American Journal of Plant Sciences
Vol.06 No.07(2015), Article ID:55936,15 pages
10.4236/ajps.2015.67107
Heterosis and Combing Ability in F1 Population of Hexaploid Wheat (Triticum Aestivum L.)
Fahad Ali Kalhoro1, Asghar Ali Rajpar1, Shahmir Ali Kalhoro2, Amanullah Mahar3,4, Amjad Ali4*, Sohail Ahmed Otho5, Rab Nawaz Soomro6, Fayaz Ali4, Zulfiqar Ali Baloch7
1Department of Plant Breeding & Genetics, Sindh Agriculture University, Tando Jam, Pakistan 2Faculty of Agriculture, Lasbela University of Agriculture, Water & Marine Sciences, Lasbela, Pakistan
3Centre for Environmental Sciences, University of Sindh, Jamshoro, Pakistan 4College of Natural Resources and Environment, Northwest A&F University, Yangling, China 5College of Plant Protection, Northwest A&F University, Yangling, China 6College of Animal Science and Technology, Northwest A&F University, Yangling, China 7College of Economics and Management Sciences, Northwest A&F University, Yangling, China
Email: *amjadali@aup.edu.pk
Copyright © 2015 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 10 March 2015; accepted 21 April 2015; published 24 April 2015
ABSTRACT
The heterosis, heterobeltiosis, general combining ability (GCA) and specific combining ability (SCA) were examined during Rabi (crop season) of 2013-2014 in hexaploid wheat genotypes. The experiment for this study was conducted at the Botanical Garden, Department of Plant Breeding & Genetics, Sindh Agriculture University Tandojam, Pakistan. The breeding material was comprised of four parents (Imdad, TD-1, SKD-1, and Moomal). These parents were crossed in half partial mating fashion; thus, six possible cross combinations (F1s) were obtained (Imdad × TD-1, Imdad × SKD-1, Imdad × Moomal, TD-1 × SKD-1, TD-1 × Moomal, and SKD-1 × Moomal). A three-replicated RCB Design was used. The mean squares corresponding to different traits of various hexaploid wheat genotypes indicated significant (P < 0.01) GCA (parents) and SCA (F1 hybrids) effects for the characters (plant height, tillers plant−1, spike length, spikelets spike−1, seeds spike−1, seed index, and grain yield plant−1). The mean performance of F1 hybrids differed significantly (P < 0.01) for all the traits studied. Among the parents, Imdad and TD-1 proved to be better general combiners for almost all the studied traits. In regards to SCA effects, the F1 hybrids Imdad × TD-1 and Imdad × SKD-1 expressed higher SCA and heterotic effects for most of the studied traits.
Keywords:
Wheat, Breeding, GCA, SCA, Heterosis, Heterobeltiosis

1. Introduction
Wheat (Tritium aestivum L.) is the most important crop and among the major three cereal crops that provides 20 percent of the total energy requirement in human food [1] . Wheat belongs to the family poaceae, originally from the Levant region of the Near East and Ethiopian Highlands, but now cultivated worldwide [2] . Major cultivated species of wheat include: Triticum aestivum, which is a hexaploid species and is widely cultivated in the world; Triticum durum, the only tetraploid form of wheat widely used today, and the second most widely cultivated wheat; Triticum monococcum, a diploid species with wild and cultivated variants; Triticum dicoccum, a tetraploid species, cultivated in ancient times but no longer has widespread use; Triticum spelta, another hexaploid species, which is cultivated in limited quantities [3] . It is used to make flour for leavened, flat and steamed breads and most of the baked foods, for fermentation to make beer and alcohol [4] . In Pakistan, wheat is averagely used for about 60 percent of daily diet of common men with average per capita consumption of 125 kg [5] .
Pakistan is among top ten wheat producing countries of the world [6] . The contribution of wheat to value addition in agriculture is 10.1 percent, while its contribution to GDP is 2.2 percent [7] . The production of wheat stood at 24.2 million tons during 2012-2013 against the target of 25.5 million tons, showing 5.1 percent decrease, while an increase of 3.2 percent over the last year production of 23.5 million tons. The area under wheat cultivation increased to 8693 thousand hectares in 2012-2013 from 8650 thousand hectares, showing an increase of 0.5 percent over preceding year area under wheat. The production of wheat stood at 24.2 million tons against the target of 25.5 million tons in 2012-2013, showing a decrease, while an increase over the production reported (23.5 million tons) for 2011-2012. The yield per hectare in 2012-2013 remained 2787 kg, showing a positive growth as compared with last year (2011-2012) [7] .
Breeding efforts have resulted in various varieties of hexaploid wheat, having improved yield and grain characters. Varieties and advanced lines with different morphological and economic characteristics are now available as breeding stock. For further progress, knowledge of breeding behavior, particularly of combining ability and type of gene action for the various traits, is necessary. The estimates of combining ability variances and effects can give an indication of the relative magnitude of genetic variance. Besides doing haphazard crosses, it is necessary to execute only high performing ones. In this context, combining ability provides a guideline for selecting elite parents and desirable cross combinations to be used in formulation of a systematic breeding project for rapid improvement. Griffing’s approach has been greatly popular among various breeders, indicating greater role of additive gene action in the inheritance of grain yield and its components [8] . It is also reported that in the inheritance of productive tillering some dominance is involved [9] . It is reported that mean squares for GCA are highly significant for yield and yield components [10] ; SCA is non-significant for all the traits [11] . Significant differences are reported for general combining ability and specific combining ability of all the characters [12] . It is also reported that all investigated features are determined by additive gene action [13] ; the parents LU26S and 4072 are good general combiners [14] . Most of these studies revealed that a large part of total genetic variability for yield and its components was associated with the GCA effects, a measure of additive genetic variance.
Heterosis is a quicker, cheaper and easier method of increasing crop production. With a sufficient level of heterosis, commercial production of hybrid varieties will be justified, and heterotic studies can provide the basis for the exploitation of valuable hybrid combinations in breeding programs. Hybrid wheat technology can play an effective role in enhancing grain production. Although the presence of heterosis in wheat was earlier reported in 1919 by Freeman, its large scale exploitation had not been realized to recent past. Studies on heterosis would help for generating breeding strategies of hybrid wheat production. Obtaining genetic information from diallel cross progenies is a common practice of plant breeders working with autogamous crops. Insight information about the desirable parental combination is a prerequisite in any plant breeding program, focusing on a high degree of heterotic response [15] .
During breeding programs, it is necessary to select pure lines of high general combining ability that indicate the additive gene effect. On the basis of that, predicting progenies and making choice of cross combinations and genotypes can be carried out. Combining ability investigations are carried out by plant breeders to select parents with efficient transferring desirable genes to the progenies. It is reported that general and specific combining ability effects are very important in constructing the next phase of a breeding program [16] [17] . Genetic diversity is one of the key factors for the improvement of many crop plants, including wheat. Evaluation of hybrids for heterosis or combining ability in the field is both expensive and time consuming. For this reason, other parameters such as pedigree information, qualitative and quantitative traits and biochemical data were adapted to study heterosis [18] [19] . In view of the facts stated above, the study was carried out on heterosis and combining ability in hexaploid wheat. The study objectives were to predict the performance of wheat varieties in hybrid combinations through diallel crosses, to work out the heterosis and combining ability in F1 generation, and to evaluate the type of gene action involved in yield and its primary components in hexaploid wheat.
2. Materials and Methods
2.1. Study Area (Geography and Climate)
Tando Jam is small town located at 25.367˚N latitude and 68.367˚E longitude with an elevation of 13 metres (43 ft.); it is located on the east bank of the Indus River and is roughly 150 kilometres (93 mi) away from Karachi, Pakistan. Tando Jam has with warm conditions year-round. The period from mid-April to late June (before the onset of the monsoon) is the hottest of the year, with highs peaking in May at 41.4˚C (106.5˚F). During this time, winds that blow usually bring along clouds of dust, while the breeze that flows at night is more pleasant. Winters are warm, with highs around 25˚C (77˚F), though lows can often drop below 10˚C (50˚F) at night. The highest temperature of 48.5˚C (119˚F) was recorded on 7 June 1991, while the lowest temperature of 1˚C (34˚F) was recorded on 8 February 2012. In recent years Tando Jam has seen great downpours. In February 2003, it received 105 millimeters (4.13 in) of rain in 12 hours, leaving many dead. The years of 2006 and 2007 saw close contenders to this record rain with death tolls estimated in the hundreds. The highest single-day rain total of 250.7 millimeters (9.87 in) was recorded on 12 September 1962, while the wettest month was September 1962, at 286 millimeters (11.26 in).
2.2. Research Design
A field experiment was conducted during Rabi season of 2012-2013 at the Botanical Garden, Department of Plant Breeding and Genetics, Sindh Agriculture University Tandojam, Pakistan located at (25˚25'60''N 68˚31'60''E). Four bread wheat cultivars (Mehran-89, Imdad-2005, TD-1, and TJ-83) were crossed in half diallel fashion. These four wheat varieties have been developed by Pakistan Agriculture Research Centre, Islamabad, Pakistan. These varieties can adapt different ecological conditions and recommended for different areas of Pakistan. The experiment was laid out in a Randomized Complete Block Design with three replications. The basic agronomic practices i.e. soil conditions, water irrigation, weed control and fertilizer utilization were applied according to standard procedures.
The experimental material was comprised of the following wheat genotypes:
A: Parents
1) Mehran-89;
2) Imdad-2005;
3) TD-1;
4) TJ-83.
B: Cross combinations
1) Mehran-89 × Imdad-2005;
2) Mehran-89 × TD-1;
3) Mehran-89 × TJ83;
4) Imdad-2005 × TD-1;
5) Imdad-2005 × TJ-83;
6) TD-1 × TJ-83.
The row to row and plant to plant space of 30 cm and 15 cm was kept, respectively. Drilling method was used for sowing and after four weeks of sowing thinning was done to maintain 15 cm plant to plant space. Ten plants of each genotype from each replication selected at random were tagged for record the data in the field and laboratory.
The height of each index plant was measured in the centimeters at the time of maturity from the surface of soil to the tip of the each head excluding awns. The number of fertile tillers in each tagged plants per replication per genotype were counted at the time of crop harvest and were recorded as number of tillers plant−1. Length of spike from primary tiller was measured in centimeter from the neck node to the apex of terminal spikelet excluding awns. The number of spikelets in each spike from indexed plants was counted. One hundred grains were randomly taken from each index plant and were weighed in grams with electric balance. After harvesting, each plant was threshed separately with single plant wheat thresher. The grains were weighed with electric balance and yield plant−1 in g was recorded.
2.3. Statistical Analysis
The data were subjected to analysis of variance (ANOVA) technique using MSTAT, a computer software package for all the traits studied to test the null hypothesis of no differences between various F1s population and their parental lines. The significant genotypic differences permit the use of simple additive-dominance model. Analysis of variance will be run on the means of the selected plants for above mentioned traits using standard method of [20] . Combining ability analysis will be done by using Method I, and Model II [21] .
2.4. Genetic Analysis
The diallel technique was adapted for genetic analysis given in “Biometrical Methods in Quantitative Genetic Analysis”.
2.5. Analysis for Heterosis and Heterobeltiosis
Heterosis was calculated in terms of percent increase (+) or decrees (−) of the F1 and F2 hybrids against its mid parent value as suggested [22] .
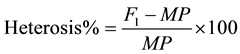
where:
F1 = Mean of the F1 cross;
MP = Mid parent.
The term heterobeltiosis, coined [23] , was also estimated in terms of percent increase or decrease of the F1 and F2 hybrid over its better parent.
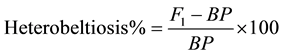
The data were further subjected to “t” test to determine whether F1 and F2 hybrid means are statically different from mid and better parental values. The “t” values was calculated by the following formula as used [24] .
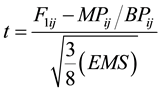
where:
F1ij = Mean of the ijth F1 cross;
MPij = Mid parent value for the ijth cross;
BPij = Better parent value for the ijth cross;
Combining ability analysis:
General combining ability effect (GCA):
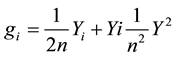
where:
gi = General combining ability effects for line I;
n = Number of present/varieties;
Yi = Total of mean values of F1s resulting from crossing ith lines with jth lines;
Y2 = Grand total of all the mean values in the table.
Specific combining ability affects (SCA):
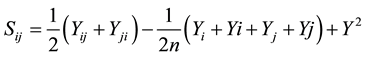
where:
Sij = Specific combining ability between ith and jth lines;
Yij = Mean value of the F1 resulting from crossing the ith and jth lines;
Yji = Mean value for F1 resulting from crossing the jth and ith varieties;
Yi = Total of mean values of F1’s resulting from crossing jth line with ith varieties;
Yi = Reciprocal values of Yi;
Yj = Total values for F1’s resulting from crossing the ith line with jth line;
Yj = Values of reciprocal F1’s of Yj;
Y2 = Grand total of the observations.
3. Results and Discussion
In order to examine the heterosis, heterobeltiosis, general combining ability (GCA) and specific combining ability (SCA) in hexaploid wheat genotypes for different characteristics such as: plant height, tillers plant−1, spike length, spikelets spike−1, seeds spike−1, seed index and grain yield plant−1. Four parents were crossed in half partial mating fashion; thus six possible cross combinations (F1s) were achieved. The F1 hybrids were evaluated by analysis of variance (A
3.1. ANOVA
The mean squares from the analysis of variance table corresponding to various growth and yield contributing traits of wheat genotypes indicated that the performance of different genotypes for traits studied varied significantly (P < 0.01). It was further found that the SCA of crosses and GCA of parents were significantly (P < 0.01) different for all the characters studied (plant height, tillers plant−1, spikelets spike−1, seeds spike−1, seed index and grain yield plant−1). The results indicated that the data for all the traits examined is useful and can be analyzed further to estimate the general and specific combining ability effects.
3.2. Mean Performance
The mean performance of parents and F1 hybrids of seven traits (plant height, tillers plant−1, spike length, spike

Table 1. Mean squares corresponding to diallel analysis for various characters in bread wheat.
**Highly significant (P < 0.01); *Significant (P < 0.05).
Table 2. Mean performance of parents and their F1 hybrids for agro-economic traits of hexaploid wheat.
Table 3. Heterotic effects of six F1 hybrids over mid parents (relative heterosis) and better parents (heterobeltiosis) for the character plant height.
Table 4. Heterotic effects of six F1 hybrids over mid parents (relative heterosis) and better parents (heterobeltiosis) for the character tillers per plant.
Table 5. Heterotic effects of six F1 hybrids over mid parents (relative heterosis) and better parents (heterobeltiosis) for the character spike length.
Table 6. Heterotic effects of six F1 hybrids over mid parents (relative heterosis) and better parents (heterobeltiosis) for the character spikelets per spike.
Table 7. Heterotic effects of six F1 hybrids over mid parents (relative heterosis) and better parents (heterobeltiosis) for the character seeds per spike.
Table 8. Heterotic effects of six F1 hybrids over mid parents (relative heterosis) and better parents (heterobeltiosis) for the character seed index.
Table 9. Heterotic effects of six F1 hybrids over mid parents (relative heterosis) and better parents (heterobeltiosis) for the character grain yield per plant.
Table 10. General combining ability (GCA) estimates of parents.
lets spike−1, seeds spike−1, seed index/100 seed weight and grain yield plant−1) were studied and the data (Table 2) indicated that among the parents the maximum plant height (83.00 cm) was recorded in parent Imdad, and the minimum plant height (59.67 cm) was obtained in parent TD-1. Among the F1 hybrids, the maximum plant height (77.93 cm) was recorded in cross combination of Imdad × Moomal; while in cross combinations Imdad × SKD-1, SKD-1 × Moomal and TD-1 × Imdad produced plants of 73.93, 72.93 and 68.27 cm, respectively. However, while the minimum plant height (57.93 cm) was recorded in cross combination of TD-1 × SKD-1.
In case of tillering capacity in parents and their F1 hybrids, the highest values among the parents for this trait (14.33 plant−1) was obtained TD-1 and minimum (9.87 plant−1) in Moomal; while in F1 hybrids, the highest number of tillers plant−1 (17.60) was recorded in cross combination of Imdad × TD-1, followed by cross combinations of Imdad × SKD-1 (14.80 plant−1) and TD-1 × SKD-1 (14.07 plant−1); while the minimum number of tillers was observed in cross combination of SKD-1 × Moomal (11.60 plant−1). Similarly, the spike length among the parents was higher i.e. 15.87 cm and 15.47 cm in parents Imdad and TD-1; while minimum (11.87 cm) in Moomal. In F1 hybrids, maximum spike length (16.93 cm) was observed in cross combination of Imdad × TD-1, followed by TD-1 × Moomal (16.33 cm) and Imdad × SKD-1 (14.87 cm) while the minimum spike length (11.60 cm) was observed in cross combination of SKD-1 × Moomal.
In case of the trait spikelets spike−1, the highest value among parents was observed in SKD-1 (20.33) and the lowest equally in TD-1 and Moomal (18.33). In F1 hybrids, the maximum number of spikelets (20.60 spike−1) was obtained in the cross combination of Imdad × TD-1; followed by cross combinations of Imdad × SKD-1 (19.93), Imdad × Moomal (19.67) and SKD-1 × Moomal (19.00); while the lowest number of spikelets (17.67) spike−1 was equally noted in case of crosses TD-1 × SKD-1 and TD-1 × Moomal.
The data on the number of seeds spike−1 indicate that the highest value among parents for this trait was observed in Moomal (69.67) and the lowest seeds spike−1 was noted in TD-1 (63.80). In F1 hybrids, the maximum number of seeds (76.27 spike−1) was noted in the cross combination of Imdad × TD-1; followed by cross combinations of Imdad × SKD-1 (73.67) and Imdad × Moomal (71.93); while the lowest number of seeds (62.40) spike−1 was noted in case of cross SKD-1 × Moomal. The seed index value on the basis of 1000 seeds weight among parents was highest (50.80 g) in TD-1 and lowest (45.20 g) in Moomal. Among F1 hybrids the highest seed index (52.00 g) was achieved in case of cross combinations of Imdad × TD-1, followed by Imdad × SKD-1 (50.00 g) and Imdad × Moomal (48.00 g); while the lowest seed index value (44.53 g) was obtained in case of the cross combination SKD-1 × Moomal.
For the character grain yield plant−1, among parents the yield plant−1 was highest (29.33 g) in SKD-1 and minimum grain yield (25.40 g) was achieved in genotype Moomal. In F1 hybrids, the grain yield plant−1 was highest (30.87 g) in the cross combination of Imdad × TD-1, followed by the cross combinations of Imdad-2005 × SKD-1 and TD-1 × SKD-1 with average grain yield of 29.27 g and 27.20 g, respectively. However, the lowest seed yield plant−1 (24.87 g) was obtained in cross combination of SKD-1 × Moomal.
3.3. Heterosis and Heterobeltiosis
3.3.1. Plant Height (cm)
Heterosis and heterobeltiosis studies on plant height of F1 hybrids of hexaploid wheat (Table 3) showed that all the F1 hybrids recorded negative heterotic effects for plant height and the cross Imdad × TD-1 resulted in maximum negative heterosis (14.39%) for plant height; and maximum heterobeltiosis estimate (26.43%) was also recorded in the same cross i.e. Imdad × TD-1 in negative sense for plant height among F1 hybrids.
3.3.2. Number of Tillers Plant−1
The estimation of heterotic effect calculated for the number of tillers plant−1 presented in Table 4 showed that all the F1 hybrids recorded positive heterotic effects for the number of tillers plant−1. The three top scoring F1 hybrids were Imdad × TD-1, Imdad × SKD-1 and Imdad × Moomal with relative heterosis of 28.78, 21.64 and 13.70 percent, respectively for the number of tillers plant−1; while the three top scorer F1 hybrids for heterobeltiosis were Imdad × TD-1, TD-1 × Moomal and SKD-1 × Moomal with 35.38, 27.03 and 20.27% heterobeltiosis for the number of tillers plant−1, respectively.
3.3.3. Spike Length (cm)
There were diverse heterotic effects for spike length as three F1 hybrids showed positive heterotic effects and equally three F1 hybrids displayed negative heterotic effects. The three top scoring F1 hybrids were for positive heterotic effect on spike length were TD-1 × Moomal, Imdad × TD-1 and Imdad × SKD-1with relative heterosis of 17.79, 8.09 and 5.94 percent, respectively; while the three top scorer F1 hybrids for heterobeltiosis were TD-1 × Moomal, Imdad × Moomal and Imdad × TD-1 with 37.64%, 14.61% and 9.48% heterobeltiosis for the character spike length in hexaploid wheat.
3.3.4. Number of Spikelets Spike−1
The heterotic effects of number of spikelets spike−1 presented in Table 6 indicated that two F1 hybrids displayed positive heterosis and four F1 hybrids showed negative heterotic effect for number of spikelets spike−1 character. The positive heterosis was displayed by cross combinations of Imdad × TD-1 (8.42%) and Imdad × Moomal (3.51%); and the negative heterosis was displayed by cross combinations of TD-1 × SKD-1 (−8.62%) and TD-1 × Moomal (−3.64%). The top high scorer F1 hybrids for positive heterobeltiosis were Imdad × Moomal, Imdad × TD-1 and SKD-1 × Moomal with 7.27%, 4.75% and 3.64% heterobeltiosis for the number of spikelets spike−1, respectively. Negative heterobeltiosis (−3.64% and −3.64%) was displayed by the cross combinations of TD × SKD-1 and TD-1 × Moomal.
3.3.5. Number of Seeds Spike−1
The heterotic effects of number of seeds spike−1 presented in Table 7 showed that out of six, four F1 hybrids showed positive heterosis and two F1 hybrids displayed negative heterosis for number of seeds spike−1 character. The positive heterosis was displayed by cross combinations of Imdad × TD-1 (19.04%), Imdad × SKD-1 (14.09%), Imdad × Moomal (7.36%), TD-1 × SKD-1 (8.86%); and the negative heterosis was displayed by cross combinations of TD-1 × Moomal (−0.30%) and SKD-1 × Moomal (−7.19%). The top high scorer F1 hybrids for positive heterobeltiosis were Imdad × ID-1, Imdad × SKD-1 and TD-1 × SKD-1 with 18.55%, 14.51% and 9.72% heterobeltiosis for the number of seeds spike−1, respectively. Negative heterobeltiosis (−10.50% and −4.50%) was displayed by the cross combinations of SKD-1 × Moomal and TD-1 × Moomal.
3.3.6. Seed Index (1000 Grains Weight g)
The study results on the heterotic effects of seed index value for six F1 hybrids are given in Table 8 which indicated that out the six F1 hybrids, three F1 hybrids showed positive heterotic and heterobeltiosis effects and equally three hybrids displayed negative heterosis and heterobeltiosis for seed index character. The higher relative heterosis was displayed by cross combinations of Imdad × TD-1 (5.12%) and Imdad × SKD-1 (5.41%); while the higher negative heterosis was displayed by the cross combinations of TD-1 × SKD-1 (7.72%) and TD-1 × Moomal (6.25%). The top high scorer F1 hybrids for heterobeltiosis were Imdad × TD-1, Imdad × Moomal and Imdad × SKD-1 with 8.03%, 6.19% and 3.88% heterobeltiosis for the seed index, respectively.
3.3.7. Seed Yield Plant−1 (g)
The results in regards to heterotic effects for seed yield plant−1 in six F1 hybrids are presented in Table 9 which suggested that out of six F1 hybrids in hexaploid wheat, three F1 hybrids showed positive heterotic effects and remaining three F1 hybrids displayed negative heterosis and heterobeltiosis for seed yield plant−1. The higher relative heterosis was displayed by cross combinations of Imdad × TD-1 (10.37%) and Imdad × SKD-1 (10.16%); while the lower relative heterosis was displayed by the cross combination of Imdad × Moomal (3.59%) for seed yield plant−1. The top high scorer F1 hybrids for heterobeltiosis were Imdad × TD-1, Imdad × SKD-1 and Imdad × Moomal with 16.04%, 10.03% and 6.04% heterobeltiosis for the seed yield plant−1, respectively.
3.4. Analysis for General Combining Ability (GCA) and Specific Combining Ability (SCA)
The mean squares from the analysis of variance (Table 1) indicated significant (P < 0.01) results for GCA and SCA in parents as well as their F1 hybrids. The data in relation to GCA in four parents for various traits of economic importance including yield plant−1 are presented in Table 10; while the results for SCA in the F1 hybrids for different traits are presented in Table 11. The results for GCA and SCA are interpreted as under:
3.4.1. Plant Height (cm)
The results in regards to GCA effects as exhibited by Table 10 on mean plant height of hexaploid wheat illustrate that among the parents; most positive GCA effects were produced by Moomal (4.65) and Imdad (4.55); while parent TD-1 expressed most negative GCA effects of −7.34. In respect to SCA effects (Table 11), the F1 hybrids SKD-1 × Moomal, TD-1 × Moomal and Imdad × SKD-1 demonstrated positive SCA effects of 1.02, 0.83 and 0.12, respectively; while the hybrids Imdad × TD-1, TD-1 × SKD-1 and Imdad × Moomal demonstrated the negative SCA effects of −6.27, −2.99 and −1.39, respectively.
3.4.2. Number of Tillers Plant−1
The GCA effects presented in Table 10 on mean number of tillers plant−1 of hexaploid wheat revealed that among the parents, TD-1 and Imdad demonstrated maximum positive GCA effects of 1.11 and 0.87; whereas the negative GCA effects were expressed by the parents Moomal (−1.51) and SKD-1 (−0.47). In case of SCA effects, (Table 11), the F1 hybrids Imdad × TD-1 demonstrated maximum positive CGA effects (2.38), and hybrid Imdad × SKD-1 ranked second for positive SCA effects (1.16); while hybrid TD-1 × Moomal demonstrated negative SCA effects of 0.31 for the number of tillers plant−1.
Table 11. Specific combining ability (SCA) estimates of F1 hybrids.
3.4.3. Spike Length (cm)
The data shown in Table 10 in regards to spike length of hexaploid wheat indicated that among the parents, TD-1 and Imdad expressed positive GCA effects of 1.23 and 0.81, respectively; whereas parents Moomal and SKD-1 exhibited negative GCA effects of −1.04 and −0.99, respectively. From diallel crosses, three hybrids i.e. TD-1 × Moomal (1.84), Imdad × SKD-1 (0.76) and Imdad × TD-1 (0.60) manifested positive SCA effects; while other three hybrids SKD-1 × Moomal (−0.67), TD-1 × SKD-1 (−0.67) and Imdad × Moomal (−0.47) demonstrated negative SCA effects on the spike length.
3.4.4. Number of Spikelets Spike−1
The results for GCA effects presented in Table 10 regarding number of spikelets spike−1 of hexaploid wheat indicated that among the parents, Imdad and SKD-1 produced positive GCA effects of 0.66 and 0.28, respectively; whereas parents Moomal and SKD-1 exhibited negative GCA effects of −0.50 and −0.43, respectively. In case of F1 hybrids, three hybrids i.e. Imdad × TD-1 (1.32), Imdad × Moomal (0.32) and SKD-1 × Moomal (0.04) manifested positive SCA effects; while other three hybrids TD-1 × SKD-1 (−1.23), TD-1 × Moomal (-0.52) and Imdad × SKD-1 (−0.12) demonstrated negative SCA effects on the number of spikelets spike−1.
3.4.5. Number of Seeds Spike−1
The GCA effects tabulated as Table 10 pertaining to number of seeds spike−1 of hexaploid wheat demonstrated that among the parents, Imdad expressed positive GCA effects of 1.47; whereas parents SKD-1, Moomal and TD-1 demonstrated negative GCA effects of −1.01, −0.25 and −0.22, respectively. In case of F1 hybrids, four F1 hybrids including Imdad × TD-1 (6.67), Imdad × SKD-1 (4.86), TD-1 × SKD-1 (2.88) and Imdad × Moomal (2.37) expressed positive SCA effects; while remaining two F1 hybrids SKD-1 × Moomal (−4.68) and TD-1 × Moomal (−1.34) illustrated negative SCA effects on the number of seeds spike−1 trait.
3.4.6. Seed Index (g)
It is obvious from the results presented in Table 10 that among the parents, Imdad and TD-1 expressed the positive GCA effects of 1.43 and 0.98, respectively for seed index value; whereas the other parent including Moomal (−1.63) and SKD-1 (−0.78) displayed negative GCA effects for seed index value. From six diallel crosses of hexaploid wheat, three F1 hybrids manifested positive SCA effects while other three gave negative SCA effects. The three top scoring hybrids were Imdad × TD-1 (2.05), Imdad × SKD-1 (1.82) and Imdad × Moomal (0.66). However, the negative SCA effects were manifested by hybrids TD-1 × SKD-1 (−2.74), TD-1 × Moomal (−1.90) and SKD-1 × Moomal (−0.60).
3.4.7. Yield Plant−1 (g)
The GCA effects tabulated as Table 10 indicated that TD-1 and Imdad were the parents which expressed positive GCA effects of 1.07 and 0.59, respectively; while Moomal (−1.28) and SKD-1 (−0.38) showed negative GCA effects for grain yield plant−1. The data in Table 11 revealed that the three diallel crosses displayed positive SCA effects, and other three produced negative SCA effects. However, the three top scoring hybrids for grain yield plant−1 were Imdad × TD-1 (1.86), Imdad × SKD-1 (1.72) and Imdad × Moomal (0.28). The negative SCA effects were manifested by hybrids TD-1 × SKD-1 (−0.83), SKD-1 × Moomal (−0.81) and TD-1 × Moomal (−0.73).
The heterosis, heterobeltiosis, GCA and SCA of hexaploid wheat genotypes were examined during the Rabi season of 2013-14 for various traits of economic importance such as: plant height, tillers plant−1, spike length, spikelets spike−1, seeds spike−1, seed index and grain yield plant−1. Four parents (Imdad, TD-1, SKD-1, Moomal) were crossed in half partial mating fashion; thus six possible cross combinations (F1s) were obtained (Imdad×TD-1, Imdad×SKD-1, Imdad × Moomal, TD-1 × SKD-1, TD-1 × Moomal and SKD-1 × Moomal). The results of the study are discussed as follows:
3.5. Heterosis and Heterobeltiosis
3.5.1. Plant Height (cm)
All the F1 hybrids recorded negative heterotic effects for plant height and the cross Imdad × TD-1 resulted in maximum negative heterosis (14.39%) for plant height; and maximum heterobeltiosis estimate (26.43%) was also recorded in the same cross i.e. Imdad × TD-1 in negative sense for plant height among F1 hybrids. These results are further confirmed by [24] and also indicated similar trend of heterosis for plant height of various wheat genotypes and their hybrids [25] [26] .
3.5.2. Number of Tillers Plant−1
The three top scoring F1 hybrids were Imdad × TD-1, Imdad × SKD-1 and Imdad × Moomal with relative heterosis of 28.78, 21.64 and 13.70 percent, respectively for the number of tillers plant−1; while the three top scorer F1 hybrids for heterobeltiosis were Imdad × TD-1, TD-1 × Moomal and SKD-1 × Moomal with 35.38%, 27.03% and 20.27% heterobeltiosis for the number of tillers plant−1, respectively. The above results are in accordance with those of [23] , also stated that experienced positive and significant amount of heterotic effects and heterobeltiosis for number of tillers plant−1 in wheat genotypes [27] - [29] .
3.5.3. Spike Length (cm)
There were diverse heterotic effects for spike length as three F1 hybrids showed positive heterotic effects and equally three F1 hybrids displayed negative heterotic effects. The three top scoring F1 hybrids were for positive heterotic effect on spike length were TD-1 × Moomal, Imdad × TD-1 and Imdad × SKD-1with relative heterosis of 17.79%, 8.09% and 5.94%, respectively; while the three top scorer F1 hybrids for heterobeltiosis were TD-1 × Moomal, Imdad × Moomal and Imdad × TD-1 with 37.64%, 14.61% and 9.48% heterobeltiosis for the character spike length in hexaploid wheat. These results are in agreement with the findings reported by [30] and found high heterotic effects and heterobeltiosis in F1 hybrids of wheat for spike length [31] .
3.5.4. Number of Spikelets Spike−1
Two F1 hybrids displayed positive heterosis and four F1 hybrids showed negative heterotic effect for number of spikelets spike−1 character. The positive heterosis was displayed by cross combinations of Imdad × TD-1 (8.42%) and Imdad × Moomal (3.51%); and the negative heterosis was displayed by cross combinations of TD-1 × SKD- 1 (−8.62%) and TD-1 × Moomal (-3.64%). The top high scorer F1 hybrids for positive heterobeltiosis were Imdad × Moomal, Imdad × TD-1 and SKD-1 × Moomal with 7.27%, 4.75% and 3.64% heterobeltiosis for the number of spikelets spike−1, respectively. Negative heterobeltiosis (−3.64% and −3.64%) was displayed by the cross combinations of TD × SKD-1 and TD-1 × Moomal. Similar results have also been reported positive heterosis for various F1 hybrids in wheat for spikelets per spike [32] - [34] .
3.5.5. Number of Seeds Spike−1
The positive heterosis was displayed by cross combinations of Imdad × TD-1 (19.04%), Imdad × SKD-1 (14.09%), Imdad × Moomal (7.36%), TD-1 × SKD-1 (8.86%); and the negative heterosis was displayed by cross combinations of TD-1 × Moomal (−0.30%) and SKD-1 × Moomal (−7.19%). The top high scorer F1 hybrids for positive heterobeltiosis were Imdad × ID-1, Imdad × SKD-1 and TD-1 × SKD-1 with 18.55%, 14.51% and 9.72% heterobeltiosis for the number of seeds spike−1, respectively. Negative heterobeltiosis (−10.50% and −4.50%) was displayed by the cross combinations of SKD-1 × Moomal and TD-1 × Moomal. It is also reported diversified heterotic effects of parents and their F1 hybrids for seeds spike−1 [35] [36] .
3.5.6. Seed Index (100 Grains Weight g)
The higher relative heterosis was displayed by cross combinations of Imdad × TD-1 (5.12%) and Imdad × SKD- 1 (5.41%); while the higher negative heterosis was displayed by the cross combinations of TD-1 × SKD-1 (7.72%) and TD-1 × Moomal (6.25%). The top high scorer F1 hybrids for heterobeltiosis were Imdad × TD-1, Imdad × Moomal and Imdad × SKD-1 with 8.03%, 6.19% and 3.88% heterobeltiosis for the seed index, respectively. These results are in concurrence with and achieved significant variation in relative heterosis by different cross combinations of wheat genotypes for seed index value [37] .
3.5.7. Seed Yield Plant−1 (g)
The higher relative heterosis was displayed by cross combinations of Imdad × TD-1 (10.37%) and Imdad × SKD-1 (10.16%); while the lower relative heterosis was displayed by the cross combination of Imdad × Moomal (3.59%) for seed yield plant−1. The top high scorer F1 hybrids for heterobeltiosis were Imdad × TD-1, Imdad × SKD-1 and Imdad × Moomal with 16.04%, 10.03% and 6.04% heterobeltiosis for the seed yield plant−1, respectively. The results of the present research are in accordance and reported significant variation in the heterosis among cross combinations for seed yield in wheat breeding programs [38] .
3.6. General and Specific Combining Ability Analysis
3.6.1. Plant Height (cm)
The GCA effects on plant height of hexaploid wheat illustrate that among the parents; most positive GCA effects were produced by Moomal (4.65) and Imdad (4.55); while parent TD-1 expressed most negative GCA effects of −7.34. In respect to SCA effects (Table 11), the F1 hybrids SKD-1 × Moomal, TD-1 × Moomal and Imdad × SKD-1 demonstrated positive SCA effects of 1.02, 0.83 and 0.12, respectively; while the hybrids Imdad × TD-1, TD-1 × SKD-1 and Imdad × Moomal demonstrated the negative SCA effects of −6.27, −2.99 and −1.39, respectively. Positive GCA effects among parents and F1 hybrids have also been reported in their respective breeding studies on wheat genotypes [23] [24] [26] [31] .
3.6.2. Number of Tillers Plant−1
The GCA effects on mean number of tillers plant−1 of wheat revealed that among the parents, TD-1 and Imdad demonstrated maximum positive GCA effects of 1.11 and 0.87; whereas the negative GCA effects were expressed by the parents Moomal (−1.51) and SKD-1 (−0.47). In case of SCA effects (Table 11), the F1 hybrids Imdad × TD-1 demonstrated maximum positive CGA effects (2.38), and hybrid Imdad × SKD-1 ranked second for positive SCA effects (1.16); while hybrid TD-1 × Moomal demonstrated negative SCA effects of 0.31 for the number of tillers plant−1. The results of the present study in regards to tillers per plant are in agreement with and found positive GCA effects for parents and high SCA effects for hybrids in wheat [39] - [41] .
3.6.3. Spike Length (cm)
Among the parents, TD-1 and Imdad expressed positive GCA effects of 1.23 and 0.81, respectively; whereas parents Moomal and SKD-1 exhibited negative GCA effects of −1.04 and −0.99, respectively. From diallel crosses, three hybrids i.e. TD-1 × Moomal (1.84), Imdad × SKD-1 (0.76) and Imdad × TD-1 (0.60) manifested positive SCA effects; while other three hybrids SKD-1 × Moomal (−0.67), TD-1 × SKD-1 (−0.67) and Imdad × Moomal (−0.47) demonstrated negative SCA effects on the spike length. The results of the present study are further in line with [26] [29] [30] .
3.6.4. Number of Spikelets Spike−1
The GCA effects on number of spikelets spike−1 showed that among parents, Imdad and SKD-1 produced positive GCA effects of 0.66 and 0.28, respectively; whereas parents Moomal and SKD-1 exhibited negative GCA effects of -0.50 and −0.43, respectively. In case of F1 hybrids, three hybrids i.e. Imdad × TD-1 (1.32), Imdad × Moomal (0.32) and SKD-1 × Moomal (0.04) manifested positive SCA effects; while other three hybrids TD-1 × SKD-1 (−1.23), TD-1 × Moomal (−0.52) and Imdad × SKD-1 (−0.12) demonstrated negative SCA effects on the number of spikelets spike−1. The results are in similarity with the findings of the present research for the number of spikelets per spike [42] .
3.6.5. Number of Seeds Spike−1
The GCA effects pertaining to number of seeds spike−1 demonstrated that among the parents, Imdad expressed positive GCA effects of 1.47; whereas parents SKD-1, Moomal and TD-1 demonstrated negative GCA effects of −1.01, −0.25 and −0.22, respectively. In case of F1 hybrids, four F1 hybrids including Imdad × TD-1 (6.67), Imdad × SKD-1 (4.86), TD-1 × SKD-1 (2.88) and Imdad × Moomal (2.37) expressed positive SCA effects; while remaining two F1 hybrids SKD-1 × Moomal (−4.68) and TD-1 × Moomal (−1.34) illustrated negative SCA effects on the number of seeds spike−1 trait. These results are further confirmed and diversified GCA effects and SCA effects and expressed the genetic makeup of parental material plays major role to develop positive combining abilities among parents and their crosses [43] [44] .
3.6.6. Seed Index (g)
Among the parents, Imdad and TD-1 expressed the positive GCA effects of 1.43 and 0.98, respectively for seed index value; whereas the other parent including Moomal (−1.63) and SKD-1 (−0.78) displayed negative GCA effects for seed index value. From six diallel crosses of hexaploid wheat, three F1 hybrids manifested positive SCA effects while other three gave negative SCA effects. The three top scoring hybrids were Imdad × TD-1 (2.05), Imdad × SKD-1 (1.82) and Imdad × Moomal (0.66). However, the negative SCA effects were manifested by hybrids TD-1 × SKD-1 (−2.74), TD-1 × Moomal (−1.90) and SKD-1 × Moomal (−0.60). These findings are further confirmed and were of the conclusion that F1 hybrids manifested positive SCA for seed index in wheat genotypes in their breeding programs [31] [45] .
3.6.7. Yield Plant−1 (g)
The parent TD-1 and Imdad expressed positive GCA effects of 1.07 and 0.59, respectively; while Moomal (−1.28) and SKD-1 (−0.38) showed negative GCA effects for grain yield plant−1. The data in Table 11 revealed that the three diallel crosses displayed positive SCA effects, and other three produced negative SCA effects. However, the three top scoring hybrids for grain yield plant−1 were Imdad × TD-1 (1.86), Imdad × SKD-1 (1.72) and Imdad × Moomal (0.28). The negative SCA effects were manifested by hybrids TD-1 × SKD-1 (−0.83), SKD-1 × Moomal (−0.81) and TD-1 × Moomal (−0.73). Similar results in regards to yield per plant of wheat for SCA among F1 hybrids have also been reported [46] -[49] .
4. Conclusion
The heterosis, heterobeltiosis, general combining ability (GCA) and specific combining ability (SCA) were examined in hexaploid wheat genotypes for various traits of economic importance, including plant height, tillers plant−1, spike length, spikelets spike−1, seeds spike−1, seed index, and grain yield plant−1. Four parents (Imdad, TD-1, SKD-1, and Moomal) were crossed in half partial mating fashion; thus, six possible cross combinations (F1s) were obtained (Imdad × TD-1, Imdad × SKD-1, Imdad × Moomal, TD-1 × SKD-1, TD-1 × Moomal, and SKD-1 × Moomal). The F1 hybrids were statistically analyzed using analysis of variance (A
References
- Shewry, P.R. (2009) Wheat. Journal of Experimental Botany, 60, 1537-1553. http://dx.doi.org/10.1093/jxb/erp058
- Belderok, B. (2000) Bread-Making Quality of Wheat: A Century of Breeding in Europe. Kluwer Academic Publishers, Netherlands, 34. http://dx.doi.org/10.1007/978-94-017-0950-7
- Moon, D. (2008) In the Russian Steppes: The Introduction of Russian Wheat on the Great Plains of the United States of America. Journal of Global History, 3, 203-225. http://dx.doi.org/10.1017/S1740022808002611
- Tsenov, N., Atanasova, D., Todorov, I. and Dochev, V. (2008) Environmental Effect on Common Winter Wheat Productivity. Proceedings of 18th EUCARPIA General Congress, Valencia, 9-12 September 2008, 480-484.
- Khan, A.K., Salim, I. and Ali, Z. (2003) Heritability of Various Morphological Traits in Wheat. International Journal of Agriculture and Biology, 5, 138-140.
- Ihsanullah, Jan, A., Taj, F.H., Khan, I.A. and Khan, N. (2002) Effect of Sowing Dates on Yield and Yield Components of Mash Bean. Asian Journal of Plant Sciences, 1, 622-624. http://dx.doi.org/10.3923/ajps.2002.622.624
- GOP (2013) Agriculture: Economic Survey of Pakistan 2013-2014. Ministry of Food and Agriculture, Government of Pakistan, Islamabad.
- Parashar, R.R. and Janoria, M.P. (1998) Combining Ability over Environments in Wheat. Jawaharlal Nehru Krishi Vishwavidyalaya (JNKVV) Res., 28-29, 34-39.
- Lonc, W. and Zalewski, D. (1996) Combining Ability of Seven Lines of Winter Wheat. Biuletyn Inst. Hodowli- i-Akli- matyzacji Roslin, 200, 259-265.
- Ali, Z. and Khan, A.S. (1998) Combining Ability Studies of Some Morphophysiological Traits in Bread Wheat (Triticum aestivum L). Pakistan Journal of Agricultural Sciences, 35, 1-4.
- Kakar, A.A., Larik, A.S., Kumbhar, M.B., Anwar, M.S. and Naz, M.A. (1999) Estimation of Heterosis, Potence Ratio and Combining Ability in Bread Wheat (Triticum aestivum L). Pakistan Journal of Agricultural Sciences, 36, 169-174.
- Sheikh, S. and Singh, I. (2000) Combining Ability Analysis in Wheat Plant Characters and Harvest Index. International Journal of Tropical Agriculture, 18, 29-37.
- Zalewski, D. (2001) Estimation of Combining Ability of Seven Winter Wheat Lines. Biuletyn Inst. Hodowli-i-Akli- matyzacji Roslin, 218-219, 75-81.
- Mahmood, N. and Chowdhry, M.A. (2002) Ability of Bread Wheat Genotypes to Combine for High Yield under Varying Sowing Conditions. Journal of Genetics and Breeding, 56, 119-125.
- Hassan, G., Muhammad, F., Khalil, F.H. and Raziuddin. (2006) Heterosis and Heterobeltiosis Studies for Morphological Traits in Bread Wheat. Sarhad Journal of Agriculture, 22, 51-54.
- Madic, M., Paunovic, A., Durovic, D., Kraljvic-Balalic, M. and Knezevic, D. (2005) The Analysis of Gene Effect in the Inheritance of Kernel Number Per Spike in Barley Hybrid. Genetika, 37, 261-269. http://dx.doi.org/10.2298/GENSR0503261M
- Singh, R., Agarwal, M. and Shrotria, P.K. (2005) Combining Ability Analysis for Forage Yield and Quality Characters in Sorghum. Forage Research, 31, 8-11.
- Wang, Z.Y., Second, G. and Tanksley, S.D. (1992) Polymorphism and Phylogenetic Relationships among Species in the Genus Oryza as Determined by Analysis on Nuclear RFLPs. Theoretical and Applied Genetics, 83, 565-581. http://dx.doi.org/10.1007/BF00226900
- El-Maghraby, M.A., El-Shehawi, A.M. and Harby, M.H. (2010) Genetic Diversity in Bread Wheat as Revealed by RAPD Markers and Its Relationship to Leaf Rust and Hybrid Performance. Alexandria Journal of Agricultural Research, 55, 31-38.
- Steel, R.G.D. and Torrie, J.H. (1984) Principles and Procedures of Statistics: A Biometrical Approach. 2nd McGraw Hill Book Co., Singapore.
- Griffing, B. (1956) Concept of General and Specific Combining Ability in Relation to Diallel Crossing Systems. Australian Journal of Biological Sciences, 9, 463-493.
- Fehr, W.R. (1987 and 1993) Principles of Cultivar Development. Vol. 1. Macmillan Publishing Company, New York. 536 p.
- Fonseca, S.N. (1965) Heterosis Heterobeltiosis Diallel Analysis and Gene Action in Crosses of Triticum aestivum L. Ph.D. Thesis, Purdue University, West Lafayette.
- Wynne, J.C., Emery, D.A. and Rice, P.W. (1970) Combining Ability Estimation in Arachis hypogaea L. II. Field Performance of F1 Hybrids. Crop Science, 10, 713-715. http://dx.doi.org/10.2135/cropsci1970.0011183X001000060036x
- Rui, W., Kun, N., Yi, W. and Lianmeng, D. (1996) The Combining Ability and Genetical Models of Spike Characteristics in Hexaploid Wheat. Acta Agriculturae Boreali-Occidentalis Sinica, 5, 1-5.
- Larik, A.S., Mahar, A.R., Kakar, A.A. and Shafkh, M.A. (1999) Heterosis, Inbreeding Depression and Combining Ability in Triticum Aestivum L. Journal of Plant Genetics, 25, 455-450.
- Joshi, S.K., Sharma, S.N., Singhania, D.L. and Sain, R.S. (2003) Hybrid Vigour over Environments in a Ten Parent Diallel Crosses in Common Wheat. SABRAO Journal of Breeding and Genetics, 35, 81-91.
- Joshi, S.K., Sharma, S.N., Singhania, D.L. and Sain, R.S. (2004) Combining Ability in the F1 and F2 Generations of Diallel Cross in Hexaploid Wheat (Triticum aestivum L. em. Thell). Hereditas, 141, 115-121. http://dx.doi.org/10.1111/j.1601-5223.2004.01730.x
- Singh, H., Sharma, S.N. and Sain, R.S. (2004) Heterosis Studies for Yield and Its Components in Bread Wheat over Environments. Hereditas, 141, 106-114. http://dx.doi.org/10.1111/j.1601-5223.2004.01728.x
- Singh, V., Krishna, R., Singh, S. and Vikram, P. (2008) Combining Ability and Heterosis Analysis for Yield Traits in Bread Wheat (Triticum aestivum). The Indian Journal of Agricultural Sciences, 82, 56-63.
- Singh, H., Sharma, S.N. and Sain, R.S. (2011) Combining Ability for Some Quantitative Characters in Hexaploid Wheat (Triticum aestivum L. em. Thell). Procedings of 4th International Crop Science Congress, Brisbane, 26 September-1 October 2004.
- Bao, Y.G., Sen, W., Wang, X., Wang, Y., Li, X., Wang, L. and Wang, H. (2010) Heterosis and Combining Ability for Major Yield Traits of a New Wheat Germplasm Shannong 0095 Derived from Thinopyrum intermedium. Agricultural Sciences in China, 8, 753-760. http://dx.doi.org/10.1016/S1671-2927(08)60275-8
- Gowda, M., Kling, C., Würschum, T., Liu, W., Maurer, H.P., Hahn, V. and Reif, J.C. (2010) Hybrid Breeding in Durum Wheat: Heterosis and Combining Ability. Crop Science, 50, 2224-2230. http://dx.doi.org/10.2135/cropsci2009.10.0637
- Anwar, J., Akbar, M., Hussain, M., Asghar, S., Ahmad, J. and Owais, M. (2011) Combining Ability Estimates for Grain Yield in Wheat. Journal of Agricultural Research, 49, 437-445.
- Barker, T.C. and Varughese, G. (2011) Combining Ability and Heterosis among Eight Complete Spring Hexaploid Triticate Lines. Crop Science, 32, 340-344. http://dx.doi.org/10.2135/cropsci1992.0011183X003200020012x
- Rizkalla, A.A., Hussien, B.A., Al-Ansary, A.M.F., Nasseef, J.E. and Hussein, M.H.A. (2012) Combining Ability and Heterosis Relative to RAPD Marker in Cultivated and Newly Hexaploid Wheat Varieties. Australian Journal of Basic and Applied Sciences, 6, 215-224.
- Beche, E., Silva, C.L., Pagliosa, E.S., Capelin, M.A., Franke, J., Matei, G. and Benin, G. (2013) Hybrid Performance and Heterosis in Early Segregant Populations of Brazilian Spring Wheat. Australian Journal of Crop Science, 7, 51-57.
- Zhongfu, N., Yao, Y., Peng, H., Hu, Z. and Qixin, S. (2014) Genomics and Heterosis in Hexaploid Wheat. Journal of Crop Science, 23, 215-220.
- Abd-Allah, M.H., Soheir, M.H. and Amin, I.A. (2013) Genotypic Differences for Heat Tolerance Traits in Bread. Wheat Using Five Parameters Genetic Model. Alex. J. Agric. Res., 58, 83-96.
- Eduardo, S.P., Silva, C.L. and Capelin, M.A. (2013) Hybrid Performance and Heterosis in Early Segregant Populations of Brazilian Spring Wheat. Journal of Crop Science, 45, 142-146.
- Al-Hamdany, Gh.A. (2010) Genetic Analysis of F2 Diallel Crosses in Durum Wheat. Mesopotamia J. of Agric, 38.
- Cifci1, E.A. and Yagdi, K. (2010) The Research of the Combining Ability of Agronomic Traits of Bread Wheat in F1 and F2 Generations. Journal of Agricultural Faculty of Uludag University, 24, 85-92.
- Awan, J.I., Malik, M.F.A. and Siddique, M. (2005) Combining Ability Analysis in Intervarietal Crosses for Component Traits in Hexaploid Wheat. Journal of Agriculture & Social Sciences, 1, 316-317.
- Coopera, J.K., Amir, M., Ibrahim, H., Rudda, J., Haysa, D., Mallaa, S. and Bakera, J. (2012) Increasing Hard Winter Wheat Yield Potential via Synthetic Hexaploid Wheat: II. Heritability and Combining Ability of Yield and Its Components. Crop Science, 53, 67-73. http://dx.doi.org/10.2135/cropsci2011.07.0383
- Hammad, S.M., El-Seidy, E.H. and Darwesh, M.A.H. (2012) Inheritances of Leaf Rust Resistance in Six Wheat Crosses. Journal of Agronomy, 9, 222-242.
- Koumber, R.M. and El-Gammaal, A.A. (2012) Inheritance and Gene Action for Yield and Its Attributes in Three Bread Wheat Crosses (Triticum aestivum L.). World Journal of Agricultural Sciences, 8, 156-162.
- Fellahi, Z.E.A., Hannachi, A., Bouzerzour, H. and Boutekrabt, A. (2013) Line × Tester Mating Design Analysis for Grain Yield and Yield Related Traits in Bread Wheat (Triticum aestivum L.). International Journal of Agronomy, 2013, Article ID: 201851. http://dx.doi.org/10.1155/2013/201851
- Morojele, E. and Labuschagne, M. (2013) Combining Ability of Quality Characteristics of Wheat Cultivars Grown in Lesotho. Journal of Crop Science, 21, 127-132.
- Zaazaa, E.I., Hager, M.A. and El-Hashash, E.F. (2012) Genetical Analysis of Some Quantitative Traits in Wheat Using Six Parameters Genetic Model. Journal of Agricultural Science, 12, 456-462.
NOTES

*Corresponding author.