SciRes. 97  A. A. KLYOSOV Alleged Pharaoh Tutankhamun R1b1a1-M269* Haplotype and Its Possible History Recently the Swiss company iGENEA has published the al- leged 16 marker haplotype of the Pharaoh: 13 24 14 11 11 14 X X 10 13 13 30 - 16 14 19 10 15 12 (www.igenea.com/en/ index.php?c=62). Here the first 12 markers are shown in the FTDNA format, and the rest are DYS 458, 437, 448, GATA H4, DYS 456, 438. It is obviously not a typical European R1b1a2 haplotype, since it h o The mostype is th 17 9 10 11 11 25 tor of the two lived (6850 + 6200 + 3300)/2 = 81 - ce e up so m Altay, South Siberia, whe it is- doun R1b haplotypes ybp. Sea to Iberia, up north to the European continent. The first two principal routes were associated with bearers of the R wh - media very clos as DYS439 = 10, and not a common European 12. There are nly about 0.5% R1b haplotypes in Europe with DYS439 = 10. likely and the most closely related base haplot at of R1b1a2-M269, shown in Figure 5, in which the same markers as those available in the Pharaoh haplotype are noted in bold: 12 25 14 11 11 14 11 12 12 13 13 29 - 15 19 29 15 15 16 17 - 10 11 19 23 15 15 19 17 35 38 12 12 - 11 9 15 16 8 10 10 8 11 10 12 23 23 16 10 12 12 15 8 13 22 20 13 12 11 13 11 11 12 12 (M269) There are 6.8 mutations between the two 16 marker haplo- types (some alleles in M269 haplotype are fractional), which translates to 6.8/.0315 = 216 → 274 conditional generations, or 6850 years between them. Since Tutankhamun lived 3300 ybp, and the R-M269 base haplotype is 6200 ± 900 years “old”, then a common ances 75 ybp. This might have been either an ancient R-M269 an stor of the Tutankhamun from Central Asia, or the respectiv stream subclade. The main point here is that the Pharaoh haplotype is not me erroneously (or intentionally) picked European R1b1a2 haplotype; it is an archaic haplotype of R1b haplogroup, likely of the R-M269 ancient subclade. It should be mentioned here that the founder of the Egyptian pharaohs was Narmer, the origin of whom is not firmly known, and was the founder of the First Dynasty, living ca. 32nd cen- tury BC—around 5200 ybp. This date fits the migration times for the Arbins along the Mediterranean coast in North Africa. It cannot be excluded that the bearers of R1b haplogroup may have actually established the Royal lineage in ancient Egypt. It does not mean that the lineage was not interrupted through the Dynasties; however, it kept returning into the Dynasties, if the R1b origin of some Pharaohs is right. The Overview of the Migratory Path of the Arbins, Bearers of R1b Haplogroup, from Central Asia to Europe An overall map of R1b migration routes from the very “be- ginning”, where “beginning” is undefined in detail thus far and can be estimated between 16,000 and 12,000 ybp, and until their arrival to the Pyrenees by 4800 ybp as future Bell Beakers, is shown in Figure 10. Their migratory path was slowly taking place from Central Asia, apparently fro n some very different R1b haplotypes were discovered. As described in the first sections of this paper, their tremen s mutational difference with European and Middle Easter places their common ancestors at ~16,000 The legend to Figure 10 describes those Central Asian/Si- berian populations. There are many Neolithic, Chalcolithic and Eneolithic archaeological cultures in the area, such as Tersek, Ural, Surtandi, Mahandzhar, Iman-Burluk, Botai, Atbasar, Kel- teminar, and other archaeological Central Asian cultures in present-day Russia (e.g, Zakharov, 2010), which might be as- signed to the Arbins; however, it would be premature to assign any of them to R1b or to any other haplogroup. Such a task is quite new for archaeologists. It is tempting to point at Serogla- zovo, Khvalyn, Samaran, Middle-Volga and adjacent archaeo- logical cultures of 12,000 - 5000 ybp of the European Russian east as the most likely R1b cultures. We cannot, however for the same reason of prematurity and it would be irresponsible to suggest such at this time. The same may be said for Yamnaya, Catacomb and neighboring archaeological cultures of Central and South Russia, which apparently were shared by both R1b and R1a bearers, albeit in different time periods. R1b before 5000 ybp, R1a after 4500 ybp have confused archaeologists who have observed “different roots” of those cultures, spread- ing in different directions and in different times. The map shows that current bearers of R1b spread over Cen- tral Russia up to Arkhangelsk on the White Sea. Very likely it was a relatively recent relocation although it remains to be de- termined. Currently there are only about 5% of R1b bearers in the European part of Russia. As it was described above in this study, the Arbins went South through the Caucasus to Mesopotamia and the Middle East around 6000 - 5000 ybp; established the Sumer civilization; went westward via Egypt to the Atlantic, and across the Gibral- tar Strait to the Pyrenees. On their way some R1b-V88 bearers split; went deep into Africa, and currently populate Cameroon and Chad in appreciable amounts (see legend to Figure 10 and description and references above). By 4800 ybp the Arbins have reached Iberia to become the first Bell Beakers. This date was obtained for a common ancestor of haplotypes of P312 and U106 subclades (see the next Section), and is supported by archaeology data (Cardoso & Soares, 1990; Martinez et al., 1996; Cardoso, 2001; Muller & Willigen, 2001; Nocete, 2006). Several Entry Routes of R1b1a2 Haplotypes to Europe: From the Russian Plain, from Asia Minor/Middle East, from the Pyrenees To sum up the preceding section, the Arbins were entering Europe from the east by several routes: from the Russian Plain (between the Pontic Steppes and the Baltic Sea); from Asia Minor and the Middle East; and after a long way around Medi- terranean -L23 subclade. The Iberian route was made by mainly M269 ich at the time of entering the Pyrenees split R-L51 and im tely after R-L11. Both are very similar and have e timespans to their common ancestors, as it is shown in the next section. In 4850 ybp L11 promptly split off two “brother” subclades, P312 and U106 (Klyosov, 2011b) which after a long “population bottleneck” on the edge of extinction, eventually survived and expanded around 4000 - 3700 ybp, and actively populated Europe, first as Bell Beakers, between 4000 and 3000 ybp, and then up to the era of Ancient Rome, Gauls and Celts, mentioning only those names which present certain “milestones” in history. In fact, there were dozens if not hun- dreds of ancient R1b tribes in Europe. Copyright © 2012 SciRes. 98  A. A. KLYOSOV Copyright © 2012 SciRes. 99 Figure 10. The map of R1b haplotype locations considered in this study, until they arrived to Iberia ~4800 ybp. The overall migration path of the Arbins had apparently started in South Siberia, where present day R1b haplotypes of the Shors, Teleuts, Altayans, Uighurs, Khakass, Tuva people (the right-hand side pins) are tremendously different from those in Europe and the Middle East. The migration route continued through the regions in Middle Asia and Russia, where Bashkirs, Kazakhs, Uzbeks, Mari, Tatars, Kalmyks presently live. From the Central Russia region the Arbins moved to the Cauca- sus and further South to Anatolia, Middle East, present day Iraq, Lebanon, and then westward along the Mediterranean Sea onto Northern Africa t the Atlantic. On that way R1b-V88 bearers split and went deep into Africa, where their descendants currently live in Cameroon and Chad (Cruciani e eir migratory path into Africa is reproduced here from the above reference. The map shows R1b locations previous to arrival to the strea he ea. Cu- rope, ed a m o t al., 2010). Th Pyrenees ~4800 ybp, from where they moved North as Bell Beakers and populated Europe between 4800 - 3000 ybp and later. Main R1b1a2 Subclades on the European Continent: Entering Europe 4800 - 4500 ybp from the East and from the South West Nearly all haplotypes of the subclade R-M269 6000 - 5000 ybp belonged either to R-M269* para-group, or to its down- R1b1a2a1a1b4 L21/M529 R1b1a2a1a1b4b M222 R1b1a2a1a1b4g1 L226 R1b1a2a1a1b4h1 P314 The above diagram shows that the immediate downstream subclades of RSOGG-2012, in an abbreviateddes is much more understood ra- tion of the Arbins is The question is—wdes could have arisen? If they are d have split in Asia Minor, thePlain, and enter Europe from ther, that is only L51 or only L11 tion bottlenecks, hence, look “you be (in terms of mutations and th, th b- cl m subclade R-L23. The Arbins who had migrated from t st to Europe during those times mostly carried DYS393 = 12 rrently we see only those R1b1a2 (xL23) haplotypes in Eu which are derived from common ancestors who liv -L23 were L51 and then L11 (I form). The dynamics of these subcla via Iberia into the continent where the mig beakers. ing identified with the Bell Be aximum ~4500 ybp. Either descendants of more ancient L23 ancestors have not survived into the present time or there were none from the eastern Russian Plain, Asia Minor, and the Mid- dle east. R1b M343 R1b1 L278, M415, P25_1, P25_2, P25_3 R1b1a P297, L320 R1b1a1 M73, M478 R1b1a2 L265, M269, M520, S3, S10 here those L51 and L11 subcla 6000-5000 years “old”, they coul Middle East or on the Russian ese. The “intraclade” haplotyp subclade, might reflect popula nger” than they in fact should e respective TMRCA). However eir “interclade” comparison could reveal their lost (due to bottlenecks) timespans to more ancient common ancestors. To analyze those subclades, a combined L51-L11 haplotype tree is shown in Figure 11. One-third of haplotypes on the tree belong to the L51 su R1b1a2a L23/S141, L49.1 R1b1a2a1a L51/M412/S167 R1b1a2a1a1 L11, P310S129, P311 ade, and they occupy the right side. Another two-thirds are L11 haplotypes, which are on the left and make some “insert” branches on the right. The 67 marker haplotypes of L51 and R1b1a2a1a1a U106/S21 R1b1a2a1a1b P312/S116 R1b1a2a1a1b3 U152/S28  A. A. KLYOSOV Figure 11. The 67-marker tree composed with 42 haplotypes of the R-L51 subclade (15 haplotypes on the right, numbers between 1 and 23) and R-L11 (27 haplotypes on the left, numbers between 26 and 61). Haplotypes are listed in the FTDNA public Project http://www.familytreedna.com/public/ht35new/default.aspx. L11 subclades are very similar, therefore the tree could not distinguish them in a ce, the mix of the branches. The tree als clades is also very sim indicates the “age”) of same. The base haploty y of th r ~2n 28 2 = ir co = fits well with the arrival time of the Arbins to Iberia (4800 ybp), to the distribution pattern of the current bearers of L51 in yrenees and immedi- , see [Myres et al., . It is practically absent in European South-East, East, and st (ibid.). Analysis of the L11 subclade is more complicated since its nd the tree by at least four branches, ea etween them (marked in bold) present in fa number of cases; hen o shows that the “age” of the two sub- ilar, since the “height” (which generally the branches around the tree is about the pe of R-L51 subclade is as follows Europe (the highest frequency is in the P ately up north in France and on the Isles 2010] North-Ea 13 24 14 11 11 14 13 12 12 13 13 29 - 17 9 10 11 11 25 15 19 30 15 16 17 18 - 11 11 19 23 16 15 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 16 8 12 22 20 14 12 11 13 11 11 12 12 (L51) It deviates from the base R-L23 haplotype (5400 ± 800 ybp) by 9 mutations (12 mutations are marked above, but man em are fractional), that is by 9/.12 = 75 → 81 generations, o 025 years of the “lateral” distance. All 15 haplotypes contai 0 mutations from the base haplotype, which gives 280/15/.1 156 → 184 generations, or 4600 ± 535 years from the mmon ancestor, L51. In turn, it gives (2025 + 5400 + 4600)/2 6000 years, which is an “age” of a common ancestor of both L23 and L51 subclades. Obviously, it is the common ancestor of L23 subclade himself. In other words, the “age” of the L51 subclade as 4600 ± 535 years is obtained from mutations on the L51 branch on the tree (Figure 11) and confirmed 1) by a mutational distance from the base haplotype of the parent L23 subclade, 2) by the “age” of the L23 subclade, determined earlier, and 3) by the general phylogeny of the R1b subclade. The “age” of the L51 subclade haplotypes are spread arou ch with its common ancestor. They differ from each other by 32 mutations which gives 32/4/.12 = 67 → 72 generations, or 1800 years below their average “age” ~2500 years. Therefore the “age” of the R-L11 subclade is ~4300 years, which is in- deed consistent with 4600 ± 535 years for the L51 subclade. The base haplotype for the L11 subclade is as follows: 13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 30 15 15 17 17 - 11 11 19 23 16 15 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12 (L11) Five deviations b ct 3.7 mutations (since most of them are fractional), which gives 3.7/.12 = 31 → 32 generations; that is only 800 “lateral” years between them. This confirms that L51 and L11 subclades are very close to each other in time. Their common ancestor (which presumably should be L51) lived (800 + 4600 + 4300)/2 = 4850 ybp. It seems that L11 split off only ~250 years later. All of them are likely to have established the archaeological Copyright © 2012 SciRes. 100  A. A. KLYOSOV Bell Beaker culture. The oldest artifacts of the Bell Beakers were found in So W C 13 29 - 17 9 10 11 11 25 1 There are 5.5 (maations between them (D d 4175 ± 430 ybp (K+ 41 - clt ths, dee is - sp = 2 an an with seafaring carriers of L11* bearers from Ib d of R1b (with sub- cl se haplotype differs from the “b span to its com da s exs sh - nie es me time. It only confirms that their co Portugal, dated 4800 - 4600 ybp (Cardoso & ares, 1990; Martinez et al., 1996; Cardoso, 2001; Muller & illigen, 2001; Nocete, 2006). Main R1b1a2 Subclades on the European ontinent: Population of Europe after 4800 ybp, Main Subclades on the Isles and Their Likely Origin The phylogeny of R1b (see the diagram above) shows that the further downstream subclades of R-L11 are P312 and U106. Their 67 marker haplotypes are as follows: 13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 17 17 - 11 11 19 23 15 15 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12 (P312) 13 23 14 11 11 14 12 12 12 13 5 19 29 15 15 17 17 - 11 11 19 23 16 15 17 17 37 39 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 13 12 (U106) rked in bold) mut YS456 in P312 fluctuates between 15 and 16 in various datasets), which separates the two base haplotypes by 5.5/.12 = 46 → 48 generations, or by 1200 years. Since common ances- tors of both subclades lived 4350 ± 700 an lyosov, 2011b), their common ancestor lived (1200 + 4350 75)/2 = 4850 ± 700 ybp. Indeed, it is the “age” of the sub ade L11 within margin of error. Generally, it is safe to accep e “age” of both P312 and U106 as ~4200 ybp, since estimate pending on a dataset, are fluctuating around this value. Ther 1200 years between them (see above) which defines a time an to their common ancestor, L11, as (4200 + 4200 + 1200)/2 4800 ybp. As it was stated above, the four subclades, L51, L11, P31 d U106 historically represent the first wave of Bell Beaker movement from Iberia to the European continent. The highest frequency of P312* subclade in Europe is currently observed in Iberia (Myres et al, 2010), however, it is naive to judge ancient locations of the subclades based on their current distribution. The mixing of them on the Continent has been too intense in the past millennia to make any unambiguous extrapolations. The current regional distribution of the R-M269’s downstream subclades which moved from the Pyrenees nearly five millennia ago is classically non-informative, except in instances of some local subclades. One of them is R-M222. It seems that the history of R-M222 appearance on the Isles may have beg eria to the Isles some 4500 - 4000 ybp. However, it is just one theory. Other involve migrations of L21 bearers to the Isles by land. Then, P312, a downstream of L11, is also intensely rep- resented on the Isles along with its downstream subclades, as well as its “brother” subclade U106 and its downstreams (ibid). L21, a downstream of P312, is abundant on the Isles, unlike its “brother” subclade U152 (ibid). This ancient migration of the Arbins from Iberia to the Isles explains some genetics data on the “Spanish origin of the Irish”. M222, a downstream of L21, is among the most represented R1b subclades on the Isles. A massive Ireland Heritage FTDNA project (see Appendix) lists several thousan ades) haplotypes in the Isles, of which M222 is the largest, and includes about 25% of all R1b (Klyosov, 2010e). The sub- clade is expressed the most in Ireland. In Scotland it is observed mainly in the lowlands and in central region, and poorly pre- sented in England. The base haplotype of the M222 subclade is: 13 25 14 11 11 13 12 12 12 13 14 29 - 17 9 10 11 11 25 15 18 30 15 16 16 17 - 11 11 19 23 17 16 18 17 38 39 12 12 - 11 9 15 16 8 10 10 8 10 10 12 21 23 16 10 12 12 16 8 12 25 20 13 12 11 13 11 11 12 12 (M222) It is of interest that the above ba rother” base haplotypes of P312 and U106 by 18 mutations in each case (in fact, 15 mutations, since some mutations are fractional). It shows that P312 and U106 are indeed very close (in time and in heritage) base haplotypes, and that their com- mon ancestor, a bearer of L11 base haplotype, lived very close in time to both of them. 15 mutations place their common an- cestor at 4600 ± 500 ybp. Multiple analyses of M222 haplotype datasets gave a time- mon ancestor of currently living M222 descen- nts of 1450 ± 160 years (Klyosov, 2010e). However, cros amination of a number of lineages in the M222 subclade ha own that the M222 lineage arose not later than in the begin ng of the Common Era, some 400 years earlier than the abov timate. It points to a possible population bottleneck between the time when M222 arose, and the expansion of the subclade in the middle of the 1st millennium CE. One cannot possibly say, at least yet, where specifically the M222 subclade first appeared (that is, the G → A single nucleotide mutation oc- curred in certain nucleotide of Y chromosome of a certain indi- vidual, the bearer of the upstream L21 subclade), whether in the Isles, in North Western Europe, or elsewhere. We also cannot determine specifically against what R-M222 direct descendants have been struggling for survival for several centuries. Was it a long climatic event? The Roman invasions? Epidemics? Fam- ine? or a combination? We know, however, that it was the Isles where this mutation on the Y chromosome was eventually car- ried on and flourished. The phylogenic diagram shows that the M222 mutation arose concurrently with its “brother” SNP mutations which now de- fine subclades L226 and P314. The first one has a common ancestor who lived 1500 ± 170 ybp, which is at the same time with that of M222, albeit in L226 the subclade-defined muta- tion was C T. Its base haplotype is as follows: 13 24 14 11 11 14 12 12 11 13 13 29 - 17 8 9 11 11 25 15 19 29 13 13 15 17 - 11 11 19 23 15 15 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 15 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12 (L226) It differs by as many as 21 mutations from its “brother” M222 subclade, despite the observation that their common ancestors lived at the sa mmon ancestor, the founder of subclade L21, lived around 4100 ybp. It also shows that L21 formed almost immediately from P312 (see the phylogenic diagram) since P312 arose 4350 - 4200 ybp and L21 around 4100 ybp. Their 67 marker haplo- types are identical, within some fractional mutational differ- ence: 13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 17 17 - 11 11 19 23 15 15 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 Copyright © 2012 SciRes. 101  A. A. KLYOSOV 8 12 22 20 13 12 11 13 11 11 12 12 (P312) 13 24 14 11 11 14 12 12 12 13 13 29 - 17 9 10 11 11 25 15 19 29 15 15 17 17 - 11 11 19 23 16 15 18 17 36 38 12 12 - 11 9 15 16 8 10 10 8 10 10 12 23 23 16 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12 (L21) The only apparent difference, in DYS456, is also fractional, since this allele fluctuates in P312 between the values of 15 and 16. Despite their similar chronology, subclades M222 and L226 have quite different histories for the preceding 2600 years. Yet an , sin n hap; ho 2 ac mutations, respectively. It is a third, completely differ- en a rather robust system of D this paper, are as follows (iplotype per the conditional genees show exam- pl ht x?id=9 using software PHYLIP, Phm (see Klyosov, 2009c, 20ons for back mutations wc). Margins of erlyosov, 2009c). rmined by minimi- za plotype is one wr of mutations in the dacestor. The base haplo- ty losest approximation to th two base haplotypes is determined mutations ed num- other “brother” subclade, P314 (more accurately, P314.2 ce the same P314 SNP mutation, T → C, has occurred i plogroup H2a), is somewhat “older”, with its 2225 ± 300 yb wever, it could be within the same time frame when M22 tually arose. Its base haplotype 13 23 14 11 11 14 12 13 13 13 13 29 - 17 9 10 11 11 26 15 19 29 15 15 16 17 - 10 11 19 23 16 15 18 18 37 38 12 12 - 11 9 16 16 8 10 10 8 11 10 12 23 23 17 10 12 12 15 8 12 22 20 13 12 11 13 11 11 12 12 (P314.2) is again remote from those of M222 and L226 subclades, by 24 and 17 t lineage in the L21 family. The 10 full mutations from its L21 parent base haplotype gives 10/.12 = 83 → 91 generations, or 2275 years of the mutational difference, and places the be- ginning of the L21 subclade at (2275 + 4100 + 2225)/2 = 4300 years, which is within margin of error from 4100 years esti- mated above. All these base haplotypes along with the intra- clade and interclade datings form NA genealogy of the Arbins in Europe and Asia. Materials and Methods Four thousand four hundred eight (4408) of R1b haplotypes (with subclades) were collected in databases from FTDNA, YSearch, and in peer review publications. The methodology of haplotype datasets analysis was de- scribed in the preceding publication in this journal (Rozhanskii & Klyosov, 2011). In this study the linear and the logarithmic method were employed; the latter when the base haplotype in the dataset was easily identifiable, as described in (Klyosov, 2009c). The mutation rate constants are listed in (Klyosov, 2009c; Rozhanskii & Klyosov, 2011), and for a number of cases are given in the text of this paper. The most widely used mutation rate constants, including those employed in n a number of mutations per ha ration of 25 years); the referenc es of the haplotypes in the format indicated: .013, the 8 marker haplotype (Zhong et al.) .017, the 8 marker (Contu et al.) .018, the 10 marker (Myres et al.) .020, the 11 marker (Cruciani et al.) .019, the 11 marker (Zalloua et al.) .020, the 12 marker FTDNA format .024, the 12 marker (Malyarchuk et al.) .034, the 17 marker Y-filer format (Herrera) .035, the 19 marker (Balanovsky et al.) .046, the 25 marker FTDNA format .090, the 37 marker FTDNA format .120, the 67 marker FTDNA format .198, the 111 marker FTDNA format FTDNA haplotype formats are given in tp://www.familytreedna.com/faq/answers.asp Haplotype trees were composed ylogeny Inference Package progra 09d and references therein). Correcti ere introduced as described in (Klyosov, 2009 ror were calculated as described in (K Base haplotypes in the dataset were dete tion of mutations; by definition, the base ha hich has the minimum collective numbe taset, derived from one common an pe is the ancestral haplotype or the c e latter. A timespan to the common ancestor of as follows: 1) count the number of between the two base haplotypes, 2) divide the obtain ber by the mutation rate constant, 3) introduce a correction for back mutations, calculated using the following formula (Klyo- sov, 2009c): obs obs λ λ1expλ 2 (1) where: obs λ = observed average number of mutations per marker in a dataset (or in a branch, if the dataset contains several branches/ lineages). λ = average (actual) number of mutations per marker cor- rected for back mutations, 4) add the obtained value, multiplied by 25 (years), which represent the “lateral” timespan between times of appearance of the two base haplotypes, to TMRCAs for the both base haplotypes and divide by 2. The result repre- sents the TMRCA (time for the most recent common ancestor) for the two base haplotypes under study. The following exam- ples are based on specific cases in this study. Example 1: Calculation of a timespan to of the branch (the most remote on the hap ) which contains 12 of eight marker haplotypes collec- a common ancestor lotype tree shown in Figure 1 tively having 65 mutations from their base haplotype. Since the mutation rate constant for these haplotypes equal to 0.013 per haplotype per a conditional generation (25 years), we have 65/12 .013 =/ 417 → 619 generations, that is 619 × 25 = 15,475 years to a common ancestor of the branch. The arrow shows a correction for back mutations. This correction can be calculated using a formula (1) as follows. Since the observed number of mutations per marker is 65/12/8 = 0.677, we employ formula (1) and obtain obs 11exp 0.677λ 2 The obtained number of 1.484 is the coefficient of the cor- rection for back mutations. Therefore, by multiplying 417 × 1.484, we obtain that the corrected number of generations is 619, that is 619 × 25 = 15,475 years. This is usually designates as 417 → 618 (generations). Since for 65 mutations in the dataset the margin of error is 15.93% (calculated as explained in Klyosov, 2009c), we at last obtain the timespan to a common ancestor of the haplotypes to be equal to 15,475 ± 2500 years. Example 2: Two 67 marker base haplotypes of subclades R-P312 and R-U106 differ by 5.5 mutations (see the text above). Applying the same rule explained in the preceding ex- Copyright © 2012 SciRes. 102  A. A. KLYOSOV ample, we get 5.5/.12 = 46 → 48 generations, that is 1200 years from a common ancestor of the two base haplotypes (that is, of the two subclades). Should the two base aplotypes have the same “age” (the same TMRCA), their common ancestor lived 60 ybp (the timeframe between of R1a and R1a1 is uncertain) and R1b ~16, roto-Kugran” and/or “K lture and the state and migrated westward to Europe by la guages were sp . M., Janicijevic, B., Parik, J., Rootsi, S. et al. (2003a). Y chr Forensic Science International, 138, Barac, L uman Genetics, 11, 535- 542. doi:10.1038/sj.ejhg.5200992 h 0 years “deeper” in time from either one of them. However, in this particular case the two TMRCA are equal to 4350 and 4175 years, respectively. Therefore their common ancestor lived (1200 + 4350 + 4175)/2 = 4850 years ago. Conclusion The results of this study lend a support to the theory that haplogroup R1b arose in Central Asia, apparently in South Siberia or the neighboring regions, around 16,000 years before present. The preceding history of the haplogroup is directly related to the appearance of Europeoids (Caucasoids) ~58,000 ybp, likely in the vast triangle that stretched from Western Europe through the Russian Plain to the east and to Levant to the south, as it was suggested in (Klyosov, 2011d). A subse- quent sequence of SNP mutations in Y chromosome, with the appearance of haplogroups NOP ~48,000 ybp and P ~38,000 ybp in the course of their migration eastward to South Siberia, eventually gave rise to haplogroup R ~30,000 ybp and R1 ~26,000 ybp, and then to haplogroup R1a/R1a1 ~20,000 the appearance 000 ybp (ibid.). Based on the respective syllables, we call bearers of R1a the Aryans, and those of R1b the Arbins. In the first case that name is justified since the bearers of the R1a haplogroup became the legendary Aryans who arrived in the Hindustan and Iranian Plateau ~3500 ybp. In other words, those Aryans belonged to R1a haplogroup, hence, the double meaning (albeit coinciding) of the term the Aryans. The Arbins is a convenient common term in avoidance of the repetition: “The bearers of haplogroup R1b”. At some point in time, the Arbins began migration to the west, across Central Asia, North Kazakhstan, South Urals, to the Russian Plain where they have established a number of archaeological cultures between 12,000 and 4500 ybp (include- ing apparently Seroglazovo, Khvalyn, Samaran, Middle Volga, Drevneyamnay, Catacomb, and also “P urgan” cultures which are largely considered as controversial and not accepted by many historians; it should be emphasized that all those above suggestions regarding the archaeological cultures can be viewed at present only as very tentative ones). They migrated southward, in part (leaving their R1b haplo- group and the respective haplotypes behind), over the Caucasus to Anatolia around 6000 ybp; to the rest of Asia Minor, and to the Middle East. The Arbins have apparently established the Sumer cu several routes, carrying mainly the R-M269 subclade and its downstream L23 subclade. One route is the northern route, from the Russian Plain to the west, ~4600 - 4400 ybp; another, concurrently along Asia Minor and the Middle East westward with the same two subclades; and yet another which will popu- late Europe the most, migrating along North Africa-Mediter- ranean Sea via ancient Egypt to the Pyrenees, to arrive ~4800 ybp. On this route the R1b-V88 tribe split off and went south, eventually to Central Africa (mainly Cameroon and Chad judg- ing by their present-day distribution), where a common ances- tor of the current R1b-V88 haplotype lived ~4400 ybp. At the arrival time to Iberia ~4800 ybp, the M269 subclade split off M51 and soon thereafter L11 and its downstream sub- clades. They became the Bell Beakers and moved north along with newly arisen subclades P312 and L21, the latter within a few centuries after P312. Those subclades and their downstream clades have effectively without major interruptions populated Europe from the Atlantic to the Balkans, Carpathian Mountains, present day Poland, the western border of the Russian Plain, to the Baltic Sea evidenced by the smooth haplotype trees as wit- ness to non-stop proliferation of R1b haplotypes. The Isles had a different history of their R1b haplotypes and lineages. The bearers of L11, P312 and L21 moved to the Isles by land and sea concurrently with those Arbins who were populating Europe between 4000 and 2500 ybp, and formed the respective “local” subclades, such as P314, M222, L226, which rgely populated the Isles. As a result, a significant part of the Isles is populated almost exclusively by the Arbins whose fre- quency reaches 92% - 96% among the population. In general, the frequency of the Arbins in Western and central Europe reaches—albeit not uniformly—some 60% of the population. This study essentially presents an example of the application of DNA Genealogy for studying the history of mankind. This example is a complex and challenging endeavor which also touches upon some mysterious puzzles of history and linguis- tics. One of those puzzles is what language or lan oken by the Arbins from 16,000 to 3000 ybp and it almost certainly was a continuing, in its dynamics, a non-Indo-Euro- pean language. Assumptively, these languages are considered by linguists as assorted and disconnected “dead” and not-so dead languages, such as proto-Turkic, Sumer, North-Caucasian, Dene-Caucasian, Basque, and many pre-IndoEuropean lan- guages in Europe of 5000 - 2000 ybp, some later. The language of the Arbins may have been originally one language easily flowing through millennia and across Eurasia. However, this is a subject of another study. Acknowledgements The author is indebted to Susan Hedeen for her valuable help with the preparation of the manuscript. REFERENCES Balanovsky, O., Dibirova, K., Dybo, A., Mudrak, O., Frolova, S., Pocheshkhova, E. et al. (2012). Parallel evolution of genes and languages in the Caucasus region. Molecular Biology and Evolution, 29, 359-365. Balaresque, P., Bowden, G. R., Adams, S. M., Leung, H.-Y., King, T. E., Rosser, Z. H. et al. (2010). A predominantly Neolithic origin for European paternal lineages. PL oS Biology, 8, 1000285 Barac, L., Pericic, M., Klaric, I omosome STRs in Croatians. 127-133. ., Pericic, M., Klaric, I. M., Rootsi, S., Janicijevic, B., Kivisild, T. et al. (2003b). Y chromosomal heritage of Croatian population and its island isolates. European Journal of H Bengtson, J. D. (1997)an: A Dene-Caucasian language? Mother To. The riddle of Sumeri ngue, 3, 63-74. Capelli, C., Brisighelli, F., Scarnicci, F., Arredi, B., Caglia, A., Vetrugno, G. et al. (2007). Y chromosome genetic variation in the Italian pen- insula is clinal and supports an admixture model for the Meso- lithic- Neolithic encounter. Molecular Phylogenetics and Evolution, 44, 228-239. doi:10.1016/j.ympev.2006.11.030 Cardoso, J. L. (2001). Le phenomene campaniformedans les basses Copyright © 2012 SciRes. 103  A. A. KLYOSOV vallees du Tage et du Sado (Portugal). In F. Nicolis (Ed.), Bell Beak- ers Today, Trento: Ufficio Beni Archeologici. Cardoso, J. L., & Soares, A. M. (1990). Chronologia absoluta para o campaniforme da Estremadura e do Sudoeste de Portugal. O Ar- queologo Portugues, 8-10, 203-228. Cinnioglu, C., King, R., Kivisild, T., Kalfoglu, E., Atasoy, S., Cavalleri, G. L. et al. (2004). Excavating Y-chromosome haplotype strata In Anatolia. Human Genetics, 114, 127-148. doi:10.1007/s00439-003-1031-4 Contu, D., Morelli, L., Santoni, F., Foster, J. W., Francalacci, P., Cucca, F. (2008). Y-Chromosome based evidence for pre-Neolithic origin of the genetically homogeneous but diverse Sardinian population: in- ference for association scans. PLoS ONE, 3. doi:10.1371/journal.pone ruciani, F., Trombetta, B., Sellitto, D., Massaia, A., Destro-Bisol, G., Watson, E. et al. (2010). Human Y chromosome haplogroup R-V88: C A paternal genetic record of early mid Holocene trans-Saharan con- nections and the spread of Chadic languages. European Journal of Human Genetics, 18, 800-807. doi:10.1038/eihg.2009.231 Di Gaetano, C., Cerutti, N., Crobu, F., Robino, C., Inturri, S., Gino, S. et al. (2009). Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome. European Journal of H um an Ge ne ti cs , 17, 91-99. doi:10.1038/ejhg.2008.120 mmer, M. F., Behar, D. M., Karafet, T. M., Mendez, F. L., Hallmark, B., Erez, T. et al. (2009). Extended Y chr Ha omosome haplotypes re- solve multiple and unique lineages of the Jewish priesthood. Human Genetics, 126, 707-717. doi:10.1007/s00439-009-0727-5 errera, K. J., Lowery, R. K., Hadden, L., Calderon, S., Chiou, C., Ye- piskoposyan, L. et al. (2011). Neolithic patrineal signals indicate that the Armenian plateau was repopulated by agr H iculturalists. European Journal of Human Genetics, 20, 313-320. doi:10.1038/ejhg.2011.192 ang, L., Lu, Y., Wang, C., Hu, K., Chen, F., Liu, K. et al. (2011). Y- chromosome O3 haplogroup diversity in Sino-Tibetan populations reveals two migration routes into the Eastern Himalayas. Annals of K Human Genetics, 76, 92-99. doi:10.1111/j.1469-1809.2011.00690 Klyosov, A. A. (2008a). Mysteries of the “Western European” hap- logroup R1b. Proceedings of the Russian Academy of DNA Geneal- ogy, 1, 568-630 (in Russian). lyosov, A. A. (2008b). Origin of the Jews via DNAK genealogy. Pro- ceedings of the Russian Academy o f DNA Genealogy, 1, 54-232. Klyosov, A. A. (2009a). A comment on the paper: Extended Y chromo- some haplotypes resolve multiple and unique lineages of the Jewish priesthood. Human Genetics, 126, 719-724. doi:10.1007/s00439-009-0739-1 Klyosov, A. A. (2009b). Once again on the “population mutation rate” of L. Zhivotovsky, or how fables are born. Proceedings of the Rus- sian Academy of DNA Genealo gy , 2, 1162-1181 (in Russian). lyosov, A. A. (2009c). DNA Genealogy, mKutation rates, and some K K in Russian). K K2-P312 and related sub- K. Haplogroups and haplotypes in Armenia (hap- Kut of Africa” theory re-examined. Proceed- in Anthropology, 2, 1- historical evidences written in Y-chromosome. I. Basic principles and the method. Journal of Genetic Geneal ogy, 5, 186-216. lyosov, A. A. (2009d). DNA Genealogy, mutation rates, and some historical evidences written in Y-chromosome. II. Walking the map. Journal of Genetic Genealogy, 5, 217-256. Klyosov, A. A. (2010a). Haplogroup R1b1 and its subclades in Asia. Proceedings of the Russian Academy of DNA Genealogy, 3, 1676- 1695 (in Russian). Klyosov, A. A. (2010b). Haplotypes of haplogroup R1b in central Af- rica. Proceedings of the Russian Academy of DNA Genealogy, 3, 369-378 (in Russian). lyosov, A. A. (2010c). Haplogroup R1b. Part 2. Proceedings of the Russian Academy of DNA Genealogy, 3, 406-475 ( Klyosov, A. A. (2010d). The “age” of subclade R1b1a2-M269 and its subclades (L23, L51, L11). Proceedings of the Russian Academy of DNA Genealogy, 3, 1310-1315 (in Russian). Klyosov, A. A. (2010e). Irish haplotypes and haplogroups. Proceedings of the Russian Academy of DN A G e nealogy, 3, 1029-1053. lyosov, A. A. (2011a). Origin of ancient subclades of haplogroup R1b—Regions and times. Proceedings of the Russian Academy of DNA Genealogy, 4, 2227-2245 (in Russian). lyosov, A. A. (2011b). Haplotypes of R1b1a clades: Origin and “ages” of most recent common ancestors. Pro- ceedings of the Russian Academy o f DNA Genealogy, 4, 1127-1195. lyosov, A. A. (2011c) logroups J2, R1b-L23, R1b-M269, and T-M184). Proceedings of the Russian Academy of DNA Genealogy, 4, 1985-1993 (in Russian). lyosov, A. A. (2011d). “O ings of the Russian Academy of DNA Genealogy, 4, 1908-1977 (in Russian). Klyosov, A. A., & Rozhanskii, I. L. (2012a). Haplogroup R1a as the Proto-Endo-Europeans and the legendary Aryans as witnessed by the DNA of their current descendants. Advances 13. doi:10.4236/aa.2012.21001 lyosov, A. A., & Rozhanskii, I. L. (2012b). Re-examining theK “out of Kr ory, culture, and char- L L M (1989). In search of the indo-Europeans: Language, M y subtyping the Y chromosome hap- M Iberica y NE, 5, e10419. africa” theory and the origin of Europeoids (Caucasoids) in light of DNA genealogy. Advances in Anthropo lo gy, 2, in press. amer, S. N. (1971). The Sumerians: Their hist acter. Chicago, IL: University of Chicago Press. ashgary, Z., Khodadadi, A., Singh, Y., Houshmand, S. M., Mahjoubi, F., Sharma, P. et al. (2011). Y chromosome diversity among the Ira- nian religious groups: A reservoir of genetic variations. Annals of Human Biology, 38, 364-372. obov, A. S. (2009). Structure of the gene pool of the Bashkir subpopu- lations. Ufa. http://ftp.anrb.ru/molgen/Lobov_AS.PDF allory, J. P. archaeology and myth. London: Thames and Hudson. alyarchuk, B., Derenko, M., Denisova, G., Maksimov, A., Wozniak, M., Grzybowski, T. et al. (2011). Ancient links between Siberians and native Americans revealed b logroup Q1a. Journal o f Hu m a n G e n e ti c s , 56, 583-588. artinez, P. V., Lull, V., & Mico, R. (1996). El vaso campaniforme, in : Cronologia de la Prehistoria Reciente de la Peninsula Baleares (c. 2800-900 cal. A.N.E.). British Archaeological Reports, International Series 652, 105-110. Morelli, L., Contu, D., Santoni, F., Whalen, M., Francalacci, P., & Cucca, F. (2010). A comparison of Y-chromosome variation in Sar- dinia and Anatolia is more consistent with cultural rather than demic diffusion of agriculture. PLoS O doi:10.1371/journal.pone.0010419 European Bell Beakers and the consequen Muller, J., & Van Willigen, S. (2001). New radiocarbon evidence for ces for the diffusion of the P n Europe traces major episodes Bell Beaker phenomenon. In F. Nicolis (Ed.), Bell Beakers Today, Trento: Ufficio Beni Archeologici. Myres, N. M., Rootsi, S., Lin, A. A., Jarve, M., King, R. J., Kutuev, I. et al. (2010). A major Y-chromosome haplogroup R1b holocene era founder effect in central and western Europe. European Journal of Human Genetics, 19, 95-101. Nocete, F. (2006). The first specialized copper industry in the Iberian Peninsula: Cabezo Jure (2900-2200 BC). Antiquity, 80, 646-657. ericic, M., Lauc, L. B., Klaric, A. M. et al. (2005). High-resolution phylogenetic analysis of southeaster of paternal gene flow among Slavic populations. Molecular Biology and Evolution, 22, 1964-1975. doi:10.1093/molbev/msi185 oewer L., Willuweit S., Kruger C., Nagy M., Rychkov S., Morozowa I. et al. (2008). Analysis of Y chromosome STR hap European part of Russia reveals hi R lotypes in the gh diversities but non-significant genetic distances between populations. International Journal of Le- gal Medicine, 122, 219-223. doi:10.1007/s00414-007-0222-2 ozhanskii, I. L., & Klyosov, A. A. (20R11). Mutation rate constants in DNA genealogy (Y chromosome). Advances in Anthropology, 1, 26- 34. doi:10.4236/aa.2011.12005 emino, O., Passarino, G., Oefner, P. J., Lin, A. A., Arbuzova, S., Beckman, L. E. et al. (2000). Th S e genetic legacy of paleolithic Homo sapiens sapiens in extant Europeans: A Y chromosome perspective. Science, 290, 1155-1159. doi:10.1126/science.290.5494.1155 Underhill, P. A., Myres, N. M., Rootsi, S., Metspalu, M., Zhivotovsky, M. A., King, R. J. et al. (2009). Separating the post-Glacial coances- Copyright © 2012 SciRes. 104  A. A. KLYOSOV Copyright © 2012 SciRes. 105 9 try of European and Asian Y chromosomes within haplogroup R1a. European Journal of Human Ge net ics , 18, 479-484. Weale, M. E., Yepiskoposyan, L., Jager, R. F., Hovhannisyan, N., Khu- doyan, A., Barbage-Hall, O. et al. (2001). Armenian Y chromosome haplotypes reveal strong regional structure within a single ethnona- tional group. Human Genetics, 109, 659-674. doi:10.1007/s00439-001-0627- Wakiev, R., Underhill, P. A., Evseeva, 0244-10249. ells, R. S., Yuldasheva, N., Ruzib I., Blue-Smith, J. et al (2001). The Eurasian heartland: a continental perspective on Y-chromosome diversity. Proceedings of the National Academy of Sciences USA, 98, 1 doi:10.1073/pnas.171305098 iik, K. (2008). Where did European men come from? Journal of Genetic Genealogy, 4, 35-85. akharov, S. V. (2010). On the origin of Botai culture. Vestnik W Zof Ar- ography, 1, 49-58. chaeology, Anthropology, and Ethn Zalloua, P. A., Xue, Y., Khalife, J., Makhoul, N., Debiane, L., Platt, D. E. et al. (2008). Y-Chromosomal diversity in Lebanon is structured by recent historical events. American Journal of Human Genetics, 82, 973-882. doi:10.1016/j.ajhg.2008.01.020 hivotovsky, L. A., Underhill, P.Z A., Cinnioglu, C., Kayser, M., Morar, application to human population B., Kivisild, T. et al. (2004). The effective mutation rate at Y-chro- mosome short tandem repeats, with divergence time. American Journal of Human Genetics, 74, 50-61. doi:10.1086/380911 Zhong, H., Shi, H., Qi, X.-B., Duan, Z.-Y., Tan, P.-P., Jin, L. et al. (2010). Extended Y-chromosome investigation suggests post-Glacial migrations of modern humans into East Asia via the northern route. Molecular Biology and Ev o luti on, 28, 717-727. PPENDIX The following DNA projects were selected as primary hap- type databases: http://www.familytreedna.com/public/R1b1b1/default.aspx?s ction=yresults http://www.familytreedna.com/public/ht35new/default.aspx treedna.com/public/AssyrianHeritageDNA px?section=yresults http://www.familytreedna.c om/public/IrelandHeritage/defaul t.aspx?section=yresults http://www.familytreedna.com/public/R-L21/de fault.aspx? se ction=yresults Reference data were selected according to SNP assignment from YSearch database: (http://www.ysearch.org) and public projects of FTDNA (http://www.familytreedna.com) A lo e http://www.family Project/default.as
|