 Open Journal of Forestry 2012. Vol.2, No.2, 77-87 Published Online April 2012 in SciRes (http://www.SciRP.org/journal/ojf) http://dx.doi.org/10.4236/ojf.2012.22011 Copyright © 2012 SciRes. 77 Predicting Stem Windthrow Probability in a Northern Hardwood Forest Using a Wind Intensity Bio-Indicator Approach Philippe Nolet1,2, Frédérik Doyon1,2, Daniel Bouffard 3 1Insitut des Sciences de la Forêt tempérée, Ripon, Canada 2Université du Québec en Outaouais, Gatineau, Canada 3Insitut Québécois d’Aménagement de la Forêt Feuillue, Ripon, Canada Email: philippe.nolet@uqo. ca Received March 2nd, 2012; revised April 1st, 2012; accepted April 8th, 2012 Unlike fire or insect outbreaks, for which a suppression program can be implemented, it is impossible to prevent a windstorm event or stop it while it is occurring. Reducing stand susceptibility to windstorms requires a good understanding of the factors affecting this susceptibility. Distinct species- and size-related differences in stem windthrow susceptibility are difficult to obtain because it is impossible to distinguish their relative effects from those of wind intensity. Using a damage assessment database (60 20-metre ra- dius plots) acquired after an exceptional wind storm in Western Quebec in 2007, we developed an ap- proach in which proportions of windthrown sugar maple poles were used as bio-indicators of wind inten- sities affecting the plots. We distinguished between single and interactive effects of wind intensity, spe- cies, stem size, and local basal area on stem windthrow susceptibility. The best logistic regression model predicting stem windthrow included the wind intensity bio-indicator, species, basal area, and the species by diameter at breast height (DBH, 1.3 m) interaction. Stem windthrow probability generally increased with DBH and decreased with basal area. Species wind-firmness was ordered as: yellow birch > sugar maple = eastern hemlock = American beech > ironwood > basswood = other hardwoods = other soft- woods. Our method remained an indirect method of measuring wind intensity and its real test would re- quire a comparison with anemometer measurements during a windstorm. Despite its indirect nature, the method is both simple and ecologically sound. Hence, it opens the door to conducting similar windthrow studies in other ecosystems. Keywords: Windthrow Prediction; Species Wind-Firmness; Tolerant Hardwood Forests; Silviculture Guidelines Introduction Windthrow represents one of the most important catastrophic natural disturbances in shade-tolerant hardwood stands of North America (Bormann & Likens, 1979; Canham & Loucks, 1984; Foster, 1988; Seymour et al., 2002). Mortality from windthrow in these forests can be comparable to that which results from single tree mortality (Woods, 2004; Nolet et al., 2007). As with other natural disturbances, windthrow affects ecosystem ele- ments, such as stand structural complexity (Han son & Lorimer, 2007; Fukui et al., 2011; Kneeshaw et al., 2011) and soil char- acteristics (Clinton & Baker, 2000; Simon et al., 2011), which have key roles in ecosystem processes (Putz et al., 1983). It also affects forestry operations by reducing timber volume availabil- ity and by drastically changing harvest schedules, which both bring economic losses. Unlike fire (Stephens & Ruth, 2005) or insect outbreaks (e.g. Stedinger, 1984), for which a suppression program can be implemented to reduce such economic losses, a windstorm event cannot be prevented nor can it be stopped while it is occurring. The only action that forest managers can take is to reduce stand susceptibility to windstorms, which first requires a good understanding of the factors that affect this susceptibility. Many studies have addressed this issue, but these have been mainly conducted for coniferous stands (Huggard et al., 1999; Ruel, 2000; Ni Dhubhain et al., 2001). Several factors influence a tree’s probability of being wind- thrown (Everham & Brokaw, 1996) and such factors operate at different scales (Ruel, 1995; Boose et al., 2001; Scott & Mitchell, 2005; Valinger & Fridman, 2011). These range from the landscape scale (exposure to prevailing storm winds) to the stand scale (moisture regime, soil thickness, proximity to openings, stand basal area) and down to the individual stem (species, stem size, stem health). At the level of the individual stem, species and size play essential roles in windthrow suscep- tibility. It has been generally observed that 1) larger stems are more vulnerable than smaller ones (Peterson, 2007), 2) soft- wood species are more vulnerable than hardwoods (Foster, 1988; Scott & Mitchell, 2005), and 3) early-successional spe- cies are more vulnerable than late-successional species (Foster, 1988; Everham & Brokaw, 1996; Rich et al., 2007). In forests, where many species and diameter classes may be found, such generalities are not sufficient to develop prevention practices that are aimed at decreasing stand susceptibility to windstorms. Disentangl i ng t he E ffe cts of Wind Intensi t y and Susceptibility Factors The severity of a windthrow event in a given stand is a  P. NOLET ET AL. product of the wind intensity that affects the stand and the various factors that influence stem windthrow susceptibility. Distinct differences in stem susceptibility to windthrow, which vary according to species and size, are difficult to obtain be- cause, for most of the time, it is impossible to distinguish their relative effects from those of wind intensity and site factors. For instance, if an eastern hemlock (Tsuga canadensis L. Carr.) dominated stand is more greatly affected by windthrow during a storm event than the surrounding hardwood stands, it is very difficult to determine whether this is a result of 1) more intense winds at that specific location, 2) this particular species’ sus- ceptibility, 3) the size of the stems, or 4) simply the site condi- tions (e.g., soil moisture) at that specific location. To disentan- gle the effects of wind intensity and susceptibility factors re- quires a measure of wind intensity that has affected each spe- cific stand. Given the frequency and unpredictability of such events, it is almost impossible to obtain direct measures (e.g., anemometer readings) of wind intensity during a windstorm. Canham et al. (2001) have developed an innovative approach that computes the relative importance of wind intensity and stem characteristics in estimating the probability that a given stem will be windthrown. The novelty of this approach relies on three main aspects: 1) the measure of wind intensity is indirect; 2) the differences in wind intensity that are experienced by plots (or stands) are relative to one another; and 3) this indirect and relative wind intensity measure is obtained simultaneously through the computation of species-specific susceptibility to windthrow using a global pseudo-optimization (GPO) proce- dure. Having an indirect and relative measure of wind intensity (the first two aspects of the approach) is a major innovation since windthrow causality due to wind intensity can be distin- guished from the effect of species-specific characteristics, to- gether with testing the interaction between these variables. However, we believe that the third aspect of the approach (the GPO procedure) is weakest since it requires the use of a large number of parameters in the statistical model (64 in Canham et al., 2001). To avoid spurious effects that could be incurred by including many parameters in a model, Anderson et al. (2001) argue that their numbers should be limited and that models with 30-plus parameters often find little support. We sought a method that could use the best aspects of the approach developed by Canham et al. (2001) (1 and 2) and improve its weakest aspect (3). We investigated a damage as- sessment database (60 20-metre radius plots) that was acquired after an exceptional wind storm in Western Quebec (Environ- ment Canada, 2007). We observed sugar maple poles (i.e., stems 9.1 - 19.0 cm diameter at breast height, DBH, 1.3 m above the ground surface) in all pre-storm stands. This allows us to implement an idea put forth by Wood (1995), which con- sists of using the proportion of windthrown stems of a definite species and size as an indicator of wind intensity. By calculat- ing the proportion of windthrown sugar maple (Acer saccharum Marsh.) poles within each plot, we obtain a value that can be interpreted as an indirect and relative measure of wind intensity. In other words, we believe that sugar maple poles in our dataset may be used as a bio-indicator of wind intensity that has af- fected the plots. Therefore, the objectives of this paper are: to distinguish between the individual and interactive effects of wind intensity, species, stem size, and local basal area on stem susceptib ility to windthrow using a wind bi o-indicator approach, and to verify how such results may be taken into account in selection cuts to prevent eventual loss from windthrow in northern hardwood forests. Materials and Methods Study Area This study was conducted in the Papineau-Labelle Wildlife Reserve (46˚13'48''W, 75˚09'55''N) of Quebec, between Lakes Montjoie and Du Sourd, and about 100 km northeast of Can- ada’s capital, Ottawa. The area is located in the eastern portion of Lac du Poisson Blanc landscape unit (Robitaille & Saucier, 1998) of the western sugar maple-yellow birch (Betula al- leghaniensis Britton) bioclimatic region (Saucier et al., 2011). The landscape contains numerous hills with elevations < 450 m a.s.l. and averaging 300 m in height. Mean annual temperature is 3.7˚C, mean annual precipitation is roughly 1100 mm (in- cluding 250 mm as snow), and the number of degree days above 0˚C is 2716 (Environment Canada, 2007). Surficial ge- ology for the study area is characterized by thin to moderately thin glacial till composed of metamorphic rocks, such as gneiss, topped by sandy Dystric Brunisols (GPPC, 2010). The forest canopy is dominated by sugar maple in association with yellow birch, American beech (Fagus grandifolia Ehrh.), American basswood (Tilia Americana L.), ironwood (Ostrya virginiana Mill. K. Koch), eastern hemlock, and balsam fir (Abies bal- samea L. Mill.). The forest inventory map, which was produced for the area by the provincial government, indicates that partial harvesting was conducted in the stands in 2001. Observations made on site at the time of sampling confirmed this informa- tion. Storm Event The storm that caused the windthrow event discussed here had occurred on July 17th 2006. On the day of the storm, the nearest airport (Maniwaki Airport; 46˚16'W, 75˚59'N) regis- tered peak wind speeds reaching 61 km·hour–1 (Environment Canada, 2011), while the weather station at Rouyn-Noranda, Quebec (48˚03'W, 77˚47'N), registered peak winds of 117 km·hour–1 (Environment Canada, 2011). Given the damage observed in some stands, peak winds in the study area were likely closer to the winds observed at Rouyn than those re- ported at Maniwaki. Sampling Protocol Sixty 20-metre radius plots were established within the study area. A stratified random sampling scheme was applied to ob- tain a range of windthrow severities; 20 plots were established in each class of windthrow severity (high, medium, and low based on visual estimation). These classes were only used to stratify the sampling scheme and were not used in the ensuing analysis. To minimize spatial autocorrelation (stand composi- tion, wind severity) or other spatial dependencies among plots, plots were separated by a minimum distance of 100 m. In addi- tion, to avoid pseudo-replication and to maintain interspersion of disturbance levels, we ensured that there was a change in the disturbance severity classes in between the plots. We sampled only mesic sites with gentle slopes (<15%) to minimize the effects of site differences on windthrow probabilities. Selection and characterization of windthrown stems followed (Canham et al., 2001; Woods, 2004). All windthrown trees Copyright © 2012 SciRes. 78  P. NOLET ET AL. within plots that had a diameter at breast height (DBH, 1.3 m above the soil surface before windthrow) greater than 9.1 cm were included in the study. A stem was considered to be wind- thrown if it rested at a horizontal angle < 45˚, or if the stem was broken below the base of the crown. For all windthrown stems, species and DBH were also recorded. Data Analysis As previously mentioned, we calculated a bio-indicator of wind intensity (BWI) for each sampling plot by dividing the number of windthrown sugar maple poles (DBH class 9.1 - 19.0 cm) by the total number of sugar maple poles that were present. We used only plots having at least four sugar maple poles prior windthrow, which required that eight of the initial 60 plots be excluded. The probability of a stem being wind- thrown was predicted as a function of four potential variables, i.e., BWI, species, DBH, and stand basal areal (BA), together with interactions among these variables. Using the model selec- tion approach (Anderson et al., 2000; Johnson & Omland, 2004), we compared 22 logistic regression models (using the R package, R Development Core Team, 2011). Nine models had only single effects, while thirteen models included two-way interactions among the explanatory variables. The model selec- tion approach is especially well-suited to complex systems for which a common model selection procedure could lead inaccu- rately to the identification of one “best” model without ac- counting for the large uncertainty in the model selection itself. Model performance was verified mainly through Akaike’s In- formation Criterion (AIC), which was corrected for small sam- ple sizes (AICc). From this value, we calculated the Akaike weight to rank the models and, for each model, to evaluate its probability of being the best option. We did not test for any three-way interactions, or for any interaction term that included BA. We could not find any ecological reasons to test interac- tions including BA, which is a local stand-level variable. BA was used in some models to verify whether or not its addition would improve model performance compared to the corre- sponding models that did not incorporate it. Once the highest performing and most ecologically sound model was chosen, we tested its goodness-of-fit in three ways. First, we verified if the model fit equally well across the range of predicted probabilities. To do so, we computed and plotted the observed mean proportion of windthrown stems by classes of stem-predicted windthrow probabilities. Second, we com- pared observed windthrow proportions in the plots with the mean predicted windthrow probabilities. Finally, we compared observed windthrow proportions to mean predicted windthrow probabilities by species-DBH (10-cm classes) combinations. To verify how the model can be useful in decreasing stand susceptibility to windthrow, we compared the possible loss due to windthrow after applying two hypothetical partial cut treat- ments. The first treatment was designed to mimic a typical single-tree selection cut (SC) in which 30% of the basal area is harvested equally among species and DBH classes. The second treatment, a wind-firm optimized partial cut (WOP), was de- signed to harvest the less wind-firm stems of the stand, as pre- dicted by our model, until 30% basal area removal was attained. These treatments were applied to the same and single hypo- thetical stand, represented by the same structure and composi- tion as that described in Table 1, except that stem number was multiplied to achieve a basal area of 26 m2·ha–1. This basal area is representative of the pre-harvest stands in maple-dominated forests in Québec. After applying these two treatments on this initial stand, two theoretical residual stands remained. Then, employing our model, we computed the expected loss of basal area in these stands that would result from an eventual wind storm (relative intensity = .5). Results Sugar maple (SM) was the most abundant species in the plots, followed by American beech (AB) and yellow birch (YB) (Ta- ble 1). Three other species, viz., ironwood (IW), eastern hem- lock (EH) and, American basswood (BW), were represented by at least 20 individual trees in the dataset. All species were pre- sent in all DBH classes, except for IW, which occurred only in the two smallest DBH classes. Species with a frequency < 20 were grouped with “other hardwoods” (OH) or “other soft- woods” (OS). The OH group included red maple (Acer rubrum L.), white or American ash (Fraxinus americana L.), white or paper birch (Betula papyrifera Marsh.), trembling aspen (Populus tremuloides Michx.), and black cherry (Prunus serot- ina Ehrh.). The OS group included balsam fir, eastern white- cedar (Thuja occidentalis L.), and white spruce (Picea glauca Moench Voss). Such species compositions and DBH distribu- tions were representative of stands in the study area. Comparing models that used only one explanatory variable (models 1 to 4; Table 2) clearly showed that BWI was the best variable for predicting stem windthrow probabilities, followed in decreasing order by species, DBH, and BA. By gradually increasing model complexity, we observed that adding species and DBH (model 8) markedly increased model performance (decreasing AICc) compared to the use of BWI alone (model 1). At this step of the analyses, adding BA (model 9) decreased model performance. The use of BWI*DBH interaction de- creased model performance slightly (models 10 to 13), while BWI*species slightly increased it (models 14 to 16). The DBH*species (models 17 to 19) interaction clearly contributed the most to model performance. Including both BWI*species and DBH*species interactions led to the best model perform- ance (model 21)—slightly better than model 19—but also re- quired a substantial increase in the number of parameters used in the model. After the incorporation of interactions in the models (models 10 to 22), models using BA consistently per- formed better than the corresponding models not using BA (i.e., model 13 vs 12; model 16 vs 15; model 19 vs 18; model 21 vs 20). Overall, and according to the model weights (Wi in Table 2), two models (models 19 and 21) were superior to the others; since model 21 did not clearly outperform model 19, we cannot affirm that the former was definitely the best model. The plot- ting of windthrow probability against BWI (with DBH and BA held constant), according to models 19 and 21 (Figures 1 and 2), revealed two important facts. First, the confidence intervals were much wider for model 21 than for model 19. Second, the effect of BWI on windthrow probability was positive for most species and for both models, but it was negative for the OH group, according to model 21. We argue that it is impossible that a stem windthrow probability decreases with wind intensity; therefore, we can no longer consider this model as valid. We believe that the increased numbers of parameters in model 21, compared to model 19, led to an artificial and misleading in- crease in model performance (beter AICc). Due to these issue s, t Copyright © 2012 SciRes. 79  P. NOLET ET AL. Copyright © 2012 SciRes. 80 Table 1. Stem density (individuals ·ha–1) by species and diameter classes for the whole dataset (52 plots). DBH class/species AB BW EH IW OH OS SM YB Total 9.1 - 19.0 cm 231 10 1 64 5 19 606 135 1071 19.1 - 1 - 29.0 cm 123 9 12 9 14 3 623 76 869 29.1 - 39.0 cm 57 3 15 8 2 321 20 426 >39.1 cm 20 2 23 2 6 147 5 205 Total 431 24 51 73 29 30 1697 236 2571 AB = American beech; BW = American basswood; EH = eastern hemlock; IW = ironwood; OH = other hardwood species; OS = other softwood species; SM = sugar maple; YB = yellow birch. Table 2. Comparison of vari ous stem windthrow logistic models. No Interactions Single effects K Log-likelihood AICc Wi 1 none BWI 2 –852.2 1708.4 0% 2 none DBH 2 –908.7 1821.3 0% 3 none SP 8 –886.4 1788.9 0% 4 none BA 2 –915.8 1835.6 0% 5 none BWI + DBH 3 –846.9 1699.8 0% 6 none BWI + SP 9 –836.0 1690.1 0% 7 none DBH + SP 9 –881.7 1781.4 0% 8 none BWI + DBH + SP 10 –826.4 1672.8 0% 9 none BWI + DBH + SP + BA 11 –826.1 1674.2 0% 10 BWI*DBH SP 9 –831.2 1680.5 0% 11 BWI*DBH DBH + SP 10 –831.2 1682.4 0% 12 BWI*DBH BWI + DBH + SP 11 –828.7 1679.5 0 % 13 BWI*DBH BWI + DBH + SP + BA 12 –824.7 1673.5 0% 14 BWI*SP DBH 10 –834.0 1688.0 0% 15 BWI*SP DBH + SP 17 –818.3 1670.9 0% 16 BWI*SP DBH + SP + BA 18 –813.1 1662.4 0% 17 DBH*SP BWI 9 –829.9 1677.9 0% 18 DBH*SP BWI + SP 17 –812.2 1658.6 3% 19 DBH*SP BWI + SP + BA 18 –809.3 1654.8 20% 20 DBH*SP + BWI*SP SP 24 –805.0 1658.4 3% 21 DBH*SP + BWI*SP SP + BA 25 –800.9 1652.3 70% 22 DBH*SP + BWI*SP BA 18 –811.1 1658.5 3% AICc = corrected Akaike info rmation criteri on; K = number of model pa rameters; Wi = weight of the model compared to othe r models; B WI = bio -indicator of wind i nt en- sity; DBH = diameter at breast height; SP = species; BA = basal area. we concluded model 19 to be our best model. Model 19 adequately fitted the observed data regardless of how the data were summarized. First, the model provided valid results for most of its prediction range (Figure 3). For the classes of predicted windthrow probability ranging from .0 to .6, the model closely predicted the observed probabilities. For probability classes ranging from .6 to .9, the model overesti- mated stem windthrow probability by about 15%, while fitting the highest class (.9 to 1.0) very well. Second, the predicted proportion of windthrown stems by plot was strongly associ- ated with the observed proportion of windthrown stems (R2 = .72) and the relation is close to 1 (1.17, Figure 4). Th ird, an d most importantly, the model closely predicted the proportion of windthrown stems that were observed in most species-DBH combinations (Figure 5). It fitted the data very well for the most frequently encountered speces, SM, AB, and YB, although i 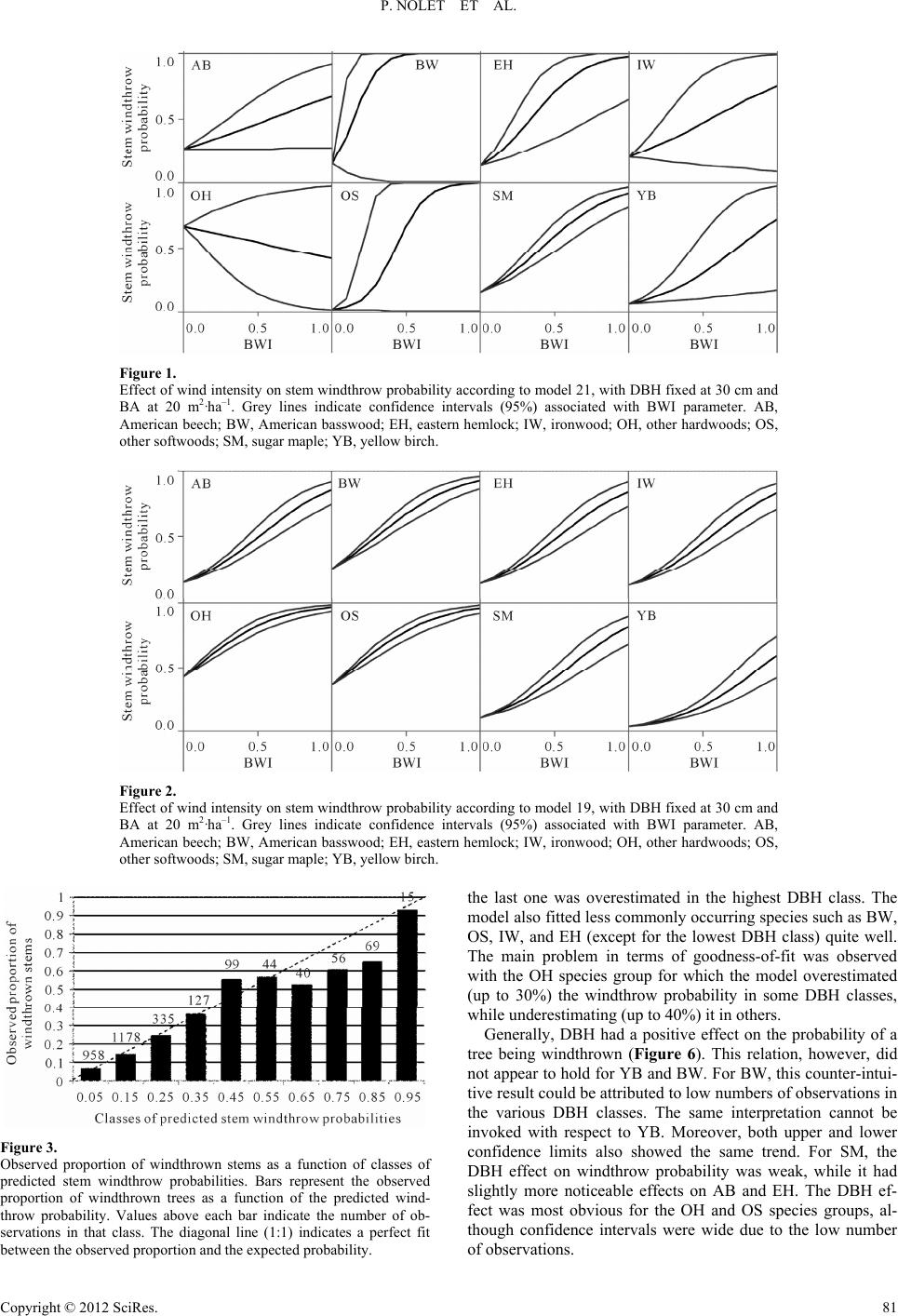 P. NOLET ET AL. Figure 1. Effect of wind intensity on stem windthrow probability according to model 21, with DBH fixed at 30 cm and BA at 20 m2·ha–1. Grey lines indicate confidence intervals (95%) associated with BWI parameter. AB, American beech; BW, American basswood; EH, eastern hemlock; IW, ironwood; OH, other hardwoods; OS, other softwoods; SM, sugar maple; YB, yellow birch. Figure 2. Effect of wind intensity on stem windthrow probability according to model 19, with DBH fixed at 30 cm and BA at 20 m2·ha–1. Grey lines indicate confidence intervals (95%) associated with BWI parameter. AB, American beech; BW, American basswood; EH, eastern hemlock; IW, ironwood; OH, other hardwoods; OS, other softwoods; SM, sugar maple; YB, yellow birch. Figure 3. Observed proportion of windthrown stems as a function of classes of predicted stem windthrow probabilities. Bars represent the observed proportion of windthrown trees as a function of the predicted wind- throw probability. Values above each bar indicate the number of ob- servations in that class. The diagonal line (1:1) indicates a perfect fit between the observed proportion and the expected proba b ility. the last one was overestimated in the highest DBH class. The model also fitted less commonly occurring species such as BW, OS, IW, and EH (except for the lowest DBH class) quite well. The main problem in terms of goodness-of-fit was observed with the OH species group for which the model overestimated (up to 30%) the windthrow probability in some DBH classes, while underestimating (up to 40%) it in others. Generally, DBH had a positive effect on the probability of a tree being windthrown (Figure 6). This relation, however, did not appear to hold for YB and BW. For BW, this counter-intui- tive result could be attributed to low numbers of observations in the various DBH classes. The same interpretation cannot be invoked with respect to YB. Moreover, both upper and lower confidence limits also showed the same trend. For SM, the DBH effect on windthrow probability was weak, while it had slightly more noticeable effects on AB and EH. The DBH ef- fect was most obvious for the OH and OS species groups, al- though confidence intervals were wide due to the low number of observations. Copyright © 2012 SciRes. 81 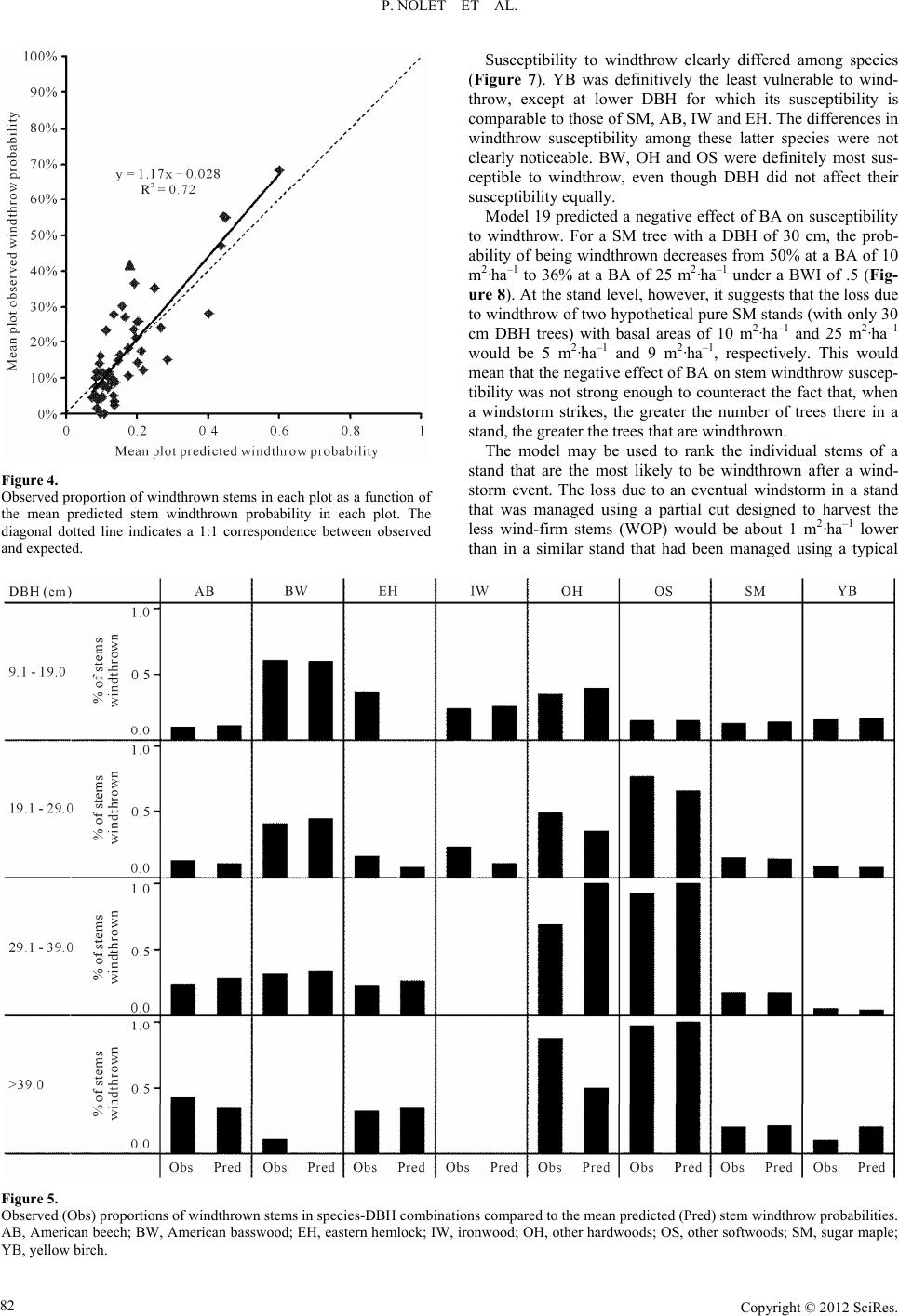 P. NOLET ET AL. Figure 4. Observed proportion of windthrown stems in each plot as a function of the mean predicted stem windthrown probability in each plot. The diagonal dotted line indicates a 1:1 correspondence between observed and expected. Susceptibility to windthrow clearly differed among species (Figure 7). YB was definitively the least vulnerable to wind- throw, except at lower DBH for which its susceptibility is comparable to those of SM, AB, IW and EH. The differences in windthrow susceptibility among these latter species were not clearly noticeable. BW, OH and OS were definitely most sus- ceptible to windthrow, even though DBH did not affect their susceptibility eq ually. Model 19 predicted a negative effect of BA on susceptibility to windthrow. For a SM tree with a DBH of 30 cm, the prob- ability of being windthrown decreases from 50% at a BA of 10 m2·ha–1 to 36% at a BA of 25 m2·ha–1 under a BWI of .5 (Fig- ure 8). At the stand level, however, it suggests that the loss due to windthrow of two hypothetical pure SM stands (with only 30 cm DBH trees) with basal areas of 10 m2·ha–1 and 25 m2·ha–1 would be 5 m2·ha–1 and 9 m2·ha–1, respectively. This would mean that the negative effect of BA on stem windthrow suscep- tibility was not strong enough to counteract the fact that, when a windstorm strikes, the greater the number of trees there in a stand, the greater the trees that are windthrown. The model may be used to rank the individual stems of a stand that are the most likely to be windthrown after a wind- storm event. The loss due to an eventual windstorm in a stand that was managed using a partial cut designed to harvest the less wind-firm stems (WOP) would be about 1 m2·ha–1 lower than in a similar stand that had been managed using a typical Figure 5. Observed (Obs) proportions of windthrown ste ms in species-DBH combinations compared to the mean predicted (Pred) stem windthrow probabilities. AB, American beech; BW, American basswood; EH, eastern hemlock; IW, ironwood; OH, other hardwoods; OS, other softwoods; SM, sugar maple; YB, yellow birch. Copyright © 2012 SciRes. 82  P. NOLET ET AL. Figure 6. Effect of DBH on stem windthrow probability according to model 19 with BWI fixed at .5 and basal area at 20 m2·ha–1. Grey lines indicate confidence intervals (95%) associated with DBH parameters. AB, American beech; BW, American basswood; EH, eastern hemlock; IW, ironwood; OH, other hardwoods; OS, other softwoods; SM, sugar maple; YB, yellow birch. Figure 7. Comparisons among species of the effect of DBH on stem windthrow probability according to model 19 with BWI fixed at .5 and BA at 20 m2·ha–1. The curves, which are the same than those shown in Figure 6, are presented without the confidence intervals to facilitate the com- parison among species. AB, American beech; BW, American basswood; EH, eastern hemlock; IW, ironwood; OH, other hardwoods; OS, other softwoods; SM, sugar maple; YB, yellow birch. tree-selection cut (Figure 9). This result was valid only for the example we used (see methodology). Results could vary de- pending on the initial stand characteristics and storm intensity. Discussion Wood (1995) first proposed the use of species-DBH combi- nations as surrogates for wind intensity. Rich et al. (2007) partly applied the idea a few years later building a surrogate that implied many species having a similar DBH distribution. To our knowledge, the present study was the first to implement Wood’s idea in its exact form. To be used as a surrogate for a specific disturbance agent, a bio-indicator should exhibit bal- anced sensitivity to the disturbance agent. A bio-indicator that Figure 8. Effect of BA on stem windthrow probability according to model 19, with BWI fixed at 0.5 and DBH at 30 cm. Grey lines indicate confi- dence intervals (95%) associated to BA parameter. is too sensitive will fail to distinguish differences at higher values within the spectrum of disturbance intensities, while a lack of sensitivity will lead to a similar problem within the lower end of that same spectrum. Since the proportion of SM poles that were windthrown varied from .0 to .7 in our plots, it is reasonable to believe that SM poles exhibited sufficient sen- sitivity to windthrow to be used as a bio-indicator. Even though BWI (bio-indicator of wind intensity) was the most important variable in our model, we cannot verify how it was related to the actual wind intensity experienced by the plots. In other words, BWI seems to be related to wind intensity, but we can- not demonstrate it. The mathematical approach developed by (Canham et al., 2001) has the same limitations. In addition, using the approach developed by the aforementioned authors would have required the estimation of 85 parameters (60 for the Copyright © 2012 SciRes. 83  P. NOLET ET AL. Figure 9. Comparison of predicted loss due to windthrow after a typical tree selection partial cut (TSP) and after a wind-firm-optimized partial cut (WOP). The initial stand composition and structure correspond to the average of all plots in the da taset, standardized at 26 m2·ha–1. plots, 24 for the species, and 1 for BA). Since we already ob- served that the use of 26 parameters led to over-fitting of model 21, it is likely that using 85 would have led to a simila r problem. Anderson et al. (2000) have urged ecologists not to model the data; rather, they should attempt to isolate informa- tion in the data from the noise. The wind bio-indicator ap- proach that was developed in this study is in concordance with this advice. According to our results, we could order species from most to least wind-firm as follows: yellow birch (YB) > sugar maple (SM) = eastern hemlock (EH) = American beech (AB) > iron- wood (IW) > American basswood (BW) = other hardwoods (OH) = other softwoods (OS). Likewise, we have summarized results from several other northern hardwood forests (Table 3) to facilitate comparisons with our results. Other studies have observed that yellow birch (YB) was the least, or among the least, vulnerable species (Canham et al., 2001; Woods, 2004; Hanson & Lorimer, 2007) (Table 3). Peterson (2007), who studied many sites, found that the ranking of YB susceptibility was inconsistent when compared with other species. Sugar maple (SM) was the second most resistant species according to our study. Incidentally, Peterson (2007) observed the same classifying order for SM in his three study sites reported in Table 3. Canham et al. (2001) reported SM and YB as the most resistant species to windstorm, as in concordance with our re- sults. American beech (AB) exhibited susceptibility very simi- lar to SM (Woods, 2004; Person, 2007; this study). Only Can- ham et al. (2001) observed a much higher susceptibility for AB than for SM and this was probably due to the beech bark dis- ease (Papaik et al., 2005). Eastern hemlock (EH) shows the least consistent susceptibility ranking among the studies. While its susceptibility was similar to that of SM in our study, hem- lock appeared to be less vulnerable (than SM) according to Woods (2004), and more vulnerable according to Canham et al. (2001) and Hanson & Lorimer (2007). In the studies reported in Table 3, Peterson (2007) found that EH had a susceptibility similar to that of SM in a first site, lower susceptibility in a second site, and higher susceptibility in a third site. It was noteworthy that EH was always more wind-firm than other softwood species. When present, OS was always the most vul- nerable species, according to the rankings in Table 3. In our study, the OH species group was as vulnerable as OS. Canham et al. (2001) and Peterson (2007) also observed that this species group was usually highly vulnerable to windstorm. However, Peterson (2007) also observed a very high wind-firmness for species with strong wood, such as hickories (Carya species) and white oak (Quercus alba L.). Overall, despite the variety of ecosystems that were studied, the variety of the windstorms that had affected them, and the variety of methods that were used to study them, some general conclusions may be drawn. First, among the most frequently occurring species in the northern hardwoods, YB seemed to be the most wind-firm, followed by SM, EH, and AB. Second, softwood species were the less wind-firm most of the time, except for EH. Clearly, a species’ successional status was not a good indicator of wind-firmness. The higher wind-firmness of YB and SM compared to that of EH and AB was such an ex- ample, since the latter are recognized as late-successional spe- cies when compared to the former (Doyon et al., 1998). Also, Table 3. Comparison of species wind- fir mness relative ranking according to various studies in no rt h e as t er n ha r d wo o d f o re s t s o f N o rt h America. Species wind-firmness relative ranking1 according to Species2 This study Canham et al. (2001) Hanson & Lorimer (2007)Peterson (2007) Tionesa’94 Peterson (2007) TexHill Peterson (2007) Gould Woods (2004) AB 2 3 - 2 - - 2 BW 4 - - - - - - EH 2 2 2 1 3 2 1 IW 3 - - - - - - OH 4 2 and 4 - 2 1 and 3 3 - OS 4 4 - - - 4 - SM 2 1 1 2 2 2 2 YB 1 1 1 3 - 1 1 1Species wind-firmness rank was not always explicitly specified by the authors , so we attribu ted a ran king using figures and tab les provid ed i n th e articles. Althoug h some species rankin gs could be es tablished easily, others required some subjectivity. A ranking of 1 meant that the species was considered the most wind-firm for the study or site; 2AB = American beech; BW= American basswood; EH = eastern hemlock; IW = ironwood; OH = other hardwoods; OS = other softwoods; SM = sugar maple; YB = ellow birch. y Copyright © 2012 SciRes. 84  P. NOLET ET AL. Peterson (2007) observed that, hickories and sugar maple, con- sidered early- and late-successional species, respectively, pre- sent similar traits (deep rooting and strong wood) that result in high windfirmness. As outlined by (Nolet et al., 2008), it is more appropriate to use species-specific traits to explain eco- logical phenomena (in this case, species wind-firmness) than broad and subjective categories such as early- or late-succes- sional status. As observed in most studies (see Everham & Brokaw, 1996), stem windthrow probability increased with DBH for most spe- cies included in our study. In a study using a methodology similar to that applied in this study, but driven in a completely different ecosystem, (Rich et al., 2007) observed a positive relationship between wind intensity and the nine species that they studied. In our study, the strength of the relationships was positive and relatively weak for SM, AB, and EH, while it was positive and strong for OH and OS. Canham et al. (2001) also observed that the DBH effect on windthrow probability was stronger for less wind-firm species. However, one of the most surprising results of the present study was certainly the negative effect of DBH on windthrow probability for YB. We believe that YB was a highly wind-firm species across the whole range of DBH that was studied. Further, we observed a higher wind- throw probability for small stems only because they were more likely to be caught in a domino effect due to a large stem falling. This domino effect, which is often observed in the field (al- though very difficult to identify and quantify precisely), af- fected not only YB but all of the species. For less wind-firm species, however, the domino effect would not lead to a nega- tive relationship between windthrow probability and DBH. Although species and DBH had effects on stem windthrow probabilities, their interaction with wind intensity was not per- ceivable in our dataset. Therefore, according to our model, the relative rank of species wind-firmness was not influenced by wind intensity. This result contradicted Canham et al. (2001), whose model relied on an interaction between wind intensity, DBH, and species. In our case, the absence of any interaction between wind intensity and species on one hand, and wind intensity and DBH on the other hand, decreased the complexity of the model and simplified its potential use for silvicultural prescriptions at the site level or for planning strategies at the landscape level (e.g., Papaik & Canham, 2006). Everham and Brokaw (1996) reported many studies that have observed more windthrow damage in thinned stands than in unmanaged stands. This phenomenon may be generally attrib- uted to the wind’s ability to penetrate more thoroughly into stands that have been partially harvested, thus increasing the force exerted on trees (Peltola et al., 1999). Everham and Bro- kaw (1996) also reported studies that have observed less effect or no effect at all of thinning on stand windthrow damage. It appeared that, when a positive effect of thinning on wind- firmness is observed, it is probably because the weakest trees had already been harvested during the thinning treatment. The methodology used in this study presents the advantage of not relying on a managed vs unmanaged comparison since it di- rectly assessed the effect of surrounding densities on stem windthrow probability. Our results indicated that tree wind- throw probability was slightly higher for stems surrounded by lower BA (i.e., about 1% increase in windthrow probability for a BA reduction of 1 m2·ha–1). Given that this negative effect of BA on stem windthrow probability is weak, denser stands are expected to exper i e n c e more a bsolute damage even though their relative damage is lower. However, the strength of the rela- tionship (BA vs wind-firmness) may have been influenced by the time that had elapsed between the last partial cut and the windstorm event (5 years). Foster (1988), among others, pointed out the importance of elapsed time since thinning when he found increased damage only in recently thinned stands. Consequently the BA-wind-firmness relationship could have been stronger or weaker if time elapsed since the partial cut had been less or longe r. There have been many studies published (see Table 3) on windthrow in northern hardwood forests, but very few, if any, have discussed how their findings could be used to decrease damage attributable to windthrow. In other parts of the world, however, it is a highly developed research topic (see Savill, 1983), especially in softwood stands. Observing greater and more frequent loss in softwood stands may have influenced the research focus. With the example shown in Figure 9, we dem- onstrated that, when compared to a typical selection cut, a par- tial cut that is designed to harvest less wind-firm species could decrease by about 1 m2·ha–1 the loss that is due to windthrow. This quantity should be viewed as an order of magnitude for mid-intensity storms; in a very high intensity storm (where all the trees fall) or in a very low intensity storm, there would be no difference in damage between typical selection cuts and wind-optimized partial cuts. In Québec, northern hardwood stands generally grow at a rate of about 0.25 m2·year–1 after a commercial selection cut (Forget et al., 2007). This means that preventing a loss of 1 m2·year–1 results in a savings of four years of growth and, in turn, corresponds to about one-sixth of harvest rotation in these stands. Thus, we believe that using a wind-optimized partial cut may be a valuable strategy, espe- cially in parts of the landscape that are more prone to wind- storm damage. The fact that species wind-firmness rankings do not appear to be influenced by wind intensity, as shown by our results, and that they do not appear to be influenced by site factors, as shown by Peterson (2007), should encourage forest managers to adopt such strategies, despite the stochasticity of windstorm effects. Conclusion The bio-indicator method used in this paper for estimating wind intensity was an effective way of isolating the relative effects of species, size, and density on stem windthrow prob- ability, and of verifying the possible interaction among these variables. It remains an indirect method of measuring wind intensity and its real test would require a comparison with anemometer measurements during a windstorm. Unfortunately, such validation is unlikely because it would require knowing in advance where a windstorm would strike to install a network of anemometers. Despite its indirect nature, the method presented is both simple and ecologically sounded. Because of these characteristics, we believe it could be used in other circum- stances and other ecosystems. For instance, many jurisdictions have implemented permanent plot inventories to describe the forests under their governance. If the occurrence of windthrow is tallied during these inventories, plots that experienced them, and which contain a certain number of the same specific spe- cies-DBH combinations, could be used to conduct studies similar to the one presented in this paper. Hence, the method opens the door to the conducting of many regional windthrow studies. Copyright © 2012 SciRes. 85  P. NOLET ET AL. Finally, even though we are confident of the model devel- oped in this study, we have recognized that it is incomplete. From our field experiences, we have identified at least two variables that could improve the model performance; both variables were related to the domino effect discussed earlier. First, we think that the spatial distribution of trees within stands is important. For example, a small SM has a greater chance of being thrown down by another tree if it is close to a few big stems of a low wind-firm species, than if it is close to small stems of a high wind-firm species. Second, while it is obvious that some trees are not thrown down directly by the wind, but indirectly toppled by the fall of other trees, this information was impossible to gather with certainty in the field. Field techniques that would facilitate precise pre-storm tree mapping and stem fall chronologies during the storm should be developed. The information that is obtained could be incorporated into a statis- tical model to increase its performance. The effects of wind- storms on forest stands would probably then appear less sto- chastic. Acknowledgements We are grateful to R. Pouliot, J. Poirier and S. Delagrange for assistance in the field and to T. L. Bidot and W. Parsons for English editing. We also thank C.D. Canham for enlightening discussions during the preparation of the manuscript. Financial support was provided by the Programme de mise en valeur des ressources du milieu forestier (Ministère des Ressources naturelles et de la Faune du Québec). REFERENCES Anderson, D. R., Burnham, K. P., & Thompson, W. L. (2000). Null hypothesis testing: Problems, prevalence, and an alternative. Journal of Wildlife Management, 64, 913-923. doi:10.2307/3803199 Anderson, D. R., Link, W. A., Johnson, D. H., & Burnham, K. P. (2001). Suggestions for presenting the results of data analyses. Journal of Wildlife Management, 65, 373-378. doi:10.2307/3803088 Boose, E. R., Chamberlin, K. E., & Foster, D. R. (2001). Landscape and regional impacts of hurricanes in New England. Ecological Monographs, 71, 27-48. doi:10.1890/0012-9615(2001)071[0027:LARIOH]2.0.CO;2 Bormann, F. H., & Likens, G. E. (1979). Catastrophic disturbance and the steady state in northern hardwood forests. American Scientist, 67, 660-669. Canham, C. D., & Loucks, O. L. (1984). Catastrophic windthrow in the presettlement forests of Wisc onsin. Ecology, 65, 803-809. doi:10.2307/1938053 Canham, C. D., Papaik, M. J., & Latty, E. F. (2001). Interspecific varia- tion in susceptibility to windthrow as a function of tree size and storm severity for northern temperate tree species. Canadian Journal of Forest Research, 31, 1-10. doi:10.1139/x00-124 Clinton, B. D., & Baker, C. R. (2000). Catastrophic windthrow in the southern Appalachians: Characteristics of pits and mounds and initial vegetation responses. Forest Ecology and Management, 126, 51-60. doi:10.1016/S0378-1127(99)00082-1 Doyon, F., Bouchard, A., & Gagnon, D. (1998). Tree productivity and successional status in Québec northern hardwoods. Ecoscience, 5, 222-231. Environment Canada (2 011 ). Climate data online. http://www.climate.weatheroffice.ec.gc.ca/climateData/canada_e.html Everham, E. M. I., & Brokaw, N. V. L. (1996). Forest damage and recovery from catastrophic wind. Botanical Review, 62, 113-185. doi:10.1007/BF02857920 Forget, E., Nolet, P., Doyon, F., Delagrange, S., & Jardon, Y. (2007). Ten-year response of northern hardwood stands to commercial selec- tion cutting in southern Quebec, Canada. Forest Ecology and Man- agement, 242, 764-775. doi:10.1016/j.foreco.2007.02.010 Foster, D. R. (1988). Species and stand response to catastrophic wind in central New England, USA. Journal of Ecology, 76, 135-151 doi:10.2307/2260458. Fukui, D., Hirao, T., Murakami, M., & Hirakawa, H. (2011). Effects of treefall gaps created by windthrow on bat assemblages in a temperate forest. Forest Eco logy and Management, 261, 1546-1552. doi:10.1016/j.foreco.2011.02.001 Hanson, J. J., & Lorimer, C. G. (2007). Forest structure and light re- gimes following moder ate wind storms: Implications for multi-cohort management. Ecological Applications, 17, 1325-1340. doi:10.1890/06-1067.1 Huggard, D. J., Klenner, W., & Vyse, A. (1999). Windthrow following four harvest treatments in an Engelmann spruce—Subalpine fir forest in southern interior British Columbia, Canada. Canadian Journal of Forest Research, 29, 1547-1556. doi:10.1139/x99-135 Johnson, J. B., & Omland , K. S. (2004). Model selection in ecology and evolution. Trends in Ecology & Evolution, 19, 101-108. doi.org/10.1016/j.tree.2003.10.013 Kneeshaw, D. D., Harv ey, B. D., Reyes, G. P., Caro n, M. N ., & Barlo w, S. (2011). Spruce budworm, windthrow and partial cutting: Do dif- ferent partial disturbances produce different forest structures? Forest Ecology and Management, 262, 482-490. doi:10.1016/j.foreco.2011.04.014 GPPC. Le groupe de travail sur les Pédo-paysages du Canada (2010). Pédo-paysage du Canada version 3.2. Agriculture et Agroalimentaire Canada (carte numérique et base de données à l'échelle de 1/1 million). Ni Dhubhain, A., Walshe, J., Bulfin, M., Keane, M., & Mills, P. (2001). The initial development of a windthrow risk model for Sitka spruce in Ireland. Forestry, 74, 161-170. doi:10.1093/forestry/74.2.161 Nolet, P., Delagrange, S., Bouffard, D., Doyon, F., & Forget, E. (2008). The successional status of sugar maple (Acer saccharum), revisited. Annals of Forest Science, 65, 208. doi:10.1051/forest:2007091 Nolet, P., Hartmann, H., Bouffard, D., & Doyon, F. (2007). Predicted and observed sugar maple mortality in relation to site quality indica- tors. Northern Journal of Applied Forestry, 24, 258-264. Papaik, M. J., & Canham, C. D. (2006). Species resistance and com- munity response to wind disturbance regimes in northern temperate forests. Journal of E cology, 94, 1011-1026. doi:10.1111/j.1365-2745.2006.01153.x Papaik, M. J., Canham, C. D., Latty, E. F., & Woods, K. D. (2005). Effects of an introduced pathogen on resistance to natural distur- bance: Beech bark disease and windthrow. Canadian Journal of Forest Research, 35, 1832-1843. doi:10.1139/x05-116 Peltola, H., ki, S., Nen, H., & Ikonen, V. P. (1999). A mechanistic model for assessing the risk of wind and snow damage to single trees and stands of Scots pine, Norway spruce, and birch. Canadian Jour- nal of Forest Research, 29, 647-661. doi:10.1139/x99-029 Peterson, C. J. (2007). Consistent influence of tree diameter and species on damage in nine eastern North America tornado blowdowns. For- est Ecology and Management, 250, 96-108. doi:10.1016/j.foreco.2007.03.013 Putz, F. E., Coley, P. D., Lu, K., Montalvo, A., & Aiello, A. (1983). Uprooting and snapping of trees: Structural determinants and eco- logical consequences. Canadian Journal of Forest Research, 13, 1011-1020. doi:10.1139/x83-133 R Development Core Team (2004). R Development Core Team, R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. Rich, R. L., Frelich, L. E., & Reich, P. B. (2007). Wind-throw mortality in the southern boreal forest: Effects of species, diameter and stand age. Journal of Ecology, 95, 1261-1273. doi:10.1111/j.1365-2745.2007.01301.x Robitaille, A., & Saucier, J.-P. (1998). Paysages régionaux du Québec méridional. Les Publications du Québec, Québec. Ruel, J. C. (1995). Understanding windthrow: Silvicultural implications. The Forestry Chronicle, 71, 435-445. Ruel, J. C. (2000). Factors influencing windthrow in balsam fir forests: Copyright © 2012 SciRes. 86  P. NOLET ET AL. Copyright © 2012 SciRes. 87 From landscape studies to individual tree studies. Forest Ecology and Management, 135, 169-178. doi:10.1016/S0378-1127(00)00308-X Saucier, J.-P., Grondin, P., Robitaille, A., & Bergeron, J. F. (2011). Carte des zones de végétation et des domaines bioclimatiques du Québec. Ministère des Ressources naturelles, de la Faune et des Parcs, Direction des inventaires forestiers. Savill, P. S. (1983). Silviculture in windy climates. Forestry Abstracts, 44, 473-488. Scott, R. E., & Mitchell, S. J. (2005). Empirical modelling of wind- throw risk in partially harvested stands using tree, neighbourhood, and stand attributes. Forest Ecology and Management, 218, 193-209. doi:10.1016/j.foreco.2005.07.012 Seymour, R. S., White, A. S., & DeMaynadier, P. G. (2002). Natural disturbance regimes in northeastern North America—Evaluating sil- vicultural systems using natural scales and frequencies. Forest Ecol- ogy and Management, 155, 357-367. doi:10.1016/S0378-1127(01)00572-2 Simon, A., Gratzer, G., & Sieghardt, M., (2011). The influence of windthrow microsites on tree regeneration and establishment in an old growth mountain forest. Forest Ecology and Management, 262, 1289-1297. doi:10.1016/j.foreco.2011.06.028 Stedinger, J. R. (1984). A Spruce budworm-forest model and its impli- cations for suppression programs. Forest Science, 30, 597-615. Stephens, S. L., & Ruth, L. W. (2005). Federal forest-fire policy in the United States. Ecological Application s, 15, 532-542. doi:10.1890/04-0545 Valinger, E., & Fridman, J. (2011). Factors affecting the probability of windthrow at stand level as a result of Gudrun winter storm in south- ern Sweden. Forest Ecology and Management, 262, 398-403. doi:10.1016/j.foreco.2011.04.004 Wood, C. J. (1995). Understanding wind forces on trees. In M. P. Coutts, & J. Grace (Eds.), Wind and trees (pp. 133-164). Ca mbridge: Cambridge University Pres s. Woods, K. D. (2004). Intermediate disturbance in a late-successional hemlock-northern hardwood forest. Journal of Ecology, 92, 464-476. doi:10.1111/j.0022-0477.2004.00881.x.
|