American Journal of Plant Sciences
Vol.08 No.06(2017), Article ID:76521,15 pages
10.4236/ajps.2017.86097
Essential Oils from Mentha viridis (L). L. and Mentha pulegium L.: Cytogenotoxic Effects on Human Cells
Lucilene Fernandes Silva1, Maria das Graças Cardoso1*, Paulo Sérgio Castilho Preté1, Maria Luisa Teixeira1, David Lee Nelson2, Maísa Lamounier Magalhães3, Vanuzia Rodrigues Fernandes Ferreira1, Rafaela Vieira Souza1, Luana Isac Soares1, Silvana Marcussi1
1Departamento de Química, Universidade Federal de Lavras, Campus Universitário, Lavras, Minas Gerais, Brasil
2Departamento de Química, Universidade Federal dos Vales do Jequitinhonha e Mucuri, Campus Universitário, Diamantina, Minas Gerais, Brasil
3Departamento de Ciência dos Alimentos, Universidade Federal de Lavras, Campus Universitário, Lavras, Minas Gerais, Brasil

Copyright © 2017 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: March 29, 2017; Accepted: May 23, 2017; Published: May 26, 2017
ABSTRACT
The essential oils from Mentha viridis (L). L. and Mentha pulegium L. were studied to assess their inhibitory potential on phospholipase from snake venoms and to determine their cytogenotoxic action on human cells. These essential oils were able to inhibit the breakdown of phospholipids induced by venoms of snakes of the Bothrops genus. Both oils presented hemolytic activity, although the Mentha viridis (L). L. oil induced hemolysis only at the highest concentrations (14.6 and 29 µL∙mL−1). The essential oil from M. viridis induced 3.9; 8.6 and 16.2 times greater damage to human leukocyte DNA than that observed with the positive control (100 µg∙µL−1 doxorubicin) at concentrations of 0.25; 0.5 and 1.0 µL∙mL−1, respectively. A similar effect was observed for the oil from M. pulegium (2.1, 2.5 and 15.8 times greater damage). The results extend the characterization of these essential oils and demonstrate their potential use in industries.
Keywords:
Essential Oils, Mentha viridis (L). L., Mentha pulegium L., Hemolysis, Comet Assay

1. Introduction
Among the many compounds produced by plants, essential oils are volatile materials contained in many plant organs that are related to various functions necessary for plant survival [1] [2] . Among the families that concentrate their volatile constituents in the leaves, there is the Lamiaceae family, which has many plants that occur spontaneously among the flora. Mentha viridis (L). L. and Mentha pulegium L. belong to this family and are important sources of biologically active compounds [3] [4] .
Although disclosure of the use of natural products has grown in recent years as described by Bors et al. [5] , the use of essential oils for the preparation of medicines or as models for developing new drugs must be intimately related to extensive characterization of the active principles, the appropriate dosages and the forms of administration, as well as the associated adverse effects. Guimarães et al. [6] reported the great effort of the scientific community in validating the use of plants to treat snakebites. In this work, 87 compounds and plant extracts with the potential for protecting against snakebites were grouped. In addition, natural compounds can be used to target other diseases because they exhibit potential inhibitory activity against enzyme classes widely distributed in animal organisms with essential physiological functions, for example, phospholipases and proteases.
Yamaguchi and Veiga-Junior [7] , for example, reported the absence of hemolytic effects of essential oils obtained from leaves and branches of Endlicheria citriodora (1.22 to 625 μg∙mL−1). However, there are a few studies on genotoxicity evaluations, the effect on phospholipase or even potential anti-snakebite activities of the essential oils or their constituents, highlighting a wide range of studies to be explored [8] [9] [10] .
The Mentha, are widely used by humans, with emphasis on the use of essential oils in cosmetics, pharmaceuticals, food, confectionery and industrial alcoholic beverages [11] . The research related to phospholipase, hemolytic and genotoxic activities of natural products with the perspective of use is a priority measure because the pharmacological and toxic effects are related to the form of administration and the dose may either cure or be harmful to humans [12] . The aim of this study was to determine the toxicity and pharmacological potential of the essential oils from M. viridis and M. pulegium through induction trials and inhibition of hemolysis and phospholipid degradation, as well as to evaluate the genotoxic activity.
2. Material and Methods
2.1. Plant Material and Essential Oil Isolation
The leaves (rib and limbs) from adult plants of Mentha viridis (L.) L. and Mentha pulegium (L.) species were collected around 07:00 hours on the Campus of the Universidade Federal de Lavras (UFLA) (21˚14'S, longitude 45˚00'W Gr. and 918 m altitude) on days without precipitation in the month of November 2012. Species identification was kindly performed by Dra. Mariana Esteves Mansanares, Department of Biology of UFLA, and exsiccates were deposited in the ESAL Herbarium at UFLA under the registration numbers 27.123 and 27.122, respectively.
The essential oils from fresh leaves were extracted by hydro-distillation using a modified Clevenger apparatus adapted to a 6-L, round-bottom flask over a period of 2 hours. The hydrosols were centrifuged (Fanem Baby I Modelo 206 BL) for 10 minutes at 965g at room temperature, and the oils were packaged in amber glass bottles and stored at a temperature of 4˚C [13] .
2.2. Human Blood for Comet and Hemolysis Assays
The peripheral blood from volunteers aged 21 to 40 years was used after free informed consent; the volunteers did not present disease symptoms and reported not having used prescription drugs for at least 30 days prior to collection. These subjects were randomly selected. This study was approved by the Committee of Ethics in Research with Humans of UFLA and filed under the number 17935713.8.0000.5148.
2.3. Comet Assay for Genotoxicity Evaluation
The concentrations chosen for the evaluation of the essential oils correspond to previously determined non-cytotoxic doses (data not shown). Negative (without treatment) and positive (100 µg∙µL−1 doxorubicin) controls were conducted simultaneously. The blood samples (500 µL) were incubated in an oven at 37˚C in for 4 hours the presence of the treatments. The Comet assay was then performed according to the method described by Singh et al. [14] . All the procedures were performed in the dark.
Nucleoids and electrophoresis: An aliquot (20 μL) of each cell suspension containing the treatments was mixed with 100 μL of low-melting-point agarose solution (0.5%wv-1 in Phosphate Buffered Saline-PBS), applied to a microscope slide previously coated with standard normal-melting-point agarose solution (1% w v−1 in PBS), immediately overlaid with a coverslip, and held at 4˚C for 10 minutes. For each treatment/volunteer, three slides were prepared. The coverslips were removed and the slides were immersed in lysis solution (2.5 mol∙L−1 NaCl, 100 mmol∙L−1 EDTA, 10 mmol∙L−1 Tris, 1% Triton X-100, 10% DMSO; pH 10), where they remained for 20 hours at 4˚C to isolate their nucleoids for analysis.
After lysis, the slides were kept in a freshly prepared electrophoresis solution (1 mmol∙L−1 EDTA, 300 mmol∙L−1 NaOH; pH 13) at approximately 15˚C for 25 minutes to expose the alkali-labile sites and then subjected to electrophoresis at 25V for 30 minutes. The slides were kept in a neutralization solution (0.4 mol∙L−1 Tris; pH 7.5) for 25 minutes, dried and fixed with absolute ethanol.
Staining and analysis: The slides were stained with propidium iodide solution (0.5 mg.mL−1), overlaid with a coverslip and analyzed with an epifluorescence microscope (Nikon ECLIPSE E400) at 400× magnifications. To measure damage levels in lymphocyte DNA molecules, 100 nucleoids of each slide and three slides per treatment/volunteer (totalling 300 nucleoids per treatment/volunteer), were counted and classified by the same evaluator using visual score patterns.
The nucleoids were classified according to Collins et al. [15] and adaptations of Marcussi et al. [16] into Class 0: no damage (damage < 5%); Class 1: low damage level (5% - 20%); Class 2: intermediate damage level (20% - 40%); Class 3: high damage (40% - 95%); Class 4: totally damaged (damage > 95%). The average frequency of damage was calculated from the sum of the percentages of damage of 1, 2, 3 and 4. The arbitrary units (0 - 400; where 0 = no damage and 400 = 100% damage) were calculated by the equation (1 × damage in class 1) + (2 × damage in class 2) + (3 × damage in class 3) + (4 × damage in class 4), as described by Collins et al. [17] .
2.4. Hemolytic Activity in Liquid Medium
The blood collected (10 mL) in the presence of an anticlotting agent was immediately mixed with saline solution (2 mM NaH2PO4; 3 mM Na2HPO4; 154 mM NaCl; pH 7.4) and centrifuged at 700g (Fanem Baby® IModelo 206 BL) for 5 minutes. The plasma was removed, and the red cells were suspended in 5 mmol∙L−1 phosphate buffer, pH 7.4 and centrifuged under the same conditions. This washing procedure was repeated three times at 4˚C. The 100% red blood cell concentrate was diluted to 2% and 0.15% hematocrits (137.33 and 13 µmol∙L−1 lipid, respectively), using the same buffer [18] .
The hemolytic activity was evaluated by incubating 1 mL of erythrocyte suspension (2% or 0.15%) for 60 minutes at 37˚C with the oil solutions at the concentrations of 0.6; 1.0; 2.0; 3.0; 4.0; 5.0 and 10.0 µL∙mL−1, followed by centrifugation at 1500g for 5 minutes. The hemoglobin concentration was determined in the supernatant by measuring the absorbance at 412 nm (Shimadzu UV-160 1 PC) according to Rangel et al. [18] , with the modifications of Preté et al. [19] . The controls were performed using an erythrocyte suspension in PBS (c1 = mechanical hemolysis control) and distilled water (c2 = total hemolysis control). The hemoglobin concentration was determined using the equation:
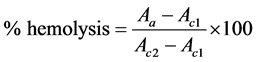
in which Aa, Ac1 and Ac2 are, respectively, the absorbance of the sample and of the controls c1 and c2 at 412 nm.
2.5. Hemolytic Activity in Solid Medium
The hemolysis of human erythrocytes in solid medium was performed according to the method described by Gutiérrez et al. [20] , with the addition of erythrocytes to replace the egg yolk. A gel was prepared (0.01 mol∙L−1 CaCl2; erythrocytes at hematocrit 100%; diluted 1:3 v v−1 in PBS; 0.005% sodium azide) with 1% agarose, pH 7.2. After solidification, holes were made (~0.5 cm diameter), and the oil solutions (5.5; 10.9; 21.8; 43.6 and 87.3 µL∙mL−1) were applied at a final volume of 25 µL. The gels containing the samples were kept in a cell culture chamber at 37˚C for 24 hours, and the formation of a translucent halo around the hole in the gel characterized hemolytic activity, which was quantified by measuring the diameter of the halo [19] . The assays were performed in triplicate. The inhibition of the snake-venom-induced hemolysis by the essential oils was also evaluated. Solutions of venoms from Bothrops moojeni and Lachesis muta (40 μg∙mL−1) were previously incubated with 5.5; 10.9 and 21.8 μL∙mL−1 of the oils at 37˚C for 1 hour.
2.6. Phospholipase Activity
The phospholipase activity was evaluated by the use of solid medium, as described by Gutiérrez et al. [20] . A gel was prepared (0.01 mol∙L−1 CaCl2; egg yolk diluted 1:3 v v−1 in PBS; 0.005% sodium azide) with 1% agarose, pH 7.2, and, after solidification, holes were made (~ 0.5 cm diameter) and the oil solutions (5.5; 10.9; 21.8; 43.6 and 87.3 µL∙mL−1) were applied in a final volume of 25 µL.
The inhibition of the snake-venom-induced phospholipase activity by the essential oils was also evaluated. Solutions of venom from B. jararaca, B. jararacussu, B. moojeni and B. alternatus (40 μg∙mL−1) were incubated with 5.5; 10.9; 21.8; 43.6 and 87.3 μL∙mL−1 concentrations of the oils at 37˚C for one hour. The gels containing the samples were kept in a cell culture chamber at 37˚C for 24 hours, and the formation of a translucent halo around the hole in the gel characterized phospholipase activity, which was quantified by measuring the halo diameter [21] . The assays were performed in triplicate.
2.7. Statistical Analysis
The results of the Comet assay were statistically evaluated by analysis of variance, and the means were compared using the Scott-Knott test (p < 0.05) with the aid of the R software (R Development Core Team, 2011) [22] . For this test, Arbitrary Units were also calculated as described by Collins et al. [17] . For the other assays, data were treated as means and standard deviation of triplicates.
3. Results and Discussion
3.1. Comet Assay
The essential oils from M. viridis and M. pulegium induced DNA fragmentation at all the concentrations evaluated. The highest levels of fragmentation (classes 3 and 4) were observed for M. viridis in all treatments evaluated and for M. pulegium at the concentration of 1 µL∙mL−1.
The arbitrary unit (A.U.) values for the M. viridis treatments were 86.2; 190.0 and 358.4 for doses of 0.25; 0.5 and 1.0 µL∙mL−1, respectively. These values are 3.9; 8.6 and 16.3 times higher than those observed for the negative control (22.0 A.U.), which demonstrates the genotoxic potential of the oils. Treatment with the same concentrations of M. pulegium resulted in A.U. values of 46.6; 55.2 and 349.6, these being 2.1, 2.5 and 15.8 times higher, respectively, than the negative control.
The essential oils from M. viridis and M. pulegium presented A.U. values of 358.4 and 349.6, respectively, when a 1.0 µL∙mL−1 concentration was administered. These values were higher than those observed for the antitumor drug used as a control (159.0), even though this drug was evaluated at a concentration 10 times higher (100 µg∙µL−1) (Table 1).
The essential oils from M. viridis and M. pulegium, at a concentration of 1 μL∙mL−1, induced percentages of damaged nucleoids (94.2 and 97.0, respectively) higher than that observed with doxorubicin (84.8%).
The major constituents found in the essential oil from Mentha viridis were linalool (40.70%), carvone (13.52%) and α-terpinene (8.56%). The constituents found in the essential oil from Mentha pulegium were pulegone (50.01%), menthol (31.90%) and menthone (16.56%). The highest percentages of totally damaged nucleoids (class 4: damage > 85%) were 16.4; 81.6 and 72.0, respectively, at concentrations of 0.5 and 1.0 µL∙mL−1 for M. viridis and 1.0 µL∙mL−1 for M. pulegium.
With the use of the Comet assay, Péres et al. [23] evaluated the effect of the essential oil from Piper gaudichaudianum, which contains (E)-nerolidol, a-hu- mulene, (E)-caryophyllene, and bicyclogermacrene as the majority sesquiterpene compounds, on the DNA from V79 cells in culture. The authors found a significant increase in the damage index corresponding to concentrations of 0.1; 1.0; 2.0; 4.0; 6.0 and 10.0 mg∙L−1.
The major constituents present in the oil evaluated by Peres et al. [23] differ from those found in this study for the Mentha oils. Although they belong to the same class of substances, these compounds may have distinct biological activities because of differences in physiological functions, enantiomeric differences and other factors inherent to the structures of the constituents. The cell type used by those authors (V79, lung fibroblasts) also differs from that assessed in this study (peripheral blood leukocytes) and is one of the factors that led to the different effects observed.

Table 1. Average nucleoid number per class of Comet, frequency of nucleoids with damage and arbitrary units after the treatment with essential oils.
1Data represent the mean values obtained for 300 nucleoids/treatment/volunteer. Five volunteers were used (1 volunteer/experiment); 2Frequency of damage: sum of the damage in classes 1 to 4; 3Arbitrary units: (1 × damage in class 1) + (2 × damage in class 2) + (3 × damage in class 3) + (4 × damage in class 4), according to the method described by Collins et al. (2004) 15; 4Positive control: 100 µg∙µL−1 doxorubicin. The results with the same letters in the columns do not differ by the Scott-Knott test (p < 0.05) in comparison with the controls.
Rassouli et al. [24] investigated the in vitro cytotoxic and apoptotic effects of stilosin monoterpenoid, extracted from the Ferula ovina plant species, on cancer cells and found that 37 mg∙L−1 stilosin induced DNA damage in about 65% of the cells treated. In the same context, Silva and Guterres [25] demonstrated the antigenotoxic potential of arjunolic acid using doxorubicin hydrochloride to induce damage to Drosophila melanogaster wing cells.
Despite major advances in cancer therapy, research for new compounds, mainly of natural origin, with potential anticancer and fewer side effects are of great value for the development of new therapies. The results of genotoxicity tests for the essential oils from M. viridis and M. pulegium, whose major constituents are represented by terpenes, emphasize the anticancer potential of these oils when compared to the allopathic drug doxorubicin.
3.2. Hemolytic Activities
The essential oil from M. pulegium presented an increase in hemolytic activity starting at a concentration of 5.5 μL∙mL−1 and ending at 87.3 μL∙mL−1, considering limitations of the volumes implicit to each method employed. Activity was observed only at the highest concentrations (43.6 and 87.3 μL∙mL−1) of the oil from M. viridis (Figure 1). In contrast, only the oil from M. viridis showed inhibitory effects against hemolysis induced by snake venoms in a solid medium. A 7.7% inhibition of the activity of L. muta venom at a concentration of 5.5 μL∙mL−1 and 6.7% inhibition of B. moojeni activity at concentrations of 5.5; 10.9 and 21.8 μL∙mL−1 (Figure 2) were observed.
During tests of inhibition of L. muta venom, a greater activity was observed for the essential oil from M. viridis at a concentration of 21.8 μL∙mL−1 (Figure 2).
Nearly 100% hemolytic activity was observed for all the concentrations (0.6 to 10 µL∙mL−1) of the two essential oils tested in liquid medium at 0.15% hematocrit

Figure 1. Hemolytic effect of essential oils on human peripheral blood erythrocytes measured on solid medium. Different volumes of oils were applied to medium containing erythrocytes where they remained for 24 hours at 37˚C. Values represent averages of triplicate analyses and the standard deviation.

Figure 2. Effect of the essential oil from Mentha viridis on the hemolytic activity induced by the venoms from Lachesis muta and Bothrops moojeni, assessed in solid medium. Venoms and oils in different proportions were incubated at 37˚C for a period of 1 hour and then applied to the medium containing the cells and incubated for 24 hours at the same temperature. The average of triplicate analyses was converted to percentage of activity, considering the controls (only venom of L. muta and B. moojeni) as 100%.
(Figure 3(a)). The increase in the hematocrit to 2% resulted in a significant reduction in the activity of the M. viridis essential oil to 6% at a concentration of 0.6 µL∙mL−1, whereas 83% activity was observed with 1 µL∙mL−1 of the same oil (Figure 3(b)). The induction or inhibition of toxic drug effects is dependent on the dose of the compounds evaluated, and it appears that the concentration of 0.6 µL∙mL−1 of M. viridis oil corresponds to the turning point wherein the number of active molecules present fail to induce hemolysis and can possibly inhibit it.
Constituents present in oils may interact with components of erythrocyte membranes, leading to destabilization of its structure and a disorderly influx of ions and water that result in rupture of the membranes. Thus, the reduction in the number of erythrocytes in solution (0.15% hematocrit) probably resulted in interactions between membrane structures and constituents present in oils and increased the hemolytic effects observed for both oils. However, a smaller number of such molecules present in the oils in the presence of a larger number of cells (2% hematocrit) can promote interactions with proteins and membrane lipids without altering their conformations, resulting in a protective effect against the action of other molecules such as enzymes with phospholipase or proteolytic activity present in venoms. The human organism has a hematocrit of approximately 45%, much higher than those used during the tests. In addition, the erythrocytes are not isolated, but rather dispersed among several other cell types, proteins, lipids, carbohydrates, ions etc., which could lead to unpredictable effects for the essential oils evaluated in this study.
Marya et al. [26] did not observe a hemolytic effect at a concentration of 0.01% in the samples when evaluating the hemolytic activity of essential oil of
 (a)
(a) (b)
(b)
Figure 3. (a) and (b) Hemolytic effect of essential oils on human peripheral blood erythrocytes evaluated in liquid medium. Different concentrations of the oils were incubated with erythrocyte solutions at 37˚C for a period of 60 minutes. (a) Evaluation of hemolysis in 0.15% hematocrit; (b) Evaluation of hemolysis in 2% hematocrit. Values represent means of triplicate analyses and the standard deviation.
cloves and the principal constituents, eugenol and eugenyl acetate, in human erythrocytes (10% hematocrit). At the highest concentration tested (0.2% v/v), eugenyl acetate, eugenol and the essential oil of cloves induced 57; 40 and 48% hemolysis, respectively. The authors showed that an increase in the concentration of the essential oil and the major constituents resulted in a higher hemolytic activity similar to the effect observed in this study. The major constituents of the essential oils of M. pulegium and M. viridis are sesquiterpenes, and those in the essential oil of cloves are phenylpropanoids. However, both essential oils were effective in inducing high hemolytic activity.
Quintans et al. [27] evaluated the effect of essential oil from Xylopia laevigata on rat erythrocytes (2% hematocrit) and observed hemolysis at concentrations ranging from 31.25 to 500 μg∙mL−1. The results observed by these authors differ from those obtained in this work for the Mentha essential oils, which presented a high hemolytic activity even when evaluated at low concentrations. Quintans et al. [27] identified γ-muruleno, δ-cadineno, germacrene-B, α-co- paene, bicyclogermacrene and E-caryophyllene (all sesquiterpenes) as the principal constituents. The essential oils from Mentha species contained oxygenated sesquiterpenes as the principal constituents, differentiating the compositions of the oils and, consequently, the activities.
Misharina et al. [28] studied the effects of the ingestion of essential oils by rats subjected to a diet containing 150 ng of Origanum vulgare L. (oregano) and Caryophyllus aromaticus L. (clove) oils. The authors collected blood from the treated rats and the untreated controls, subjected them to the same addition of water with subsequent evaluation of hemolysis. In the controls, 100% hemolysis in the blood was observed, whereas 85% and 66.3% hemolysis was observed in those animals treated with the essential oils from oregano and cloves, respectively.
3.3. Phospholipase Activity
A 20.6% inhibition of the phospholipase activity of B. jararaca (Figure 4(b)), was observed in the presence of 5.5; 21.8 and 10.9 μL∙mL−1 concentrations of the essential oil from M. pulegium, whereas no effect was observed when evaluating the M. viridis oil (Figure 4(a)). A low inhibitory activity of approximately 4.1% was observed against the venom from B. jararacussu with a concentration of 43.6 μL∙mL−1 for both oils (Figure 4(a) and Figure 4(b)). The oil from M. viridis and M. pulegium proved to be very effective in inhibiting the phospholipase activity induced by different venoms (Figure 4(a)).
We observed a 100% inhibition of phospholipase activity induced by B. alternatus in the tests with the oil from M. pulegium at concentrations of 21.8 and 43.6 μL∙mL−1. A decrease in the inhibitory effect (inhibition close to 40%) with increasing concentration of the oil (87.3 μL∙mL−1) (Figure 4(b)) was observed. The M. pulegium oil inhibited the activity induced by the B. moojeni venom in approximately 20% at concentrations of 5.5; 10.9 and 21.8 μL∙mL−1 and nearly 40% inhibition was observed with the concentrations of 43.6 and 87.3 μL∙mL−1.
Terra and Lema [29] compared the toxicity of Bothrops venoms and observed variations between the B. alternatus and B. jararaca species from different regions, as well as within the species. The authors suggested that the different levels of inhibition observed relate not only to the different constituents present in the oils, but also to differences in the compositions of the venoms.
Fernandes et al. [30] reported the inhibition of phospholipase activity of Bothrops jararacussu and Bothrops jararaca venoms by the root extract of Combretum leprosum and the triterpene arjunolic acid isolated from the extract.
 (a)
(a) (b)
(b)
Figure 4. (a) and (b) Effect of essential oils from M. pulegium and M. viridis on the phospholipase activity induced by snake venoms. Venoms and oils in different proportions were incubated at 37˚C for a period of 1 hour and then applied to the medium containing the phospholipids and incubated for 24 hours at the same temperature. (a) Evaluation of the inhibitory effect of M. viridis; (b) Inhibitory effect of M. pulegium. The average of triplicate analyses was converted to percentage of activity, considering the controls (pure venom of B. jararaca, B. jararacussu, B. moojeni and B. alternatus) as 100%.
The inhibitory effect of the extract was dose dependent, whereas the isolated compound was able to inhibit 50% of the activity of the venoms at the concentration of 300 μL∙mL−1.
The inhibitory effect of arjunolic acid is superior to those observed for M. pulegium (approximately 20.6% inhibition of the activity of B. jararaca venom and 4.0% inhibition of B. jararacussu venom) and M. viridis oils (no effect on the activity of B. jararaca venom and approximately 4.0% inhibition of B. jararacussu venom). Although, several biological activities exhibited by plant extracts or essential oils are assigned to sets of molecules that act in synergy, various activities involve isolated compounds that interact with animal enzymes.
Strauch et al. [31] reported 48%, 42% and 92% inhibition of enzyme activity of the venoms obtained from B. atrox, B. jararacussu and B. jararaca, respectively, when measured in liquid medium in the presence of 500 µg.mL−1 of crude ethanol extract of Humirianthera ampla. The high inhibitory potential reported by the authors can be the result of differences in the method of evaluation as well as a larger number of classes represented by the compounds present in the extract, such as tannins, flavonoids and terpenes, which differ from those found in the present study (only terpenes).
The inhibitory effect on phospholipase suggests a possible anti-inflammatory potential for the essential oils. The cleavage of phospholipids by phospholipase A2 present in venoms can be compared to the phospholipase activity exerted by endogenous enzymes in mammals. This specific enzymatic activity results in the breakdown of membrane phospholipids to release fatty acids and lysophospholipids, including arachidonic acid, precursor of bioactive lipids (e.g. prostaglandins, leukotrienes and thromboxanes) that operate in various physiological processes, including inflammatory and immune responses [31] [32] .
4. Conclusion
The essential oils from M. pulegium and M. viridis presented a high genotoxic potential even when evaluated at lower doses. Considering the limited options for chemotherapy available today, the oils evaluated showed promise for alternative pharmaceutical formulations. In addition, they have a potential for use as adjuvants in the treatment of snake bites and as sources of active principles for the development of anti-inflammatory or immune response modulators.
Acknowledgements
The authors acknowledge the support of the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and the Fundação de Amparo a Pesquisa do Estado de Minas Gerais (FAPEMIG) for financial support and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for a PVNS.
Conflict of Interest
The authors declare no conflict of interest.
Cite this paper
Silva, L.F., das Graças Cardoso, M., Preté, P.S.C., Teixeira, M.L., Nelson, D.L., Magalhães, M.L., Ferreira, V.R.F., Souza, R.V., Soares, L.I. and Marcussi, S. (2017) Essential Oils from Mentha viridis (L). L. and Mentha pulegium L.: Cytogenotoxic Effects on Human Cells. American Journal of Plant Sciences, 8, 1423-1437. https://doi.org/10.4236/ajps.2017.86097
References
- 1. Bakkali, F., Averbeck, S., Averbeck, D. and Idaomar, M.F. (2008) Biological Effects of Essential Oils—A Review. Food and Chemical Toxicology, 46, 446-475.
- 2. Simões, C.M.O., Schenkel, E.P., Gosmann, G., Mello, J.C.P., Mentz, L.A. and Petrovick, P.R. (2007) Pharmacognosy: From Plant to Medicine. Porto Alegre.
- 3. Dimitri, M.J. (1980) Encyclopedia Argentina of Agriculture and Gardener. Buenos Aires.
- 4. Wang, H.T., Yu, X., Liu, Y., Liang, C.Y. and Li, W.L. (2013) Analysis of Genetic Variability and Relationships among Mentha L. Using the Limonene Synthase Gene, LS. Gene, 524, 246-252.
- 5. Bors, M., Sicińska, P., Michalowicz, J., Wieteska, P., Gulewicz, K. and Bukowska, B. (2012) Evaluation of the Effect of Uncaria tomentosa Extracts on the Size and Shape of Human Erythrocytes (in Vitro). Environmental Toxicology and Pharmacology, 33, 127-134.
https://doi.org/10.1016/j.etap.2011.11.003 - 6. Guimarães, C.L., Moreira-Dill, L.S., Fernandes, R.S., Costa, T.R., Hage-Melim, L.I., Marcussi, S., Carvalho, B.M., Da Silva, S.L., Zuliani, J.P. and Fernandes, C.F. (2014) Biodiversity as a Source of Bioactive Compounds against Snakebites. Current Medicinal Chemistry, 21, 2952-2979.
https://doi.org/10.2174/09298673113206660295 - 7. Yamaguchi, K.K.L., Veiga Jr., V.F., Pedrosa, T.N., Vasconcellos, M.C. and Lima, E.S. (2013) Biological Activities of Essential Oils of Endlicheria citriodora, a Lauraceae Rich in Methyl Geranate. Química Nova, 36, 826-830.
https://doi.org/10.1590/S0100-40422013000600015 - 8. Carvalho, B.M.A., Santos, J.D.L., Xavier, B.M., Almeida, J.R., Resende, L.M., Martins, W., Marcussi, S., Marangoni, S., Stábeli, R.G. and Calderon, L.A. (2013) Snake Venom PLA2s Inhibitors Isolated from Brazilian Plants: Synthetic and Natural Molecules. BioMed Research International, 2013, Article ID: 15304.
- 9. Torres, A.M., Camargo, F.J., Ricciardi, G.A., Ricciardi, A.I. and Dellacassa, E. (2011) Neutralizing Effects of Nectandra angustifolia Extracts against Bothrops neuwiedi Snake Venom. Natural Product Communications, 6, 1393-1396.
- 10. Miranda, C.A.S.F., Cardoso, M.G., Mansanares, M.E., Gomes, M.S. and Marcussi, S. (2014) Preliminary Assessment of Hedychium coronarium Essential Oil on Fibrinogenolytic and Coagulant Activity Induced by Bothrops and Lachesis Snake Venoms. Journal of Venomous Animals and Toxins including Tropical Diseases, 20, 39-46.
https://doi.org/10.1186/1678-9199-20-39 - 11. Khanuja, S.P.S., Shasany, A.K., Srivastava, A. and Sushil, K. (2000) Assessment of Genetic Relationships in Mentha Species. Euphytica, 111, 121-125.
https://doi.org/10.1023/A:1003829512956 - 12. Benigni, R. (2005) Structure-Activity Relationship Studies of Chemical Mutagens and Carcinogens: Mechanistic Investigations and Prediction Approaches. Chemical Reviews, 105, 1767-1800.
https://doi.org/10.1021/cr030049y - 13. Brazilian Pharmacopoeia (2000) São Paulo.
- 14. Singh, N.P., Mccoy, M.T., Tice, R.R. and Schneider, E.L. (1988) A Simple Technique for Quantitation of Low Levels of DNA Damage in Individual Cells. Experimental Cell Research, 175, 184-191.
- 15. Collins, A.R., Dobson, V.L., Dusinská, M., Kennedy, G. and Stetina, R. (1997) The Comet Assay: What Can It Really Tell Us? Mutation Research, 375, 183-193.
- 16. Marcussi, S., Santos, P.R., Menaldo, D.L., Silveira, L.B., Santos-Filho, N.A., Mazzi, M.V., Da Silva, S.L., Stábeli, R.G., Antunes, L.M. and Soares, A.M. (2011) Evaluation of the Genotoxicity of Crotalus durissus terrificus Snake Venom and Its Isolated Toxins on Human Lymphocytes. Mutation Research, 724, 59-63.
- 17. Collins, A.R. (2004) The Comet Assay for DNA Damage and Repair. Principles, Applications and Limitations. Molecular Biotechnology, 26, 249-261.
https://doi.org/10.1385/MB:26:3:249 - 18. Preté, P.S., Domingues, C.C., Meirelles, N.C., Malheiros, S.V., Goñi, F.M. and De Paula, E. (2011) Multiple Stages of Detergent-Erythrocyte Membrane Interaction—A Spin Label Study. Biochimica et Biophysica Acta, 1808, 164-170.
- 19. Rangel, M., Malpezzi, E.L., Susini, S.M. and De Freitas, J.C. (1997) Hemolytic Activity in Extracts of the Diatom Nitzschia. Toxicon, 35, 305-309.
- 20. Gutiérrez, J.M., Avila, C., Rojas, E. and Cerdas, L. (1988) An alternative in Vitro Method for Testing the Potency of the Polyvalent Antivenom Produced in Costa Rica. Toxicon, 26, 411-413.
- 21. Price, M.F., Wilkinson, I.D. and Gentry, L.O. (1982) Plate Method for Detection on Phospholipase Activity in Candida albicans. Sabouraudia, 20, 7-14.
https://doi.org/10.1080/00362178285380031 - 22. Ferreira, D.F. (2011) Sisvar: A Computer Statistical Analysis System. Ciência e Agrotecnologia, 35, 1039-1042.
https://doi.org/10.1590/s1413-70542011000600001 - 23. Péres, V.F., Moura, D.J., Sperotto, A.R.M., Damasceno, F.C., Caramão, E.B., Zini, C.A. and Saffi, J. (2009) Chemical Composition and Cytotoxic, Mutagenic and Genotoxic Activities of the Essential oil from Piper gaudichaudianum Kunth Leaves. Food and Chemical Toxicology, 57, 2389-2395.
- 24. Rassouli, F.B., Matin, M.M., Iranshahi, M. and Bahrami, A.R. (2011) Investigating the Cytotoxic and Apoptosis Inducing Effects of Monoterpenoid Stylosin in Vitro. Fitoterapia, 82, 742-749.
- 25. Silva, M.R. and Guterres, Z.R. (2010) Investigation of Genotoxic and Antigenotoxic Activity of Ethanolic Extracts and Arbinic and Betulinic Acids Isolated from Combretum laxum. Anais do Encontro de Iniciação Científica, 8, 1-5.
- 26. Marya, C.M., Satija, G., Avinash, J., Nagpal, R., Kapoor, R. and Ahmad, A. (2012) In Vitro Inhibitory Effect of Clove Essential Oil and Its Two Active Principles on Tooth Decalcification by Apple Juice. International Journal of Dentistry, 2012, Article ID: 759618.
https://doi.org/10.1155/2012/759618 - 27. Quintans, J.S.S., Soares, B.M., Ferraz, R.P.C., Oliveira, A.C.A., Silva, T.B., Menezes, L.R.A., Sampaio, M.F., Prata, A.P., Moraes, M.O. and Pessoa, C. (2013) Chemical Constituents and Anticancer Effects of the Essential Oil from Leaves of Xylopia laevigata. Planta Medica, 79, 123-130.
https://doi.org/10.1055/s-0032-1328091 - 28. Misharina, T.A., FatkulliN, L.D., Alinkina, E.S., Kozachenko, A.I., Nagler, L.G., Medvedeva, I.B., Goloshchapov, A.N. and Burlakova, E.B. (2014) Effects of Low Doses of Essential Oils on the Antioxidant Status of the Erythrocytes, Liver and the Brain of Mice. Applied Biochemistry and Microbiology, 50, 88-93.
https://doi.org/10.1134/S0003683814010098 - 29. Terra, A.L.C. and Lema, T. (2007) Comparison of Toxicity between Venom of Snakes of the Genus Bothrops Present in Southern and Southeastern Brazil. Revista Brasileira de Toxicologia, 20, 55-63.
- 30. Fernandes, F.F., Tomaz, M.A., El-Kik, C.Z., Monteiro-Machado, M., Strauch, M.A., Cons, B.L., Tavares-Henriques, M.S., Cintra, A.C., Facundo, V.A. and Melo, P.A. (2014) Counteraction of Bothrops Snake Venoms by Combretum leprosum Root Extract and Arjunolic Acid. Journal of Ethnopharmacology, 155, 552-562.
- 31. Strauch, M.A., Tomaz, M.A., Monteiro-Machado, M., Ricardo, H.D., Cons, B.L., Fernandes, F.F., El-Kik, C.Z., Azevedo, M.S. and Melo, P.A. (2013) Antiophidic Activity of the Extract of the Amazon Plant Humirianthera ampla and Constituents. Journal of Ethnopharmacology, 145, 50-58.
- 32. Parente, L. (2001) Pros and Cons of Selective Inhibition of Cyclooxygenase-2 versus Dual Lipoxygenase/Cyclooxygenase Inhibition: Is Two Better than One? The Journal of Rheumatology, 28, 2375-2382.