H. L. LARSEN ET AL. 5
Poly-I:C-mediated IL-12p70 secretion and induction
of
n of PKR in αDC1s
m
pression levels in αDC1s, we ob-
se
sion, we identified a Poly-I:C dependent
lo
5. Acknowledgements
the Faculty of Health Sci-
REFERENCES
[1] F. Sallusto anicient Presentation
type I IFNs in αDC1s [5] makes this TLR3 ligand in-
dispensable in the α-Type-1 polarizing cocktail, and thus
removal of Poly-I:C from the cytokine cocktail in order
to restore DC viability is undesirable. However, apop-
tosis of DCs generated for therapeutic cancer vaccination
is undesirable due to reduced DC yields from valuable
and limited DC precursor cells and also decreases lon-
gevity upon in vivo injection. Furthermore, the presence
of apoptotic cells can impair DC phenotypic maturation
by suppressing the expression of co-stimulatory mole-
cules [12], hereby impairing DC functionality and ulti-
mately vaccine efficacy. However, this does not seem to
occur at the percentage of dead cells occurring in our
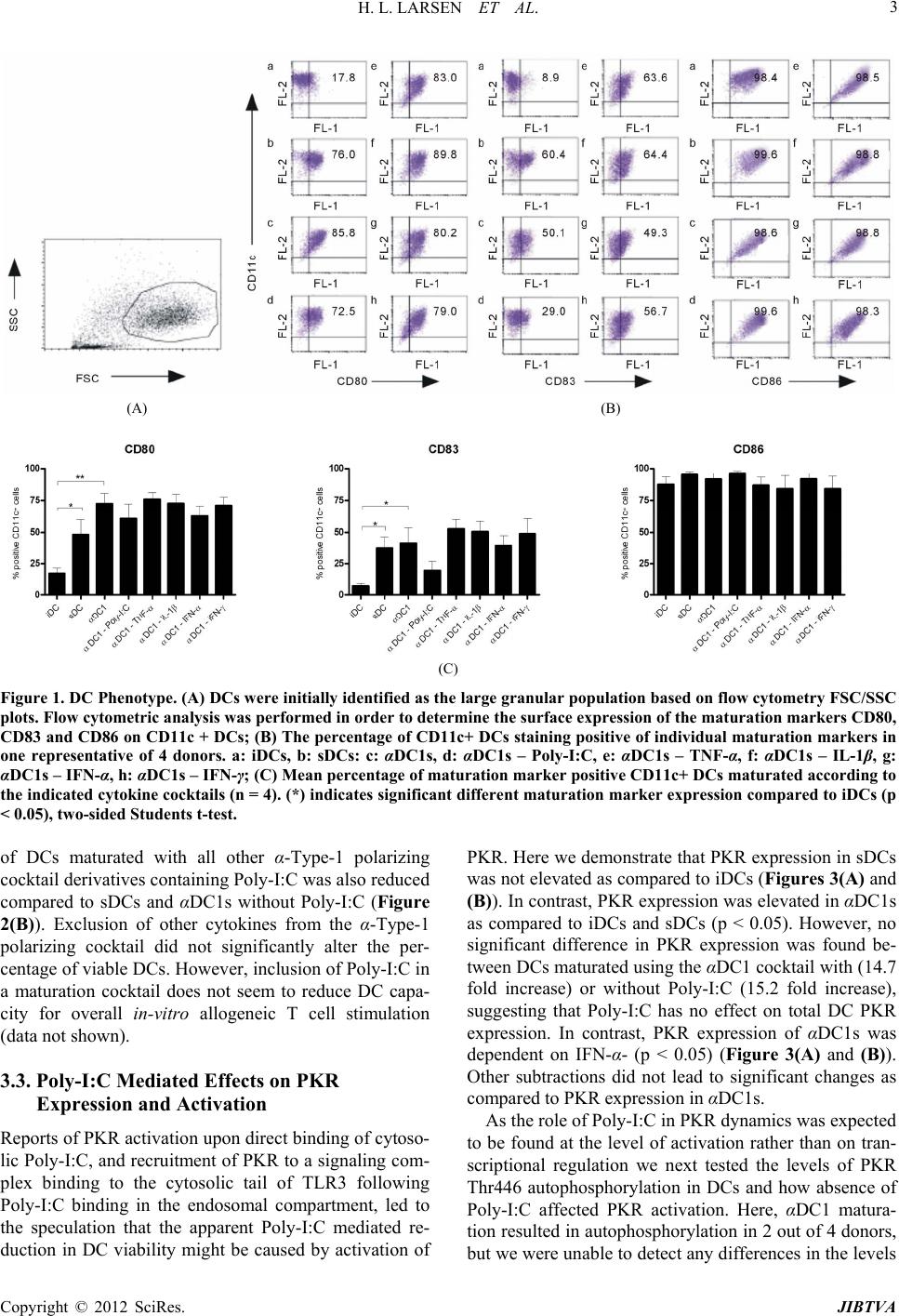
system (Figures 1(B) and (C)).
We hypothesized that activatio
ight cause the apparent Poly-I:C mediated cell death, as
active PKR induces translational inhibition and apoptosis
as a defense mechanism against viral infection [7,8].
Characterization of PKR expression in DCs revealed
significantly higher amounts of PKR in αDC1s and
αDC1s maturated without Poly-I:C as compared to sDCs
(Figure 3). The finding that presence of Poly-I:C has no
effect on PKR expression is in compliance with the
known role of Poly-I:C as a PKR activator and not as an
inducer of PKR transcription [6]. The high PKR expres-
sion observed in αDC1s seems to be induced by IFNs, as
αDC1s maturated without IFN-α showed a significantly
reduced PKR expression as compared to αDC1s. Matura-
tion of αDC1s without IFN-γ also seemed to reduce PKR
expression as compared to αDC1s however this reduced
expression was non-significant. These results confirm the
established role of IFNs as inducers of PKR expression
(reviewed in [13]).
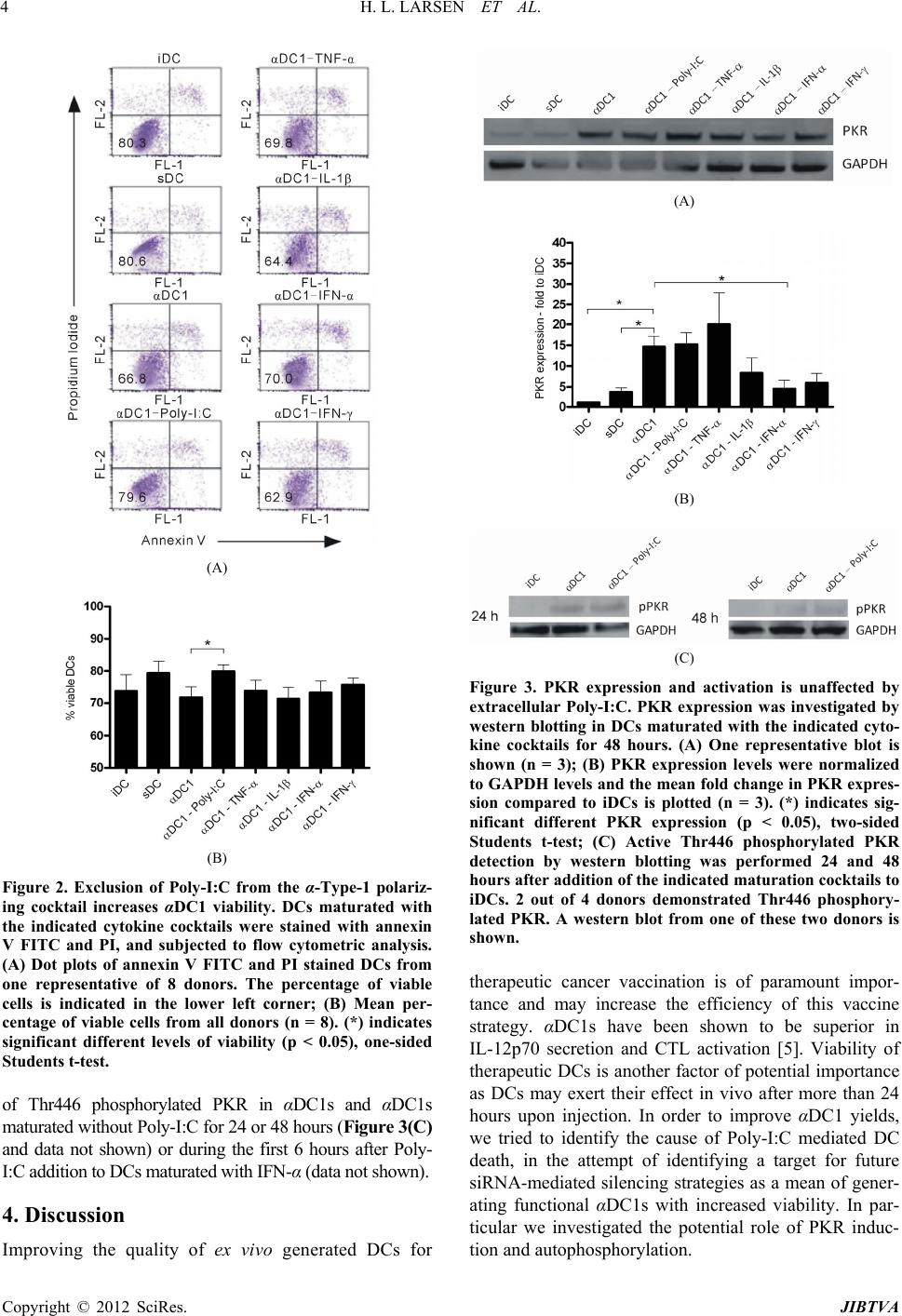
Despite high PKR ex
rved no significant difference in the amounts of active
Thr446 phosphorylated PKR in αDC1s and αDC1s
maturated without Poly-I:C (Figure 3(C)). The observed
lack of Poly-I:C mediated PKR activation at neither 48
hours after addition nor during the first 6 hours (data not
shown) suggests that PKR is not activated by extracellu-
lar Poly-I:C. These results argue against a role of acti-
vated PKR as the inducer of cell death in αDC1s despite
our previous findings of Poly-I:C as the cause of the
lowered viability of αDC1s. The apparent discrepancy
regarding the ability of Poly-I:C to induce Thr446 PKR
autophosphorylation in our hands and others might be
due to Poly-I:C residing in different compartments. PKR
activation by Poly-I:C reported by McAllister et al. was
observed after transfection, hereby enabling direct cyto-
solic binding of Poly-I:C to PKR [6], whereas Poly-I:C
was added extracellularly in our protocol for DC matura-
tion. We were unable to find any reports of translocation
of Poly-I:C from the endosomal compartment to the cy-
toplasm of DCs, thus explaining the lack of PKR activa-
tion. Furthermore, PKR activation following recruitment
to a TLR3 signaling complex in the endosome, activated
upon Poly-I:C addition was not directly detected by Jiang
et al., as the effect of transfection with dominant negative
PKR was only investigated on the level of NF-κB activ-
tion [10].
In conclu
wered viability of αDC1s where the mechanisms of
apoptosis was independent of PKR expression and acti-
vation. Future studies may reveal the targets for this
Poly-I:C mediated apoptosis which could be exploited
for siRNA silencing, thus maintaining the efficient αDC1
mediated maturation without hampering DC viability as
compared to sDCs.
This work was supported by
ences, University of Copenhagen and a grant from The
Augustinus Foundation.
d A. Lanzavecchia, “Eff
of Soluble Antigen by Cultured Human Dendritic Cells Is
Maintained by Granulocyte/Macrophage Colony-Stimula-
ting Factor plus Interleukin 4 and Downregulated by
Tumor Necrosis Factor Alpha,” The Journal of Experi-
mental Medicine, Vol. 179, No. 4, 1994, pp. 1109-1118.
doi:10.1084/jem.179.4.1109
[2] I. J. de Vries, W. J. Lesterhuis, N. M. Scharenborg, L. P.
Steinbrink, L.
Engelen, D. J. Ruiter, M. J. Gerritsen, et al., “Maturation
of Dendritic Cells Is a Prerequisite for Inducing Immune
Responses in Advanced Melanoma Patients,” Clinical
Cance Research, Vol. 9, 2003, p. 5091.
[3] H. Jonuleit, U. Kuhn, G. Muller, K.
Paragnik, E. Schmitt, et al., “Pro-Inflammatory Cytokines
and Prostaglandins Induce Maturation of Potent Immuno-
stimulatory Dendritic Cells under Fetal Calf Serum-Free
Conditions,” European Journal of Immunology, Vol. 27,
No. 12, 1997, pp. 3135-3142.
doi:10.1002/eji.1830271209
[4] R. Trepiakas, A. E. Pedersen, O. Met, M. H. Hansen, A.
Berntsen and I. M. Svane, “Comparison of Alpha-Type-1
Polarizing and Standard Dendritic Cell Cytokine Cocktail
for Maturation of Therapeutic Monocyte-Derived Den-
dritic Cell Preparations from Cancer Patients,” Vaccine,
Vol. 26, No. 23, 2008, pp. 2824-2832.
doi:10.1016/j.vaccine.2008.03.054
[5] R. B. Mailliard, A. Wankowicz-Kalinska, Q. Cai, A.
Wesa, C. M. Hilkens, M. L. Kapsenberg, et al., “Alpha-
Type-1 Polarized Dendritic Cells: A Novel Immunization
Tool with Optimized CTL-Inducing Activity,” Cancer
Research, Vol. 64, No. 17, 2004, pp. 5934-5937.
doi:10.1158/0008-5472.CAN-04-1261
[6] C. S. McAllister and C. E. Samuel, “The RNA-Activated
Protein Kinase Enhances the Induction of Interferon-Beta
Copyright © 2012 SciRes. JIBTVA